

Co-Localization of Resistance and Metabolic Quantitative Trait Loci on Carrot Genome Reveals Fungitoxic Terpenes and Related Candidate Genes Associated with the Resistance to Alternaria dauci

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design and Crop Management
2.3. Sampling Design
2.4. Headspace Solid-Phase Microextraction Followed by Gas Chromatography–Mass Spectrometry (HS-SPME-GC-MS) for Terpene Analyses
2.5. Correlation between Metabolite Accumulation and Disease Score
2.6. Resistance and Metabolite-QTL Detection
2.7. Transcriptomic Analysis
2.8. Statistical Analysis and Highlighting of Differentially Expressed Genes within the Co-Localization Area
2.9. Fungal Growth Inhibition Assays
3. Results
3.1. Consistency of rQTL among Years
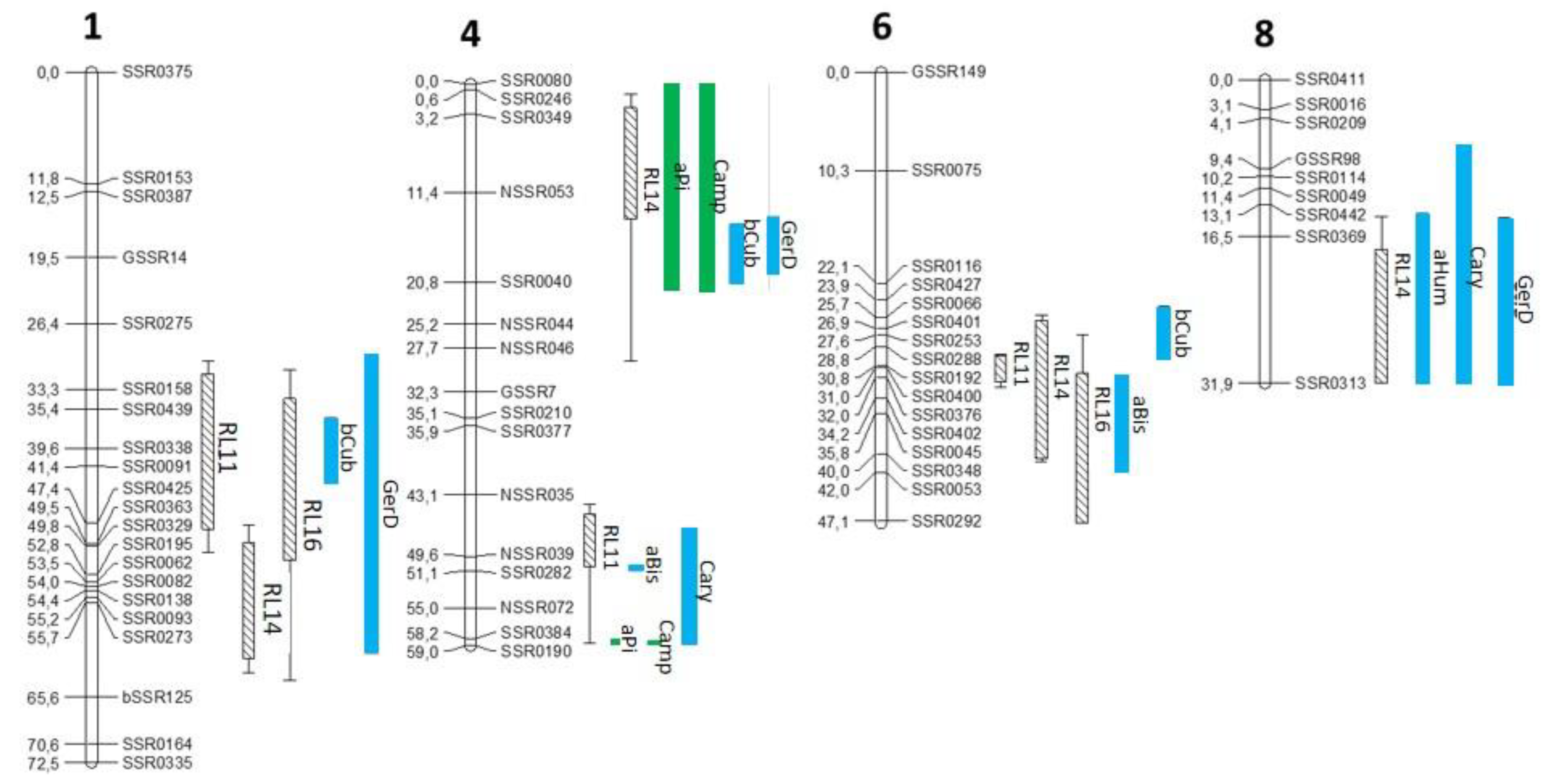
3.2. mQTL-rQTL Co-Localization Analysis Reveals Candidate Terpenes for Resistance to ALB
3.3. Differential Expression Analysis of Genes Underlying the Co-Localization Regions
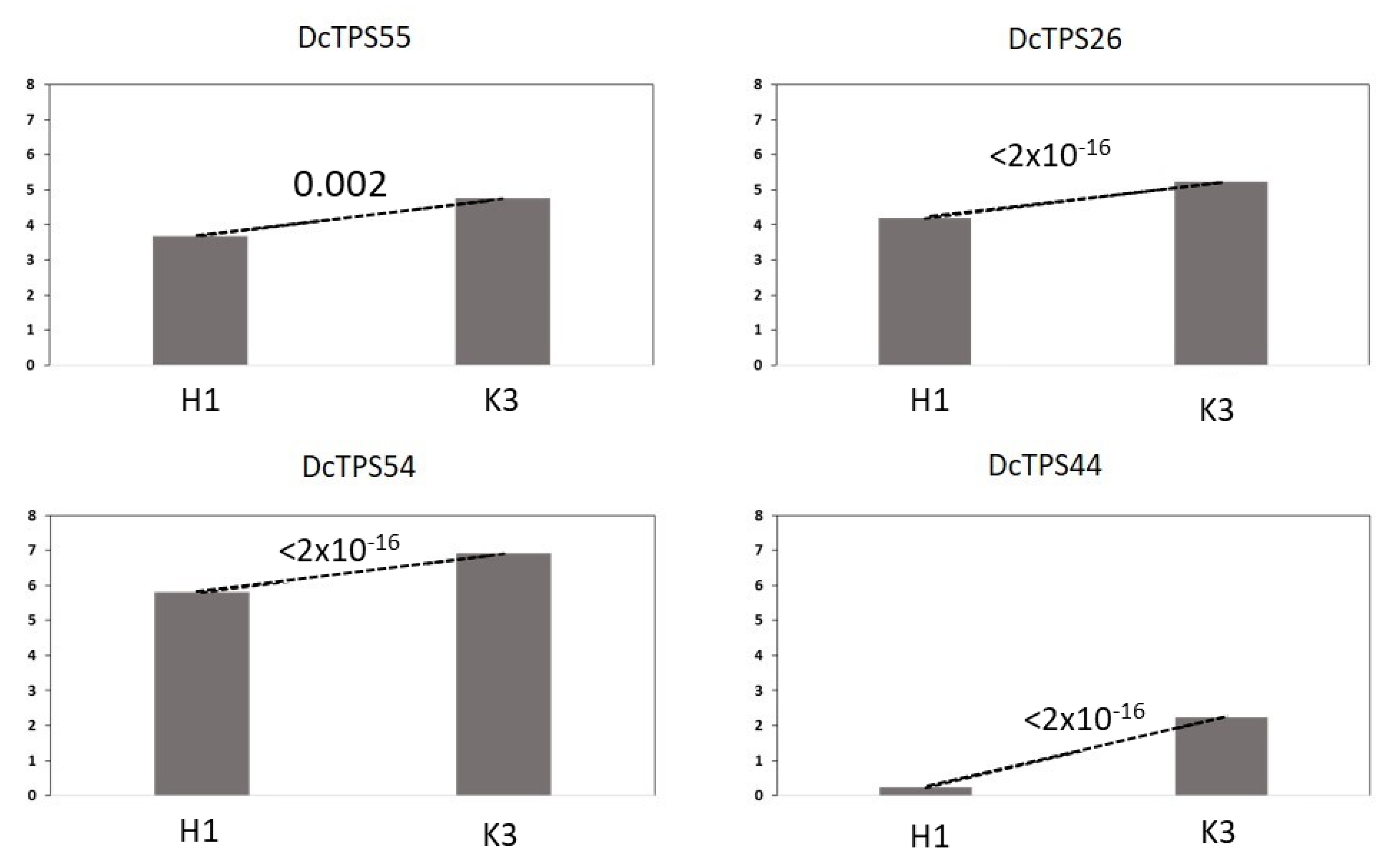
3.4. Further Analysis of TPS Genes Overexpressed in K3
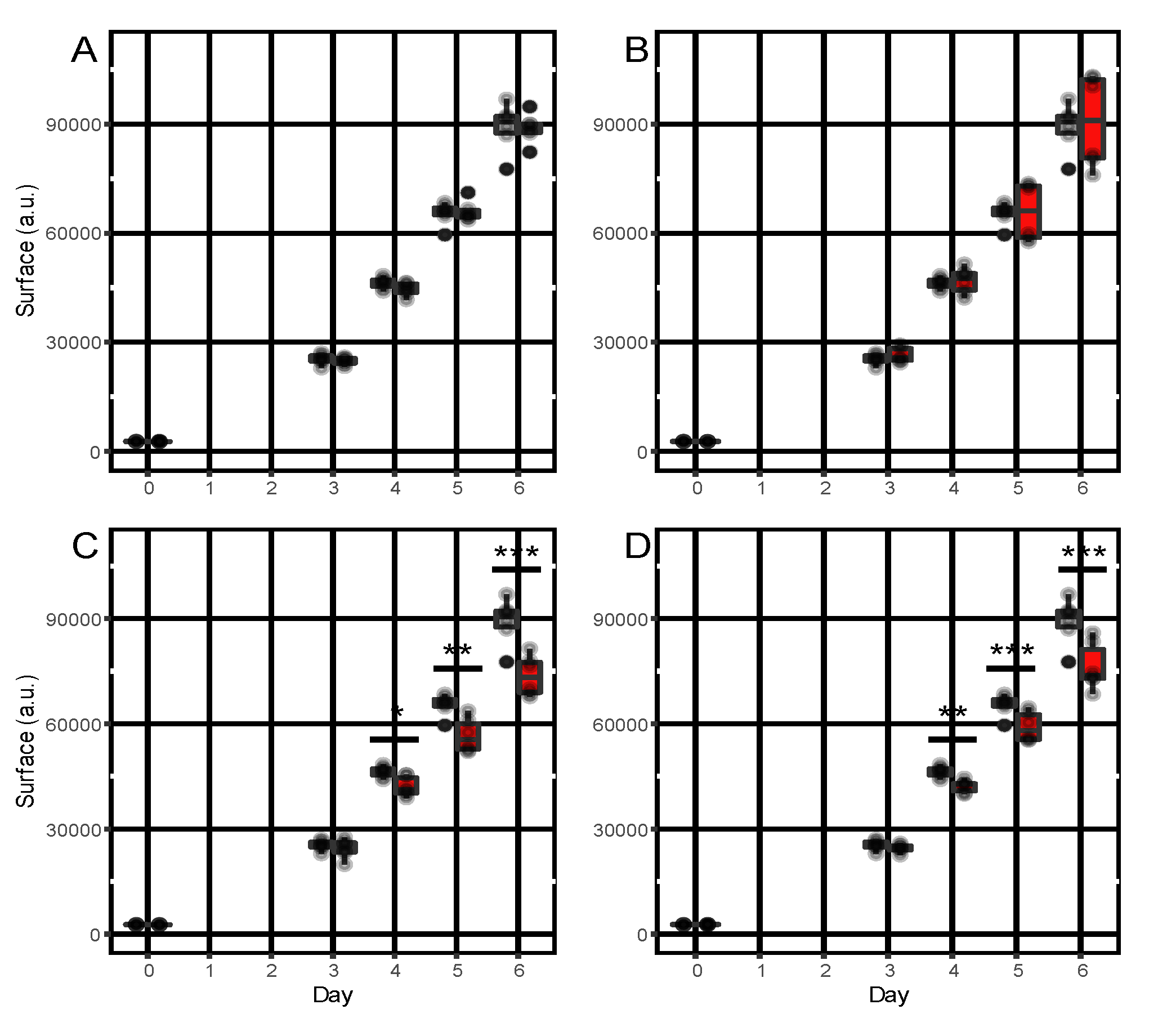
3.5. In Vitro Bioactivity of α-Pinene, Camphene, Caryophyllene and Humulene towards A. dauci
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Goodacre, R.; Vaidyanathan, S.; Dunn, W.B.; Harrigan, G.G.; Kell, D.B. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004, 22, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Biotechnol. Isoprenoids 2015, 148, 63–106. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Yeh, S. Isoprene emission from plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 407–436. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Gil, M.; Pontin, M.; Berli, F.; Bottini, R.; Piccoli, P. Metabolism of terpenes in the response of grape (Vitis vinifera L.) leaf tissues to UV-B radiation. Phytochemistry 2012, 77, 89–98. [Google Scholar] [CrossRef]
- Rodríguez, A.; Andrés, V.S.; Cervera, M.; Redondo, A.; Alquézar, B.; Shimada, T.; Gadea, J.; Rodrigo, M.; Zacarías, L.; Palou, L.; et al. The monoterpene limonene in orange peels attracts pests and microorganisms. Plant Signal. Behav. 2011, 6, 1820–1823. [Google Scholar] [CrossRef] [Green Version]
- Pontin, M.; Bottini, R.; Burba, J.L.; Piccoli, P. Allium sativum produces terpenes with fungistatic properties in response to infection with Sclerotium cepivorum. Phytochemistry 2015, 115, 152–160. [Google Scholar] [CrossRef]
- Abbas, F.; Ke, Y.; Yu, R.; Yue, Y.; Amanullah, S.; Jahangir, M.M.; Fan, Y. Volatile terpenoids: Multiple functions, biosynthesis, modulation and manipulation by genetic engineering. Planta 2017, 246, 803–816. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Verdonk, J.C.; Haring, M.A.; Van Tunen, A.J.; Schuurink, R.C. ODORANT1 regulates fragrance biosynthesis in petunia flowers. Plant Cell 2005, 17, 1612–1624. [Google Scholar] [CrossRef] [Green Version]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant volatile organic compounds evolution: Transcriptional regulation, epigenetics and polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef]
- Farrar, J.J.; Pryor, B.M.; Davis, R.M. Alternaria diseases of carrot. Plant Dis. 2004, 88, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Le Clerc, V.; Pawelec, A.; Birolleau-Touchard, C.; Suel, A.; Briard, M. Genetic architecture of factors underlying partial resistance to Alternaria leaf blight in carrot. Theor. Appl. Genet. 2009, 118, 1251–1259. [Google Scholar] [CrossRef]
- Le Clerc, V.; Marques, S.; Suel, A.; Huet, S.; Hamama, L.; Voisine, L.; Auperpin, E.; Jourdan, M.; Barrot, L.; Prieur, R.; et al. QTL mapping of carrot resistance to leaf blight with connected populations: Stability across years and consequences for breeding. Theor. Appl. Genet. 2015, 128, 2177–2187. [Google Scholar] [CrossRef]
- Koutouan, C.E.; Le Clerc, V.; Baltenweck, R.; Claudel, P.; Halter, D.; Hugueney, P.; Hamama, L.; Suel, A.; Huet, S.; Bouvet Merlet, M.H.; et al. Link between carrot leaf secondary metabolites and resistance to Alternaria dauci. Sci. Rep. 2018, 8, 13746. [Google Scholar] [CrossRef] [Green Version]
- Pawelec, A.; Dubourg, C.; Briard, M. Evaluation of carrot resistance to Alternaria leaf blight in controlled environments. Plant Pathol. 2006, 55, 68–72. [Google Scholar] [CrossRef]
- van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 1–15. Available online: http://www.biomedcentral.com/1471-2164/7/142 (accessed on 7 December 2022). [CrossRef] [Green Version]
- Le Clerc, V.; Aubert, C.; Cottet, V.; Yovanopoulos, C.; Piquet, M.; Suel, A.; Huet, S.; Koutouan, C.E.; Hamama, L.; Chalot, G.; et al. Breeding for carrot resistance to Alternaria dauci without compromising taste. Mol. Breed. 2019, 39, 59. [Google Scholar] [CrossRef]
- Jourjon, M.F.; Jasson, S.; Marcel, J.; Ngom, B.; Mangin, B. MCQTL: Multi-allelic QTL mapping in multi-cross design. Bioinformatics 2005, 21, 128–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Lovene, M.; Sanseverino, W.; Cavagnaro, P.; et al. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnot, S.; Tamby, J.P.; Martin-Magniette, M.L.; Bitton, F.; Taconnat, L.; Balzergue, S.; Aubourg, S.; Renou, J.P.; Lecharny, A.; Brunaud, V. CATdb: A public access to Arabidopsis transcriptome data from the URGV-CATMA platform. Nucleic Acids Res. 2007, 36, 986–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celton, J.M.; Gaillard, S.; Bruneau, M.; Pelletier, S.; Aubourg, S.; Martin-Magniette, M.L.; Navarro, L.; Laurens, F.; Renou, J.P. Widespread anti-sense transcription in apple is correlated with si RNA production and indicates a large potential for transcriptional and/or post-transcriptional control. New Phytol. 2014, 203, 287–299. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Copper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Boedo, C.; Benichou, S.; Berruyer, R.; Bersihand, S.; Dongo, A.; Simoneau, P.; Lecomte, M.; Briard, M.; Le Clerc, V.; Poupard, P. Evaluating aggressiveness and host range of Alternaria dauci in a controlled environment. Plant Pathology 2012, 61, 63–75. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Keilwagen, J.; Lehnert, H.; Berner, T.; Budahn, H.; Nothnagel, T.; Ulrich, D.; Dunemann, F. The terpene synthase gene family of carrot (Daucus carota L.): Identification of QTLs and candidate genes associated with terpenoid volatile compounds. Front. Plant Sci. 2017, 8, 1930. [Google Scholar] [CrossRef] [Green Version]
- Muchlinski, A.; Ibdah, M.; Ellison, S.; Yahyaa, M.; Nawade, B.; Laliberte, S.; Senalik, D.; Simon, P.; Whitehead, S.R.; Tholl, D. Diversity and function of terpene synthases in the production of carrot aroma and flavor compounds. Sci. Rep. 2020, 10, 9989. [Google Scholar] [CrossRef]
- Reichardt, S.; Budahn, H.; Lamprecht, D.; Riewe, D.; Ulrich, D.; Dunemann, F.; Kopertekh, L. The carrot monoterpene synthase gene cluster on chromosome 4 harbours genes encoding flavour-associated sabinene synthases. Hortic. Res. 2020, 7, 190. [Google Scholar] [CrossRef]
- Kearsey, M.J. The principles of QTL analysis (a minimal mathematics approach). J. Exp. Bot. 1998, 49, 1619–1623. [Google Scholar] [CrossRef]
- Keurentjes, J.J.; Fu, J.; De Vos, C.H.; Lommen, A.; Hall, R.D.; Bino, R.J.; van der Plas, L.H.W.; Jansen, R.C.; Vreugdenhil, D.; Koornneef, M. The genetics of plant metabolism. Nat. Genet. 2006, 38, 842–849. [Google Scholar] [CrossRef] [Green Version]
- Rowe, H.C.; Kliebenstein, D.J. Complex genetics control natural variation in Arabidopsis thaliana resistance to Botrytis cinerea. Genetics 2008, 180, 2237–2250. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Kwon, S.T.; Chen, F.; Kliebenstein, D.J. Isolate dependency of Brassica rapa resistance QTLs to Botrytis cinerea. Front. Plant Sci. 2016, 7, 161. [Google Scholar] [CrossRef] [Green Version]
- Gambliel, H.; Croteau, R. Pinene cyclases I and II. Two enzymes from sage (Salvia officinalis) which catalyze stereospecific cyclizations of geranyl pyrophosphate to monoterpene olefins of opposite configuration. J. Biol. Chem. 1984, 259, 740–748. [Google Scholar] [CrossRef]
- Huber, D.P.; Philippe, R.N.; Godard, K.A.; Sturrock, R.N.; Bohlmann, J. Characterization of four terpene synthase cDNAs from methyl jasmonate-induced Douglas-fir, Pseudotsuga menziesii. Phytochemistry 2005, 66, 1427–1439. [Google Scholar] [CrossRef]
- Matsuba, Y.; Nguyen, T.T.; Wiegert, K.; Falara, V.; Gonzales-Vigil, E.; Leong, B.; Schäfer, P.; Kudrna, D.; Wing, R.A.; Bolger, A.M.; et al. Evolution of a complex locus for terpene biosynthesis in Solanum. Plant Cell 2013, 25, 2022–2036. [Google Scholar] [CrossRef] [Green Version]
- Nützmann, H.W.; Osbourn, A. Gene clustering in plant specialized metabolism. Curr. Opin. Biotechnol. 2014, 26, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Schluttenhofer, C.; Yuan, L. Regulation of Specialized Metabolism by WRKY Transcription Factors. Plant Physiol. 2015, 167, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Spyropoulou, E.A.; Haring, M.A.; Schuurink, R.C. RNA sequencing on Solanum lycopersicum trichomes identifies transcription factors that activate terpene synthase promoters. BMC Genom. 2014, 15, 402. Available online: http://www.biomedcentral.com/1471-2164/15/402 (accessed on 7 December 2022). [CrossRef]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; San Andrés, V.; Cervera, M.; Redondo, A.; Alquézar, B.; Shimada, T.; Gadea, J.; Rodrigo, M.J.; Zacarias, L.; Palou, L.; et al. Terpene down-regulation in orange reveals the role of fruit aromas in mediating interactions with insect herbivores and pathogens. Plant Physiol. 2011, 156, 793–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, A.; Shimada, T.; Cervera, M.; Alquézar, B.; Gadea, J.; Gómez-Cadenas, A.; José De Ollas, C.; Rodrigo, M.J.; Zacarias, L.; Peña, L. Terpene down-regulation triggers defense responses in transgenic orange leading to resistance against fungal pathogens. Plant Physiol. 2014, 164, 321–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Sun, T.H.; Zhao, L.; Pan, X.W.; Lu, S. The bZIP transcription factor HY5 interacts with the promoter of the monoterpene synthase gene QH6 in modulating its rhythmic expression. Front. Plant Sci. 2015, 6, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuizen, N.J.; Chen, X.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural variation in monoterpene synthesis in kiwifruit: Transcriptional regulation of terpene synthases by NAC and ETHYLENE-INSENSITIVE3-like transcription. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, Y.; Shen, S.; Yin, X.; Klee, H.; Zhang, B.; Chen, K.; Hancock, R. Transcription factor CitERF71 activates the terpene synthase gene CitTPS16 involved in the synthesis of E-geraniol in sweet orange fruit. J. Exp. Bot. 2017, 68, 4929–4938. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.X.; Wang, L.J.; Zhao, B.; Shan, C.M.; Zhang, Y.H.; Chen, D.F.; Chen, X.Y. Progressive regulation of sesquiterpene biosynthesis in Arabidopsis and Patchouli (Pogostemon cablin) by the miR156-targeted SPL transcription factors. Mol. Plant 2015, 8, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antifungal activity of the components of Melaleuca alternifolia (tea tree) oil. J. Appl. Microbiol. 2003, 95, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Sati, S.C.; Sati, N.; Ahluwalia, V.; Walia, S.; Sati, O.P. Chemical composition and antifungal activity of Artemisia nilagirica essential oil growing in northern hilly areas of India. Nat. Prod. Res. 2013, 27, 45–48. [Google Scholar] [CrossRef]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The anticancer, antioxidant and antimicrobial properties of the sesquiterpene β-caryophyllene from the essential oil of Aquilaria crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef]
- Garcia-Rellán, D.; Verdeguer, M.; Salamone, A.; Blázquez, M.A.; Boira, H. Chemical composition, herbicidal and antifungal activity of Satureja cuneifolia essential oils from Spain. Nat. Prod. Commun. 2016, 11, 1934578X1601100636. [Google Scholar] [CrossRef] [Green Version]
- Feiner, A.; Pitra, N.; Matthews, P.; Pillen, K.; Wessjohann, L.A.; Riewe, D. Downy mildew resistance is genetically mediated by prophylactic production of phenylpropanoids in hop. Plant Cell Environ. 2021, 44, 323–338. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code of mQTL | Chr | 1-LOD SI (cM) | Max Position (cM) | R2 (%) | Global R2 (%) | Heritability H2 (%) | Additive Effect of Allele | |
|---|---|---|---|---|---|---|---|---|
| H1 | K3 | |||||||
| aPi | 4 t | 0–21.6 | 20.4 | 11.4 | 58.3 | 68 | −0.157 | 0.157 |
| 4 b | 58.3–58.7 | 58.7 | 42.9 | −0.388 | 0.388 | |||
| Camp | 4 t | 0–21.7 | 20.4 | 10.7 | 57.6 | 65 | −0.153 | 0.153 |
| 4 b | 58.4–58.7 | 58.7 | 42.5 | −0.389 | 0.389 | |||
| aBis | 4 b | 50.6–51 | 50.8 | 51.7 | 53.1 | 34 | −0.436 | 0.436 |
| 6 | 31.8–41.9 | 40.2 | 9.7 | −0.140 | 0.140 | |||
| aHum | 8 | 14.1–31.9 | 16.4 | 12.2 | 12.2 | 39 | 0.209 | −0.209 |
| bCub | 1 | 36.3–43.2 | 39.6 | 20.5 | 41.7 | 31 | −0.263 | 0.263 |
| 4 t | 14.7–20.8 | 16.4 | 26.9 | −0.273 | 0.273 | |||
| 6 | 24.6–30.1 | 28.9 | 12.6 | −0.167 | 0.167 | |||
| Cary | 4 b | 46.6–58.7 | 58.7 | 9.3 | 18.6 | 33.3 | −0.161 | 0.161 |
| 8 | 6.9–31.9 | 11.3 | 9.1 | 0.174 | −0.174 | |||
| GerD | 1 | 29.7–61 | 39.6 | 10 | 46 | 36.2 | −0.173 | 0.173 |
| 4 t | 13.9–19.8 | 16.4 | 26.4 | −0.271 | 0.271 | |||
| 8 | 14.5–31.9 | 21.4 | 18.7 | 0.235 | −0.235 | |||
| Chr | Located Genes | Genes Differentially Expressed between K3 and H1 | ||
|---|---|---|---|---|
| Underexpressed | Overexpressed | Related to Terpenes | ||
| 1 | 2181 | 207 | 206 | 5 |
| 4 t | 1229 | 76 | 92 | 2 |
| 4 b | 1718 | 154 | 196 | 10 |
| 6 | 577 | 63 | 45 | 3 |
| 8 | 1025 | 100 | 105 | 5 |
| Chr | Gene Name (Locus Number) | Function | Relative Expression | p-Value | |
|---|---|---|---|---|---|
| H1 | K3 | ||||
| 1 | bZIPHY5 (108204232) | TF | 3.94 | 4.48 | 0.004 |
| WRKY33 (108204915) | TF | 2.65 | 4.64 | <2 × 10−16 | |
| WRKY48 (108204668) | TF | 2.67 | 3.79 | <2 × 10−16 | |
| ERF71 (108206243) | TF | 0.98 | 0.15 | 0.0004 | |
| bZIP53 (108206338) | TF | 3.28 | 2.23 | <2 × 10−16 | |
| 4 t | NAC29 (108218926) | TF | 4.44 | 5.90 | 0.0004 |
| WRKY33 (108219317) | TF | 4.00 | 6.21 | <2 × 10−16 | |
| 4 b | Terpene synthase 10-like (DcTPS55;108217470) | TPS | 3.68 | 4.76 | 0.002 |
| Terpene synthase 10-like (DcTPS26;108217599) | TPS | 4.19 | 5.22 | <2 × 10−16 | |
| Terpene synthase 10-like (DcTPS54;108217617) | TPS | 5.80 | 6.92 | <2 × 10−16 | |
| β-bisabolene synthase-like (108216085) | TPS | 6.67 | 5.67 | 0.0004 | |
| WRKY7 (108215789) | TF | 5.39 | 4.74 | 0.0008 | |
| ERF054 (108216387) | TF | 3.45 | 2.1 | <2 × 10−16 | |
| NAC2 (108215781) | TF | 4.51 | 5.26 | 0.0004 | |
| ERF4 (108217832) | TF | 1.87 | 2.87 | <2 × 10−16 | |
| bZIP17 (108218833) | TF | 3.98 | 4.76 | 0.0004 | |
| bZIP27 (108217633) | TF | 3.60 | 0.51 | <2 × 10−16 | |
| 6 | bZIP61 (108225065) | TF | 4.28 | 4.89 | 0.007 |
| ERF4 (108225207) | TF | 0.82 | 3.29 | 0.002 | |
| SPL1 (108224238) | TF | 3.98 | 2.84 | <2 × 10−16 | |
| 8 | α-farnesene synthase-like(DcTPS44; 108198720) | TPS | 0.23 | 2.23 | <2 × 10−16 |
| AP2/ERF (108198780) | TF | 0.92 | 5.22 | <2 × 10−16 | |
| ERF1B-like (108198802) | TF | 1.35 | 3.21 | <2 × 10−16 | |
| MYB (108197621) | TF | 0.74 | 0.19 | 0.005 | |
| ERF_like (108197006) | TF | 4.38 | 2.66 | <2 × 10−16 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koutouan, C.E.; Le Clerc, V.; Suel, A.; Hamama, L.; Claudel, P.; Halter, D.; Baltenweck, R.; Hugueney, P.; Chich, J.-F.; Moussa, S.A.; et al. Co-Localization of Resistance and Metabolic Quantitative Trait Loci on Carrot Genome Reveals Fungitoxic Terpenes and Related Candidate Genes Associated with the Resistance to Alternaria dauci. Metabolites 2023, 13, 71. https://doi.org/10.3390/metabo13010071
Koutouan CE, Le Clerc V, Suel A, Hamama L, Claudel P, Halter D, Baltenweck R, Hugueney P, Chich J-F, Moussa SA, et al. Co-Localization of Resistance and Metabolic Quantitative Trait Loci on Carrot Genome Reveals Fungitoxic Terpenes and Related Candidate Genes Associated with the Resistance to Alternaria dauci. Metabolites. 2023; 13(1):71. https://doi.org/10.3390/metabo13010071
Chicago/Turabian StyleKoutouan, Claude Emmanuel, Valérie Le Clerc, Anita Suel, Latifa Hamama, Patricia Claudel, David Halter, Raymonde Baltenweck, Philippe Hugueney, Jean-François Chich, Sitti Anlati Moussa, and et al. 2023. "Co-Localization of Resistance and Metabolic Quantitative Trait Loci on Carrot Genome Reveals Fungitoxic Terpenes and Related Candidate Genes Associated with the Resistance to Alternaria dauci" Metabolites 13, no. 1: 71. https://doi.org/10.3390/metabo13010071
APA StyleKoutouan, C. E., Le Clerc, V., Suel, A., Hamama, L., Claudel, P., Halter, D., Baltenweck, R., Hugueney, P., Chich, J. -F., Moussa, S. A., Champlain, C., Huet, S., Voisine, L., Pelletier, S., Balzergue, S., Chevalier, W., Geoffriau, E., & Briard, M. (2023). Co-Localization of Resistance and Metabolic Quantitative Trait Loci on Carrot Genome Reveals Fungitoxic Terpenes and Related Candidate Genes Associated with the Resistance to Alternaria dauci. Metabolites, 13(1), 71. https://doi.org/10.3390/metabo13010071