

Abnormalities of Sphingolipids Metabolic Pathways in the Pathogenesis of Psoriasis

{kind=link}
{kind=link}
{kind=link}
Abstract
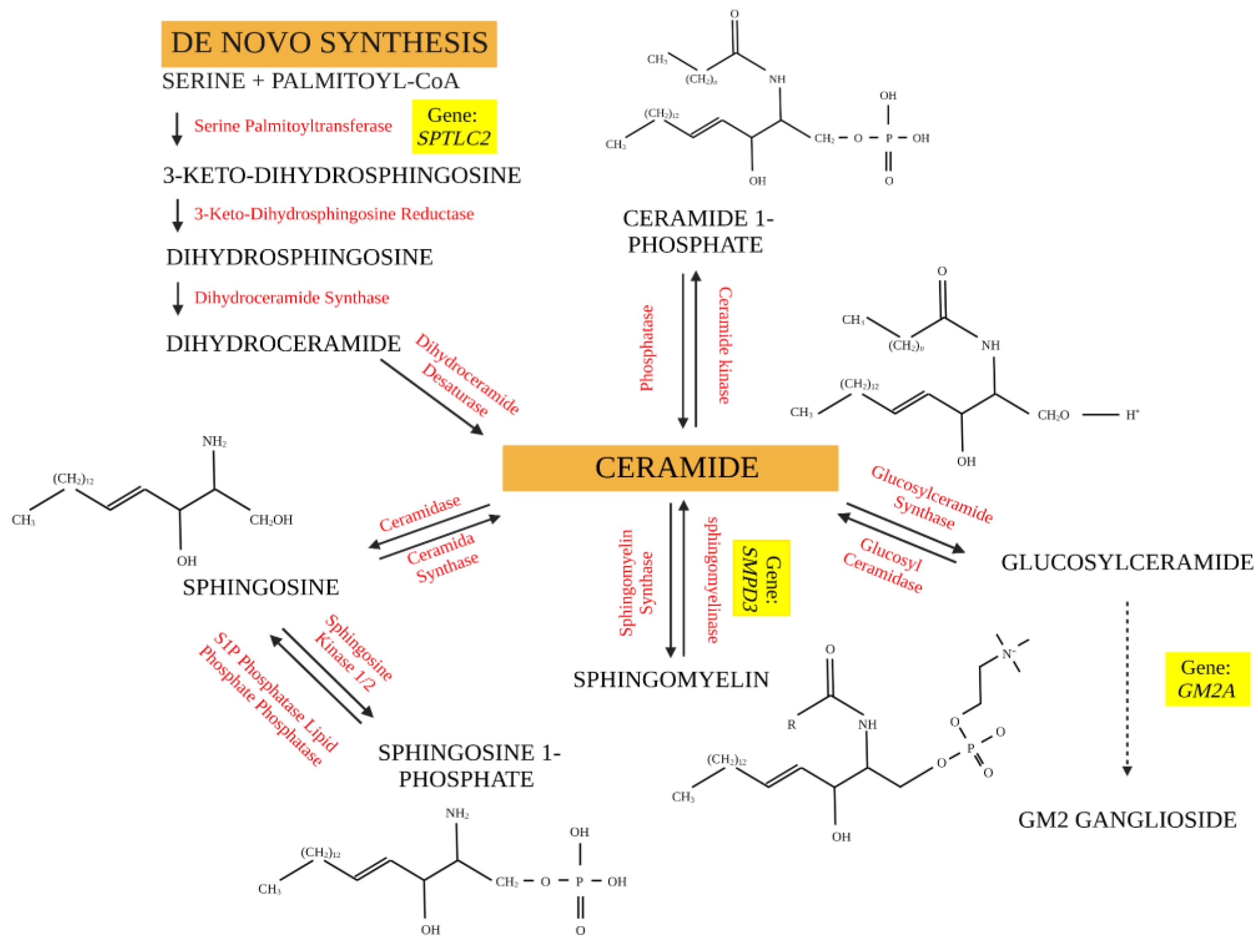
:1. Introduction
2. Methodology
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Meglio, P.D.; Perera, G.K.; Nestle, F.O. Review the Multitasking Organ: Recent Insights into Skin Immune Function. Immunity 2011, 35, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, R.; Chen, X.; Hu, J.; Fu, Y.; Peng, J.; Li, Y. Ghrelin Protects against Contact Dermatitis and Psoriasiform Skin Inflammation by Antagonizing TNF-α/NF-κ B Signaling Pathways. Sci. Rep. 2019, 9, 1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, S.; O’Boyle, N.M. Skin Lipids in Health and Disease: A Review. Chem. Phys. Lipids 2021, 236, 105055. [Google Scholar] [CrossRef] [PubMed]
- Koster, M.I.; Roop, D.R. Mechanisms Regulating Epithelial Stratification. Annu. Rev. Cell Dev. Biol. 2007, 23, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.; Horsley, V. Developing Stratified Epithelia: Lessons from the Epidermis and Thymus. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Luger, T.; Amagai, M.; Dreno, B.; Dagnelie, M.; Liao, W.; Kabashima, K.; Schikowski, T.; Proksch, E.; Elias, P.M.; Simon, M.; et al. Atopic Dermatitis: Role of the Skin Barrier, Environment, Microbiome, and Therapeutic Agents. J. Dermatol. Sci. 2021, 102, 142–157. [Google Scholar] [CrossRef]
- Pasquali, L.; Srivastava, A.; Meisgen, F.; Mahapatra, K.D.; Xia, P.; Landén, N.X.; Pivarcsi, A.; Sonkoly, E. The Keratinocyte Transcriptome in Psoriasis: Pathways Related to Immune Responses, Cell Cycle and Keratinization. Acta Derm. Venereol. 2019, 99, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Matejuk, A. Skin Immunity. Arch. Immunol. Ther. Exp. 2018, 66, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, A.W.; Read, C. Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. JAMA-J. Am. Med. Assoc. 2020, 323, 1945–1960. [Google Scholar] [CrossRef]
- Mysliwiec, H.; Baran, A.; Harasim-Symbor, E.; Mysliwiec, P.; Milewska, A.; Chabowski, A.; Flisiak, I. Serum Fatty Acid Profile in Psoriasis and Its Comorbidity. Arch Dermatol. Res. 2017, 309, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, Regional, and Worldwide Epidemiology of Psoriasis: Systematic Analysis and Modelling Study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Capon, F. The Genetic Basis of Psoriasis. Int. J. Mol. Sci. 2017, 18, 2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iskandar, I.Y.K.D.; Parisi, R.D.; Griffiths, C.E.M.; Ashcroft, D.M. Systematic Review Examining Changes over Time and Variation in the Incidence and Prevalence of Psoriasis by Age and Gender. Br. J. Dermatol. 2021, 184, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Pezzolo, E.; Naldi, L. Epidemiology of Major Chronic Inflammatory Immune-Related Skin Diseases in 2019. Expert Rev. Clin. Immunol. 2020, 16, 155–166. [Google Scholar] [CrossRef]
- Kamiya, K.; Kishimoto, M.; Sugai, J.; Komine, M.; Ohtsuki, M. Risk Factors for the Development of Psoriasis. Int. J. Mol. Sci. 2019, 20, 4347. [Google Scholar] [CrossRef] [Green Version]
- Pietrzak, A.; Michalak-stoma, A.; Szepietowski, J.C. Lipid Disturbances in Psoriasis: An Update. Mediat. Inflammatio 2010, 2010, 535612. [Google Scholar] [CrossRef] [Green Version]
- Nowowiejska, J.; Baran, A.; Flisiak, I. Aberrations in Lipid Expression and Metabolism in Psoriasis. Int. J. Mol. Sci. 2021, 22, 6561. [Google Scholar] [CrossRef]
- Faber, E.; Nail, M.; Watison, W. Natural History of Psoriasis in 61 Twin Pairs. Arch Dermatol. 1974, 109, 207–211. [Google Scholar] [CrossRef]
- Duffy, D.; Spelman, L.; Martin, N. Psoriasis in Australian Twins. J. Am. Acad Dermatol. 1993, 29, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandrup, F.; Hauge, M.; Henningsen, K.; Eriksen, B. Psoriasis in an Unselected Series of Twins. Arch. Dermatol. 1978, 114, 874–878. [Google Scholar] [CrossRef]
- Dand, N.; Duckworth, M.; Baudry, D.; Russell, A.; Curtis, C.J.; Lee, S.H.; Evans, I.; Mason, K.J.; Alsharqi, A.; Becher, G.; et al. HLA-C*06:02 Genotype Is a Predictive Biomarker of Biologic Treatment Response in Psoriasis. J. Allergy Clin. Immunol. 2019, 143, 2120–2130. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.P.; Stuart, P.E.; Nistor, I.; Hiremagalore, R.; Chia, N.V.C.; Jenisch, S.; Weichenthal, M.; Abecasis, G.R.; Lim, H.W.; Christophers, E.; et al. Sequence and Haplotype Analysis Supports HLA-C as the Psoriasis Susceptibility 1 Gene. Am. J. Hum. Genet. 2006, 78, 827–851. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.P.; Duffin, K.C.; Helms, C.; Ding, J.; Stuart, P.E.; Goldgar, D.; Gudjonsson, J.E.; Li, Y.; Tejasvi, T.; Feng, B.J.; et al. Genome-Wide Scan Reveals Association of Psoriasis with IL-23 and NF-ΚB Pathways. Nat. Genet. 2009, 41, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strange, A.; Capon, F.; Spencer, C.C.; Knight, J.; Weale, M.E.; Allen, M.H.; Barton, A.; Band, G.; Bellenguez, C.; Bergboer, J.G.; et al. Genome-Wide Association Study Identifies New Psoriasis Susceptibility Loci and an Interaction between HLA-C and ERAP1. Nat Genet. 2010, 42, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Cao, L.; Roberson, E.D.O.; Duan, S.; Helms, C.A.; Nair, R.P.; Duffin, K.C.; Stuart, P.E.; Goldgar, D.; Hayashi, G.; et al. Rare and Common Variants in CARD14, Encoding an Epidermal Regulator of NF-KappaB, in Psoriasis. Am. J. Hum. Genet. 2012, 90, 796–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berki, D.M.; Liu, L.; Choon, S.E.; Burden, A.D.; Griffiths, C.E.M.; Navarini, A.A.; Tan, E.S.; Irvine, A.D.; Ranki, A.; Ogo, T.; et al. Activating CARD14 Mutations Are Associated with Generalized Pustular Psoriasis but Rarely Account for Familial Recurrence in Psoriasis Vulgaris. J. Investig. Dermatol. 2015, 135, 2964–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, O.; Nestle, M.D.; Daniel, H.; Kaplan, M.D.; Jonathan Barker, M. Mechanisms of Disease Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef]
- Albanesi, C.; Madonna, S.; Gisondi, P.; Girolomoni, G. The Interplay between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front. Immunol. 2018, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Zelga, P.; Kutwin, M.; Migdalska-s, M.; Brzezia, E. An Analysis of IL-10, IL-17A, IL-17RA, IL-23A and IL-23R Expression and Their Correlation with Clinical Course in Patients with Psoriasis. J. Clin. Med. 2021, 10, 5834. [Google Scholar] [CrossRef]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The Immunological Anatomy of the Skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef]
- Abdallah, F.; Mijouin, L.; Pichon, C. Skin Immune Landscape: Inside and Outside the Organism. Mediat. Inflamm. 2017, 2017, 5095293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Chang, C.; Lu, Q. The Inflammatory Response in Psoriasis: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 50, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chen, Y.; Cui, L.; Shi, Y.; Guo, C. Advances in the Pathogenesis of Psoriasis: From Keratinocyte Perspective. Cell Death Dis. 2022, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Wang, G. Keratin 17: A Critical Player in the Pathogenesis of Psoriasis. Med. Res. Rev. 2014, 34, 438–454. [Google Scholar] [CrossRef]
- Shi, X.; Jin, L.; Dang, E.; Chang, T.; Feng, Z.; Liu, Y.; Wang, G. IL-17A Upregulates Keratin 17 Expression in Keratinocytes through STAT1-and STAT3-Dependent Mechanisms. J. Investig. Dermatol. 2011, 131, 2401–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwicky, P.; Ingelfinger, F.; de Melo, B.M.S.; Ruchti, F.; Schärli, S.; Puertas, N.; Lutz, M.; Phan, T.S.; Kündig, T.M.; Levesque, M.P.; et al. IL-12 Regulates Type 3 Immunity through Interfollicular Keratinocytes in Psoriasiform Inflammation. Sci. Immunol. 2021, 6, eabg9012. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicic, M.; Kaštelan, M.; Brajac, I.; Sotošek, V.; Massari, L.P. Current Concepts of Psoriasis Immunopathogenesis. Int. J. Mol. Sci. 2021, 22, 11574. [Google Scholar] [CrossRef]
- Rudan, M.V.; Watt, F.M. Mammalian Epidermis: A Compendium of Lipid Functionality. Front. Physiol. 2022, 12, 804824. [Google Scholar] [CrossRef]
- Fekry, B.; Esmaeilniakooshkghazi, A.; Krupenko, S.A.; Krupenko, N.I. Ceramide Synthase 6 Is a Novel Target of Methotrexate Mediating Its Antiproliferative Effect in a P53-Dependent Manner. PLoS ONE 2016, 19, e0146618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmond, V.; Dufour, F.; Poiroux, G.; Shoji, K.; Malleter, M.; Fouqué, A.; Tauzin, S.; Rimokh, R.; Sergent, O.; Penna, A.; et al. Downregulation of Ceramide Synthase-6 during Epithelial-to-Mesenchymal Transition Reduces Plasma Membrane Fluidity and Cancer Cell Motility. Oncogene 2015, 24, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Milstien, S. The Outs and the Ins of Sphingosine-1-Phosphate in Immunity. Nat. Rev. Immunol. 2011, 11, 403–415. [Google Scholar] [CrossRef]
- Aida, J.; Higuchi, S.; Hasegawa, Y.; Nagano-ito, M.; Hirabayashi, Y.; Banba, A.; Shimizu, T.; Kikuchi, A.; Saga, M.; Ichikawa, S. Up-Regulation of Ceramide Glucosyltransferase during the Differentiation of U937 Cells. J. Biochem. 2011, 150, 303–310. [Google Scholar] [CrossRef]
- Borodzicz, S.; Rudnicka, L.; Mirowska-guzel, D.; Cudnoch-jedrzejewska, A. The Role of Epidermal Sphingolipids in Dermatologic Diseases. Lipids Health Dis. 2016, 15, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, T. Sphingolipids as Functional Food Components: Benefits in Skin Improvement and Disease Prevention. J. Agric. Food Chem. 2022, 70, 9597–9609. [Google Scholar] [CrossRef]
- Kindt, R.; Jorge, L.; Dumont, E.; Couturon, P.; David, F.; Sandra, P.; Sandra, K. Profiling and Characterizing Skin Ceramides Using Reversed-Phase Liquid Chromatography—Quadrupole Time-of-Flight Mass Spectrometry. Anal. Chem. 2012, 84, 403–411. [Google Scholar] [CrossRef]
- Meckfessel, M.H.; Brandt, S. The Structure, Function, and Importance of Ceramides in Skin and Their Use as Therapeutic Agents in Skin-Care Products. J. Am. Dermatol. 2014, 71, 177–184. [Google Scholar] [CrossRef]
- Ishikawa, J. Changes in the Ceramide Profile of Atopic Dermatitis Patients. J. Investig. Dermatol. 2010, 108, 2511–2514. [Google Scholar] [CrossRef] [Green Version]
- Joo, K.-M.; Nam, G.-W.; Park, S.Y.; Han, J.Y.; Jeong, H.-J.; Lee, S.-Y.; Kim, H.K.; Lim, K.-M. Relationship between Cutaneous Barrier Function and Ceramide Species in Human Stratum Corneum. J. Dermatol. Sci. 2010, 60, 47–50. [Google Scholar] [CrossRef]
- Janůšová, B.; Zbytovská, J.; Lorenc, P.; Vavrysová, H.; Palát, K.; Hrabálek, A.; Vávrová, K. Effect of Ceramide Acyl Chain Length on Skin Permeability and Thermotropic Phase Behavior of Model Stratum Corneum Lipid Membranes. Biochim. Biophys. 2011, 1811, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Park, K. Ceramides in Skin Health and Disease: An Update. Am. J. Clin. Dermatol. 2021, 22, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Uche, L.E.; Gooris, G.S.; Bouwstra, J.A.; Beddoes, C.M. Increased Levels of Short-Chain Ceramides Modify the Lipid Organization and Reduce the Lipid Barrier of Skin Model Membranes. Langmuir 2021, 37, 9478–9489. [Google Scholar] [CrossRef] [PubMed]
- Tawada, C.; Kanoh, H.; Nakamura, M.; Mizutani, Y.; Fujisawa, T.; Banno, Y.; Seishima, M. Interferon-c Decreases Ceramides with Long-Chain Fatty Acids: Possible Involvement in Atopic Dermatitis and Psoriasis. J. Investig. Dermatol. 2014, 134, 712–718. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sandhoff, R.; Kono, M.; Zerfas, P.; Hoffmann, V.; Ding, B.C.-H.; Deng, R.L.P. Chu-Xia Depletion of Ceramides with Very Long Chain Fatty Acids Causes Defective Skin Permeability Barrier Function, and Neonatal Lethality in ELOVL4 Deficient Mice. Int. J. Biol. Sci. 2007, 3, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.P.; Vaidyanathan, N.; Rengasamy, M.; Oommen, A.M.; Somaiya, N.; Jagannath, M.R. Sphingolipid Metabolic Pathway: An Overview of Major Roles Played in Human Diseases. J. Lipids 2013, 2013, 178910. [Google Scholar] [CrossRef] [Green Version]
- Hartel, J.C.; Merz, N.; Grösch, S. How Sphingolipids Affect T Cells in the Resolution of Inflammation. Front. Pharmacol. 2022, 13, 3784. [Google Scholar] [CrossRef]
- Obinata, H.; Hla, T. Sphingosine 1-Phosphate and Inflammation. Int. Immunol. 2019, 31, 617–625. [Google Scholar] [CrossRef]
- Nomoto, K.; Itaya, Y.; Watanabe, K.; Yamashita, T.; Okazaki, T.; Tokudome, Y. Epidermal Permeability Barrier Function and Sphingolipid Content in the Skin of Sphingomyelin Synthase 2 Deficient Mice. Exp. Dermatol. 2018, 27, 827–832. [Google Scholar] [CrossRef]
- Hollmann, C.; Werner, S.; Avota, E.; Reuter, D.; Japtok, L.; Kleuser, B.; Gulbins, E.; Becker, K.A.; Schneider-Schaulies, J.; Beyersdorf, N. Inhibition of Acid Sphingomyelinase Allows for Selective Targeting of CD4 + Conventional versus Foxp3 + Regulatory T Cells. J. Immunol. 2016, 197, 3130–3141. [Google Scholar] [CrossRef] [Green Version]
- Kanda, N.; Hoashi, T.; Saeki, H. The Defect in Regulatory T Cells in Psoriasis and Therapeutic Approaches. J. Clin. Med. 2021, 10, 3880. [Google Scholar] [CrossRef]
- Bocheńska, K.; Gabig-Cimińska, M. Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis. Molecules 2020, 25, 1130. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Parashuraman, S.; Angelo, G.D. Glycosphingolipid—Protein Interaction in Signal Transduction. Int. J. Mol. Sci. 2016, 17, 1732. [Google Scholar] [CrossRef] [Green Version]
- Paller, A.S.; Arnsmeier, S.L.; Alvarez-franco, M.; Bremer, E.G. Ganglioside GM3 Inhibits the Proliferation of Cultured Keratinocytes. J. Investig. Dermatol. 1993, 100, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.B.; Bonish, B.K.; Chaturvedi, V.; Qin, J.; Nickoloff, B.J. Keratinocyte CDw60 Expression Is Modulated by Both a Th-1 Type Cytokine IFN-γ and Th-2 Cytokines IL-4 and IL-13: Relevance to Psoriasis. J. Investig. Dermatol. 2001, 116, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Gong, Y.; Cui, L.; Hu, Y.; Zhou, Q.; Chen, Z. High-Throughput Transcriptome and Pathogenesis Analysis of Clinical Psoriasis. J. Dermatol. Sci. 2020, 98, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Rønholt, K.; Nielsen, A.L.; Johansen, C.; Vestergaard, C.; Fauerbye, A.; López-vales, R.; Dinarello, C.A.; Iversen, L. IL-37 Expression Is Downregulated in Lesional Psoriasis Skin IL-37 Expression Is Downregulated in Lesional Psoriasis Skin. Immunohorizons 2020, 4, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Page, K.M.; Suarez-farinas, M.; Suprun, M.; Zhang, W.; Garcet, S.; Fuentes-duculan, J.; Li, X.; Scaramozza, M.; Kieras, E.; Banfield, C.; et al. Molecular and Cellular Responses to the TYK2/JAK1 Inhibitor PF-06700841 Reveal Reduction of Skin Inflammation in Plaque Psoriasis. J. Investig. Dermatol. 2020, 140, 1546–1555.e4. [Google Scholar] [CrossRef]
- Matwiejuk, M.; Mysliwiec, H.; Chabowski, A.; Flisiak, I. The Role of Sphingolipids in the Pathogenesis of Psoriasis. Metabolities 2022, 12, 1171. [Google Scholar] [CrossRef]
- Li, Q.; Fang, H.; Dang, E.; Wang, G. The Role of Ceramides in Skin Homeostasis and in Fl Ammatory Skin Diseases. J. Dermatol. Sci. 2020, 97, 2–8. [Google Scholar] [CrossRef]
- Moore, D.J.; Rawlings, A.V. The Chemistry, Function and (Patho) Physiology of Stratum Corneum Barrier Ceramides. Int. J. Cosmet. Sci. 2017, 39, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Kim, S.K.; Park, C.S. Novel Phytoceramides Containing Fatty Acids of Diverse Chain Lengths Are Better than a Single C18-Ceramide N-Stearoyl Phytosphingosine to Improve the Physiological Properties of Human Stratum Corneum. Clin. Cosmet. Investig. Dermatol. 2017, 10, 363–371. [Google Scholar]
- Qiao, P.; Guo, W.; Ke, Y.; Fang, H.; Zhuang, Y.; Jiang, M.; Zhang, J.; Shen, S.; Qiao, H.; Dang, E.; et al. Mechanical Stretch Exacerbates Psoriasis by Stimulating Keratinocyte Proliferation and Cytokine Production. J. Investig. Dermatol. 2019, 139, 1470–1479. [Google Scholar] [CrossRef] [Green Version]
- Motta, S.; Monti, M.; Sesana, S.; Mellesi, L.; Ghidoni, R.; Caputo, R. Abnormality of Water Barrier Function in Psoriasis: Role of Ceramides Fractions. Arch. Dermatol. 1994, 130, 452–456. [Google Scholar] [CrossRef]
- Motta, S.; Monti, M.; Sesana, S.; Caputo, R.; Carelli, S.; Ghidoni, R. Ceramide Composition of the Psoriatic Scale. Biochim. Biophys. Acta 1993, 1182, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Checa, A.; Xu, N.; Sar, D.G.; Haeggström, J.Z.; Ståhle, M.; Wheelock, C.E. Circulating Levels of Sphingosine-1-Phosphate Are Elevated in Severe, but Not Mild Psoriasis and Are Unresponsive to Anti-TNF-α Treatment. Sci. Rep. 2015, 15, 12017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Lew, B.; Seong, K.; Kim, N. An Inverse Relationship between Ceramide Synthesis and Clinical Severity in Patients with Psoriasis. J. Korean Med. Sci. 2004, 19, 859–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokose, U.; Ishikawa, J.; Morokuma, Y.; Naoe, A.; Inoue, Y.; Yasuda, Y.; Tsujimura, H.; Fujimura, T.; Murase, T.; Hatamochi, A. The Ceramide [NP]/[NS] Ratio in the Stratum Corneum Is a Potential Marker for Skin Properties and Epidermal Differentiation. BMC Dermatol. 2020, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.V.; Domenichiello, A.F.; Dey, A.K.; Yuan, Z.; Rose, S.M.; Playford, M.P.; Ramsden, C.E.; Nehal, N.; Diseases, C.; Branch, C.; et al. Bioactive Lipid Mediator Profiles in Human Psoriasis Skin and Blood. J. Investig. Dermatol. 2018, 138, 1518–1528. [Google Scholar] [CrossRef] [Green Version]
- Rioux, G.; Ridha, Z.; Simard, M.; Turgeon, F.; Guérin, S.L.; Pouliot, R. Transcriptome Profiling Analyses in Psoriasis: A Dynamic Contribution of Keratinocytes to the Pathogenesis. Genes 2020, 11, 1155. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Morin, S.; Ridha, Z.; Pouliot, R. Current Knowledge of the Implication of Lipid Mediators in Psoriasis. Front. Immunol. 2022, 13, 961107. [Google Scholar] [CrossRef]
- Setkowicz, M.; Mastalerz, L.; Gielicz, A.; Wojas-Pelc, A.; Sanak, M. Lack of Association of ALOX12 and ALOX15B Polymorphisms with Psoriasis despite Altered Urinary Excretion of 12(S)-Hydroxyeicosatetraenoic Acid. Br. J. Dermatol. 2015, 172, 337–344. [Google Scholar] [CrossRef]
- Doolen, S.; Keyes, G.S.; Ramsden, C.E. Hydroxy-Epoxide and Keto-Epoxide Derivatives of Linoleic Acid Activate Trigeminal Neurons. Neurobiol. Pain 2020, 7, 100046. [Google Scholar] [CrossRef]
- Zimmer, B.; Angioni, C.; Osthues, T.; Toewe, A.; Thomas, D.; Pierre, S.C.; Geisslinger, G.; Scholich, K.; Sisignano, M. The Oxidized Linoleic Acid Metabolite 12,13-DiHOME Mediates Thermal Hyperalgesia during Inflammatory Pain. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2018, 1863, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Garcia, A.; Thomas, C.P.; Keeney, D.S.; Zheng, Y.; Brash, A.R. The Importance of the Lipoxygenase-Hepoxilin Pathway in the Mammalian Epidermal Barrier. Biochim. Biophys Acta 2014, 1841, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Krieg, P.; Fürstenberger, G. The Role of Lipoxygenases in Epidermis. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2014, 1841, 390–400. [Google Scholar] [CrossRef]
- Shao, S.; Harms, P.W.; Gudjonsson, J.E.; Shao, S.; Chen, J.; Swindell, W.R.; Tsoi, L.C.; Xing, X.; Ma, F.; Uppala, R.; et al. Phospholipase A2 Enzymes Represent a Shared Pathogenic Pathway in Psoriasis and Pityriasis Rubra Pilaris. JCI Insight 2021, 6, e151911. [Google Scholar] [CrossRef] [PubMed]
- Chiba, H.; Michibata, H.; Wakimoto, K.; Seishima, M.; Kawasaki, S.; Okubo, K.; Mitsui, H.; Torii, H.; Imai, Y. Cloning of a Gene for a Novel Epithelium-Specific Cytosolic Phospholipase A 2, CPLA 2g, Induced in Psoriatic Skin. J. Biol. Chem. 2004, 279, 12890–12897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaranta, M.; Knapp, B.; Garzorz, N.; Mattii, M.; Pullabhatla, V.; Pennino, D.; Andres, C.; Traidl-hoffmann, C.; Cavani, A.; Theis, F.J.; et al. Intraindividual Genome Expression Analysis Reveals a Specific Molecular Signature of Psoriasis and Eczema. Sci. Transl. Med. 2014, 6, 244ra90. [Google Scholar] [CrossRef]
- Cheung, K.L.; Jarrett, R.; Subramaniam, S.; Salimi, M.; Owsiak, D.G.; Chen, Y.L.; Hardman, C.; Xue, L.; Cerundolo, V.; Ogg, G. Psoriatic T Cells Recognize Neolipid Antigens Generated by Mast Cell Phospholipase Delivered by Exosomes and Presented by CD1a. J. Exp. Med. 2016, 213, 2399–2412. [Google Scholar] [CrossRef] [Green Version]
- Chow, M.; Lai, K.; Francisco, S.; Ahn, R.; Francisco, S.; Gupta, R.; Francisco, S.; Arron, S.; Francisco, S. Effect of Adalimumab on Gene Expression Profiles of Psoriatic Skin and Blood. J. Drugs Dermatol. 2016, 15, 988–994. [Google Scholar]
- Snezhkina, A.V.; Lukyanova, E.N.; Fedorova, M.S.; Kalinin, D.V.; Melnikova, N.V. Novel Genes Associated with the Development of Carotid Paragangliomas. Mol. Biol. 2019, 53, 547–559. [Google Scholar] [CrossRef]
- Ye, J.; Liu, P.; Li, R.; Liu, H.; Pei, W.; Ma, C.; Shen, B.; Zhao, D.; Chen, X. Biomarkers of Connective Tissue Disease-Associated Interstitial Lung Disease in Bronchoalveolar Lavage Fluid: A Label-Free Mass Based Relative Quantification Study. J. Clin. Lab. Anal. 2022, 36, e24367. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Zhang, C.; Zhang, C.; Guan, X.; Jia, W. Differences of Macrophages in the Tumor Microenvironment as an Underlying Key Factor in Glioma Patients. Front. Immunol. 2022, 13, 6549. [Google Scholar] [CrossRef]
- Khatri, I.; Bhasin, M.K. A Transcriptomics-Based Meta-Analysis Combined with Machine Learning Identifies a Secretory Biomarker Panel for Diagnosis of Pancreatic Adenocarcinoma. Front. Genet. 2020, 11, 572284. [Google Scholar] [CrossRef] [PubMed]
- Corda, D.; Mosca, M.G.; Ohshima, N.; Grauso, L.; Yanaka, N. The Emerging Physiological Roles of the Glycerophosphodiesterase Family. FEBS J. 2014, 281, 998–1016. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.M.C. Liver Glycerylphosphorylcholine Diesterase. Biochem. J. 1956, 62, 689–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogusa, Y.; Fujioka, Y.; Komatsu, R.; Kato, N.; Yanaka, N. Isolation and Characterization of Two Serpentine Membrane Proteins Containing Glycerophosphodiester Phosphodiesterase, GDE2 and GDE6. Gene 2004, 337, 173–179. [Google Scholar] [CrossRef]
- Chang, P.A.; Shao, H.B.; Long, D.X.; Sun, Q.; Wu, Y.J. Isolation, Characterization and Molecular 3D Model of Human GDE4, a Novel Membrane Protein Containing Glycerophosphodiester Phosphodiesterase Domain. Mol. Membr. Biol. 2008, 25, 557–566. [Google Scholar] [CrossRef]
- Tserendavga, B.; Ohshima, N.; Fujita, C.; Yuzawa, K.; Ohshima, M.; Yanaka, N.; Minamishima, Y.A.; Izumi, T. Characterization of Recombinant Murine GDE4 and GDE7, Enzymes Producing Lysophosphatidic Acid and/or Cyclic Phosphatidic Acid. J. Biochem. 2021, 170, 713–727. [Google Scholar] [CrossRef]
- Lei, L.; Su, J.; Chen, J.; Chen, W.; Chen, X.; Peng, C. The Role of Lysophosphatidic Acid in the Physiology and Pathology of the Skin. Life Sci. 2019, 220, 194–200. [Google Scholar] [CrossRef]
- Gaire, B.P.; Lee, C.-H.; Kim, W.; Sapkota, A.; Lee, D.Y.; Choi, J.W. Lysophosphatidic Acid Receptor 5 Contributes to Imiquimod-Induced Psoriasis-Like Lesions through NLRP3 Inflammasome Activation in Macrophages. Cells 2020, 9, 1753. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, H.; Baek, J.; Roh, J. Lysophosphatidic Acid Mediates Imiquimod-Induced Psoriasis-like Symptoms by Promoting Keratinocyte Proliferation through LPAR1/ROCK2/PI3K/AKT Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 10777. [Google Scholar] [CrossRef] [PubMed]
- Key, C.C.; Bishop, A.C.; Wang, X.; Zhao, Q.; Chen, G.; Quinn, M.A.; Zhu, X.; Zhang, Q.; Parks, J.S.; Zhang, Q.; et al. Human GDPD3 Overexpression Promotes Liver Steatosis by Increasing Lysophosphatidic Acid Production and Fatty Acid Uptake. J. Lipid Res. Vol. 2020, 61, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Naka, K.; Ochiai, R.; Hoshii, T.; Araki, M.; Mitani, K.; Kim, D.; Matsubara, E.; Kondo, C.; Yang, K.; Araki, K.; et al. The Lysophospholipase D Enzyme Gdpd3 Is Required to Maintain Chronic Myelogenous Leukaemia Stem Cells. Nat. Commun. 2020, 11, 4681. [Google Scholar] [CrossRef]
- Conzelmann, E.; Sandhoff, K. AB Variant of Infantile GM2 Gangliosidosis: Deficiency of a Factor Necessary for Stimulation of Hexosaminidase A-Catalyzed Degradation of Ganglioside GM2 and Glycolipid GA2. Proc. Natl. Acad. Sci. USA 1978, 75, 3979–3983. [Google Scholar] [CrossRef] [Green Version]
- Lew, B.-L.; Cho, Y.; Kim, J.; Sim, W.-Y.; Kim, N.-I. Ceramides and Cell Signaling Molecules in Psoriatic Epidermis: Reduced Levels of Ceramides, PKC-, and JNK. J. Korean Med. Sci. 2006, 21, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Bielawska, A.; Linardic, C.M.; Hannun, Y.A. Ceramide-Mediated Biology. Determination of Structural and Stereospecific Requirements through the Use of N-Acyl-Phenylaminoalcohol Analogs. J. Biol. Chem. 1992, 267, 18493–18497. [Google Scholar] [CrossRef]
- Clarke, C.J.; Hannun, Y.A. Neutral Sphingomyelinases and NSMase2: Bridging the Gaps. Biochim. Biophys. Acta 2006, 1758, 1893–1901. [Google Scholar] [CrossRef] [Green Version]
- Sindhu, S.; Hei, Y.; Arefanian, H.; Ahmad, F.A.R.; Madiraju, S.R.M.; Prentki, M. Neutral Sphingomyelinase-2 and Cardiometabolic Diseases. Obes. Rev. 2021, 22, e13248. [Google Scholar] [CrossRef]
- Airola, M.V.; Shanbhogue, P.; Shamseddine, A.A.; Guja, K.E.; Senkal, C.E.; Maini, R. Structure of Human NSMase2 Reveals an Interdomain Allosteric Activation Mechanism for Ceramide Generation. Proc. Natl. Acad. Sci. USA 2017, 114, E5549–E5558. [Google Scholar] [CrossRef] [Green Version]
- Clarke, C.J.; Cloessner, E.A.; Roddy, P.L.; Hannun, Y.A. Neutral Sphingomyelinase 2 (NSMase2) Is the Primary Neutral Sphingomyelinase Isoform Activated by Tumour Necrosis Factor-α. Biochem. J. 2011, 435, 381–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashed, F.A.; Ahmad, Z.; Thomas, R.; Melhem, M.; Snider, A.J.; Obeid, L.M.; Mulla, F.A.; Hannun, Y.A.; Ahmad, R. Neutral Sphingomyelinase 2 Regulates Inflammatory Responses in Monocytes/Macrophages Induced by TNF-α. Sci. Rep. 2020, 10, 16802. [Google Scholar] [CrossRef] [PubMed]
- Buisson-Legendre, N.; Emonard, H.; Bernard, P.; Hornebeck, W. Relationship between Cell-Associated Matrix Metalloproteinase 9 and Psoriatic Keratinocyte Growth. J. Investig. Dermatol. 2000, 115, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhu, Z.; Li, Q.; Lin, Y.; Dang, E.; Meng, H.; Sha, N. Neutrophils Enhance Cutaneous Vascular Dilation and Permeability to Aggravate Psoriasis by Releasing Matrix Metallopeptidase 9. J. Investig. Dermatol. 2021, 141, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Alessandrini, F.; Stachowitz, S.; Ring, J.; Behrendt, H. The Level of Prosaposin Is Decreased in the Skin of Patients with Psoriasis Vulgaris. J. Investig. Dermatol. 2001, 116, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Merleev, A.A.; Le, S.T.; Alexanian, C.; Toussi, A.; Xie, Y.; Marusina, A.I.; Watkins, S.M.; Patel, F.; Billi, A.C.; Wiedemann, J.; et al. Biogeographic and Disease-Specific Alterations in Epidermal Lipid Composition and Single-Cell Analysis of Acral Keratinocytes. JCI Insight 2022, 7, e159762. [Google Scholar] [CrossRef]
- Bowser, P.A.; Gray, G.M. Sphingomyelinase in Pig and Human Epidermis. J. Investig. Dermatol. 1978, 70, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Miyai, M.; Hamada, M.; Moriguchi, T.; Hiruma, J.; Kamitani-kawamoto, A.; Watanabe, H.; Hara-chikuma, M.; Takahashi, K.; Takahashi, S.; Kataoka, K. Transcription Factor MafB Coordinates Epidermal Keratinocyte Differentiation. J. Investig. Dermatol. 2016, 136, 1848–1857. [Google Scholar] [CrossRef] [Green Version]
- Shima, K.; Inoue, T.; Uehara, Y.; Iwamura, M.; Fukagawa, S. Non-Invasive Transcriptomic Analysis Using MRNAs in Skin Surface Lipids Obtained from Children with Mild-to-Moderate Atopic Dermatitis. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1477–1485. [Google Scholar] [CrossRef]
- Brunner, P.M.; Guttman-yassky, E.; Leung, D.Y.M. The Immunology of Atopic Dermatitis and Its Reversibility with Broad-Spectrum and Targeted Therapies. J. Allergy Clin. Immunol. 2018, 139, S65–S76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boer, D.E.C.; Smeden, J.V.; Al-khakany, H.; Melnik, E.; Dijk, R.V.; Absalah, S.; Vreeken, R.J.; Haenen, C.C.P.; Lavrijsen, A.P.M.; Overkleeft, H.S.; et al. Skin of Atopic Dermatitis Patients Shows Disturbed β-Glucocerebrosidase and Acid Sphingomyelinase Activity that Relates to Changes in Stratum Corneum Lipid Composition. BBA-Mol. Cell Biol. Lipids 2020, 1865, 158673. [Google Scholar] [CrossRef] [PubMed]
- Alessandrini, F.; Behrendt, H.; Abeck, D.; Ring, J.; Stachowitz, S. Permeability Barrier Disruption Increases the Level of Serine Palmitoyltransferase in Human Epidermis. J. Investig. Dermatol. 2002, 119, 1048–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burger, B.; Sagiorato, R.N.; Cavenaghi, I.; Rodrigues, H.G. Abnormalities of Sphingolipids Metabolic Pathways in the Pathogenesis of Psoriasis. Metabolites 2023, 13, 291. https://doi.org/10.3390/metabo13020291
Burger B, Sagiorato RN, Cavenaghi I, Rodrigues HG. Abnormalities of Sphingolipids Metabolic Pathways in the Pathogenesis of Psoriasis. Metabolites. 2023; 13(2):291. https://doi.org/10.3390/metabo13020291
Chicago/Turabian StyleBurger, Beatriz, Roberta Nicolli Sagiorato, Isabella Cavenaghi, and Hosana Gomes Rodrigues. 2023. "Abnormalities of Sphingolipids Metabolic Pathways in the Pathogenesis of Psoriasis" Metabolites 13, no. 2: 291. https://doi.org/10.3390/metabo13020291
APA StyleBurger, B., Sagiorato, R. N., Cavenaghi, I., & Rodrigues, H. G. (2023). Abnormalities of Sphingolipids Metabolic Pathways in the Pathogenesis of Psoriasis. Metabolites, 13(2), 291. https://doi.org/10.3390/metabo13020291