
Comparative Metabolomic Profiling of Horse Gram (Macrotyloma uniflorum (Lam.) Verdc.) Genotypes for Horse Gram Yellow Mosaic Virus Resistance

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Screening for HgYMV Resistance
2.2. Grouping of Horse Gram Genotypes
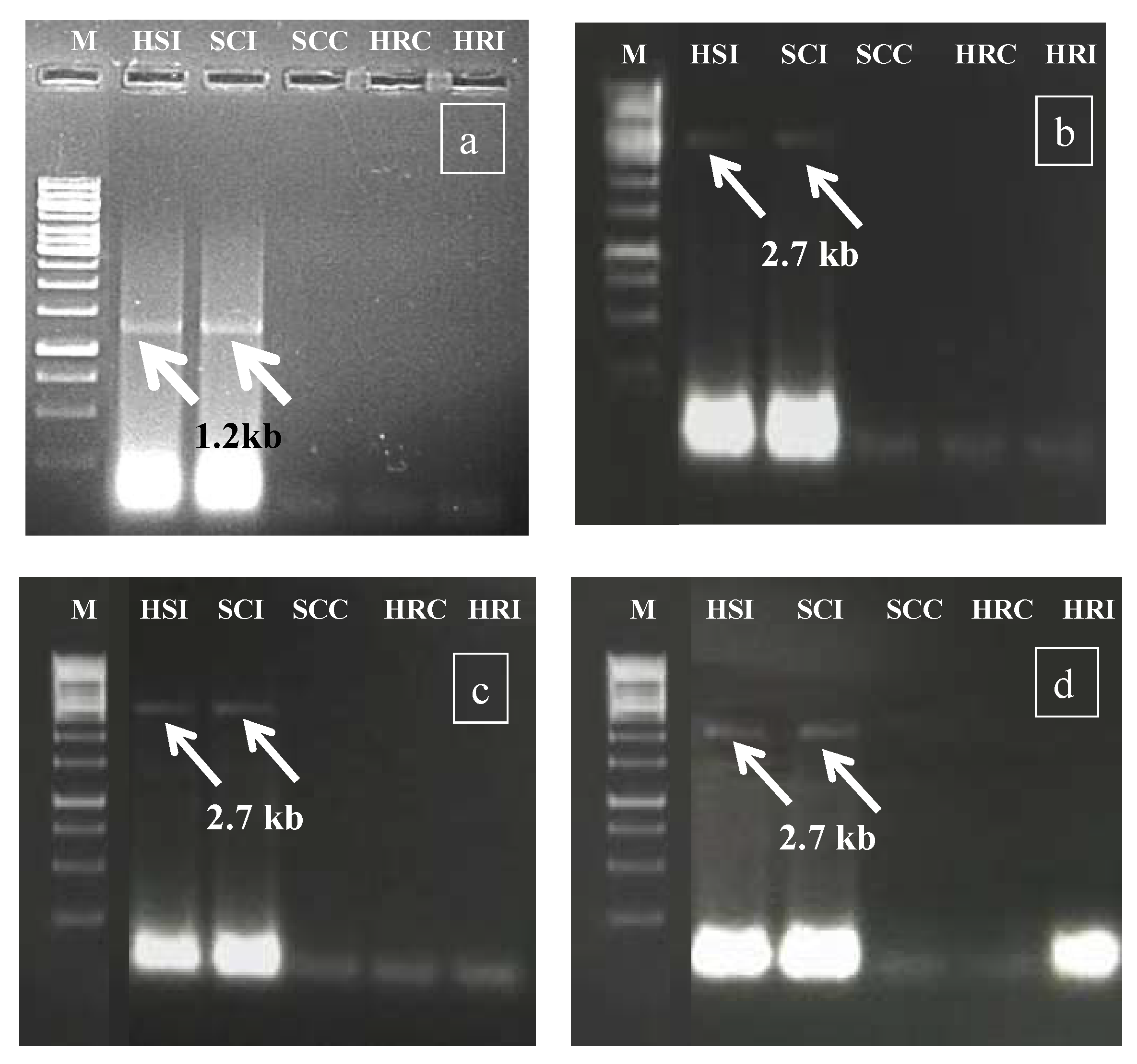
2.3. Confirmation of HgYMV Infection
2.4. Metabolome Analysis
Preparation of Sample and Extraction of Metabolites
2.5. GC/MS Analysis
2.6. PCA and HCA Analysis
3. Results
3.1. HgYMV Infection and Symptom Development
3.2. Confirmation of HgYMV Infection
3.3. Genotype Categorization and Confirmation of HgYMV
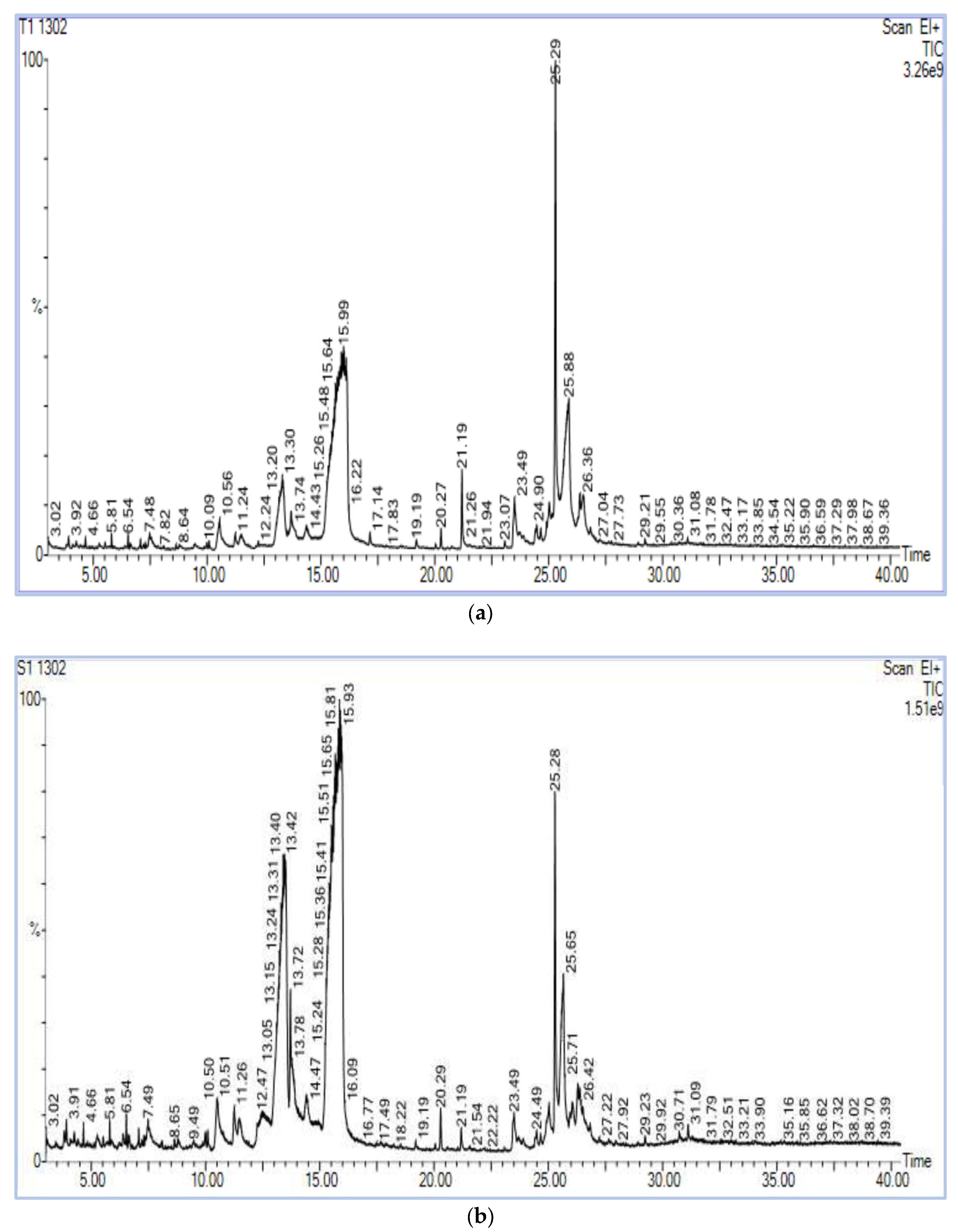
3.4. GC–MS Chromatography
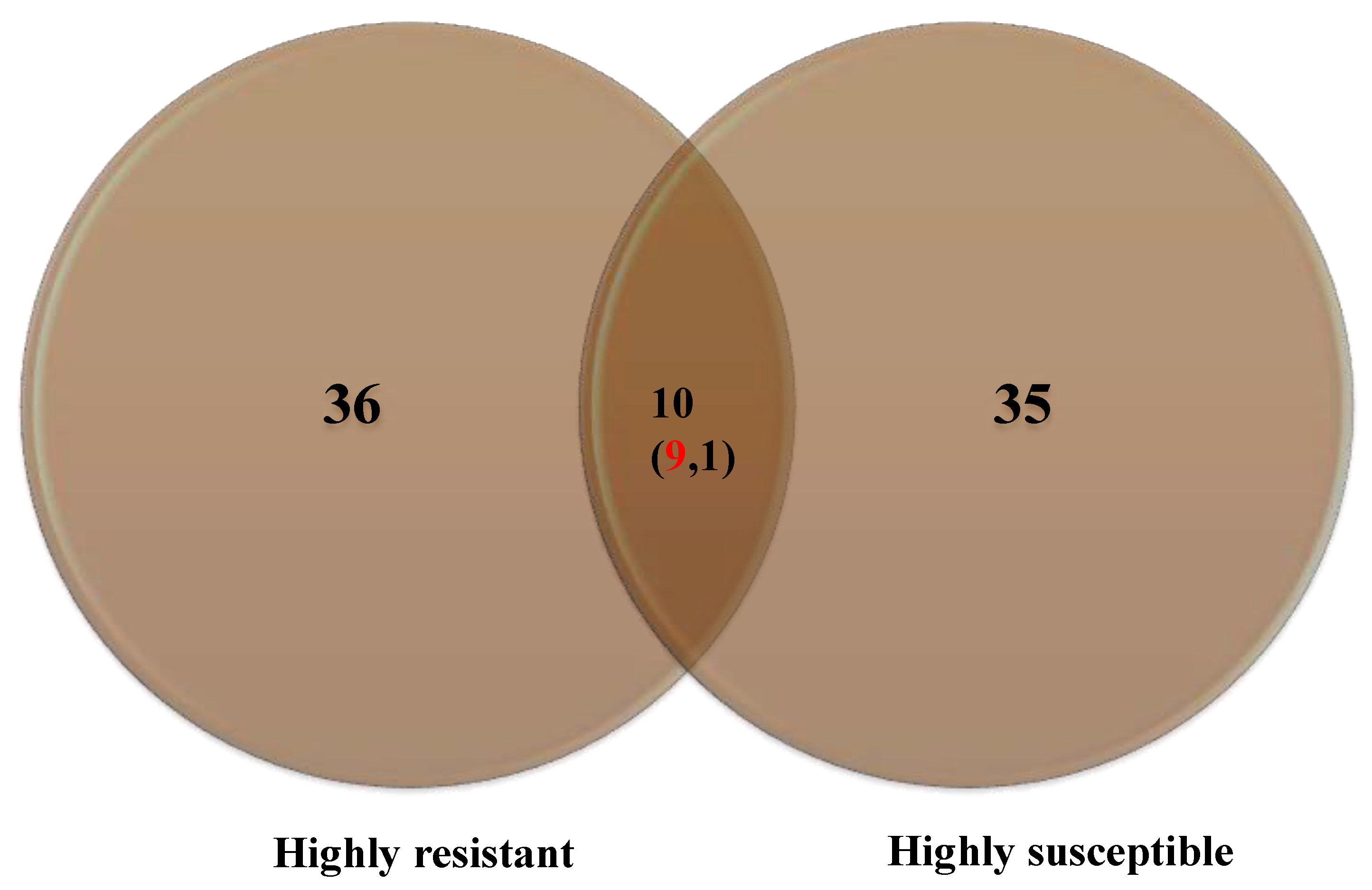
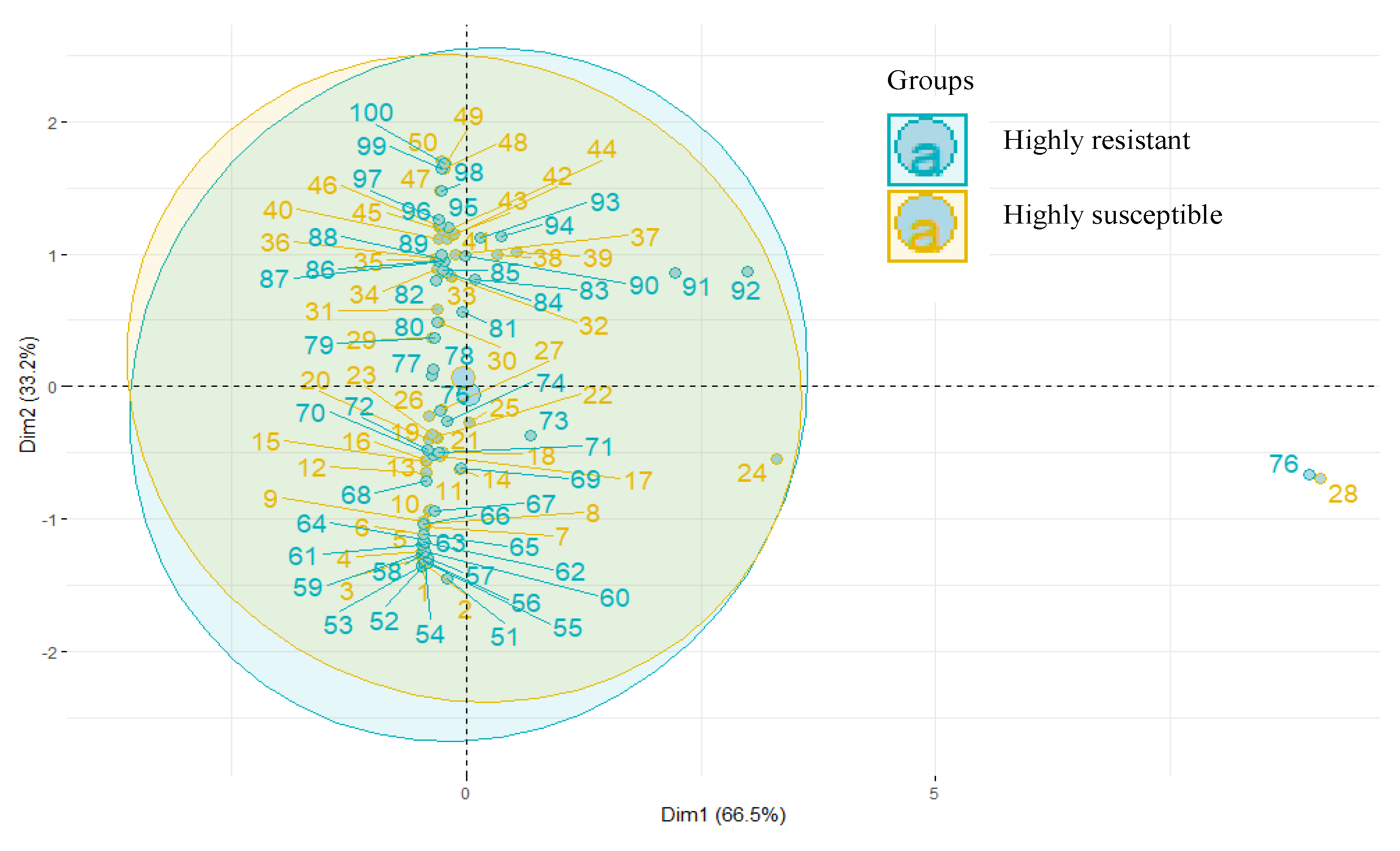
3.5. PCA and HCA
4. Discussion
4.1. HgYMV Screening, Categorization of Genotypes, and Confirmative Studies
4.2. GC/MS Analysis, PCA, and HCA
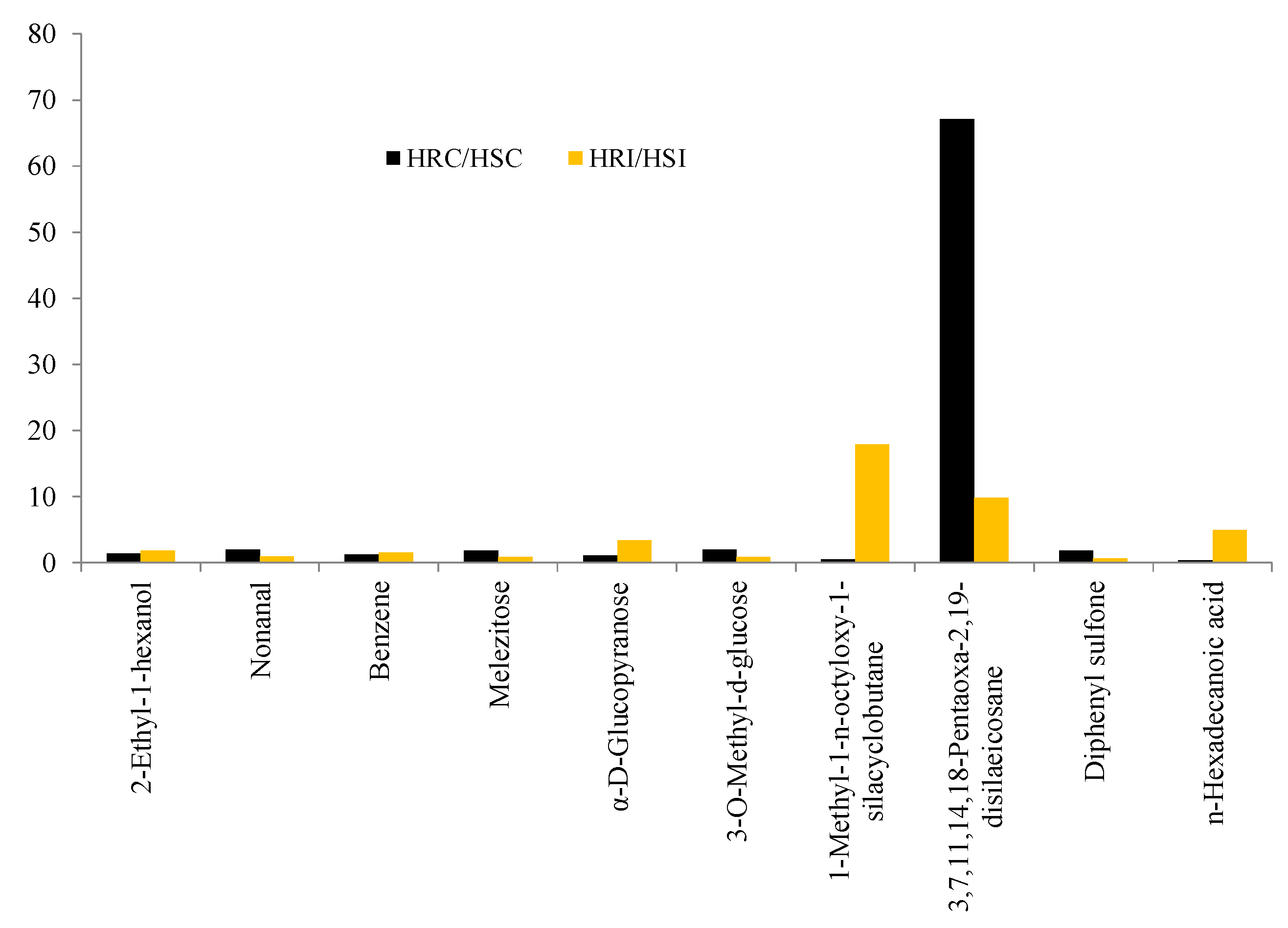
4.3. The Biological Significance of Biomolecules Accumulated in the HR Genotype PLS6002
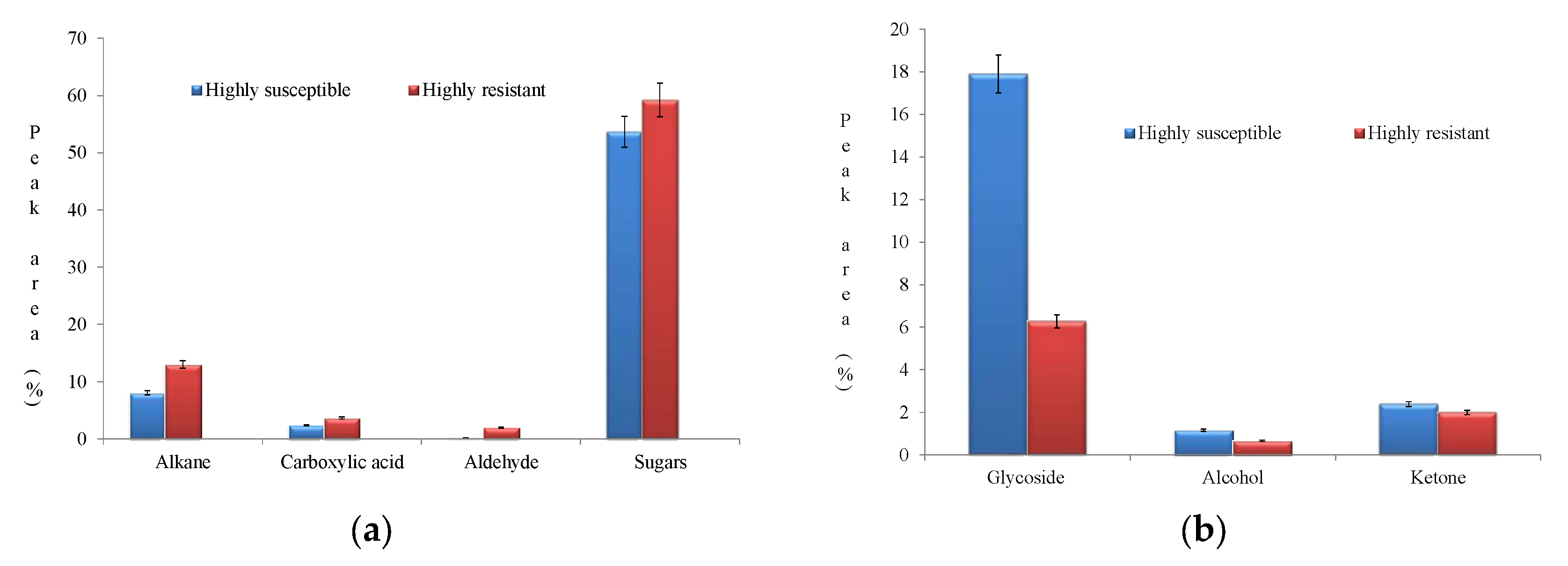
4.3.1. Unique Biomolecules
4.3.2. Other Significant Biomolecules: Sugars, Alkanes and Carboxylic Acids
4.4. The Biological Significance of Accumulation of Biomolecules in the HS Genotype PLS6194
Ketones, Glycosides, and Alcohols
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, A.; Contador, C.A.; Fan, K.; Lam, H.M. Interaction and regulation of carbon, nitrogen, and phosphorus metabolisms in root nodules of legumes. Front. Plant Sci. 2018, 9, 1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Geraldo, R.; Santos, C.S.; Pinto, E.; Vasconcelos, M.W. Widening the Perspectives for Legume Consumption: The Case of Bioactive Non-nutrients. Front. Plant Sci. 2022, 13, 772054. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Xu, B. Polyphenol-rich lentils and their health promoting effects. Int. J. Mol. Sci. 2017, 18, 2390. [Google Scholar] [CrossRef] [Green Version]
- Iannetta, P.P.M.; Hawes, C.; Begg, G.S.; Maaß, H.; Ntatsi, G.; Savvas, D.; Vasconcelos, M.; Hamann, K.; Williams, M.; Styles, D.; et al. A multifunctional solution for wicked problems: Value-chain wide facilitation of legumes cultivated at bioregional scales is necessary to address the climate-biodiversity-nutrition nexus. Front. Sust. Food Syst. 2021, 5, 692137. [Google Scholar] [CrossRef]
- Chahota, R.K.; Sharma, T.R.; Sharma, S.K.; Kumar, N.; Rana, J.C. Horsegram. In Genetic and Genomic Resources of Grain Legume Improvement; Singh, M., Upadhyaya, H.D., Bisht, I.S., Eds.; Elsevier: Oxford, UK, 2013; Chapter 12; pp. 293–305. [Google Scholar]
- Patel, D.P.; Dabas, B.S.; Sapra, R.S.; Mandal, S. Evaluation of Horsegram (Macrotyloma uniflorum Lam.) Germplasm; National Bureau of Plant Genetic Resources: New Delhi, India, 1995; pp. 37–53. [Google Scholar]
- Gopalan, C.; Rama Sastri, B.V.; Balasubramanian, S.C. Nutritive Value of Indian Foods; Indian Council of Medical Research (ICMR): New Delhi, India; National Institute of Nutrition: Hyderabad, India, 1989; pp. 110–125. [Google Scholar]
- Sodani, S.N.; Paliwal, R.V.; Jain, L.K. Phenotypic stability for seed yield in rainfed Horsegram (Macrotyloma uniflorum [Lam.] Verdc. In Proceedings of the National Symposium on Arid Legumes for Sustainable Agriculture and Trade, Jodhpur, India, 5–7 November 2004; Central Arid Zone Research Institute: Jodhpur, India, 2004; pp. 5–7. [Google Scholar]
- Murthy, S.M.; Devaraj, V.R.; Anitha, P.; Tejavathi, D.H. Studies on the activities of antioxidant enzymes under induced drought stress in vivo and in vitro plants of Macrotyloma uniflorum [Lam.] Verdc. Recent Res. Sci. Technol. 2012, 4, 34–37. [Google Scholar]
- Chaitanya, D.A.K.; Kumar, S.; Reddy, M.; Mukherjee, A.M.; Sumanth, N.S.V.; Ramesh, D.A.; Chaitanya, D.A.; Kumar, M.S.; Bommi-Reddy, A.; Mukherjee, N.; et al. Anti urolithiatic activity of Macrotyloma uniflorum seed extract on ethylene glycol induced urolithiasis in albino rats. Int. J. Innov. Pharm. Sci. Res. 2010, 1, 216–226. [Google Scholar]
- Sudhagar, R.; Priyanka, S.; Chockalingam, V.; Sendhilvel, V.; Souframanien, J.; Raja, K.; Kanagarajan, S. Development and Metabolic Characterization of Horse Gram (Macrotyloma uniflorum Lam. (Verdc.)) Mutants for Powdery Mildew Resistance. Agronomy 2022, 12, 800. [Google Scholar] [CrossRef]
- Malathi, V.G.; John, P. Gemini viruses infecting legumes. In Characterization, Diagnosis & Management of Plant Viruses, Volume 3: Vegetables and Pulse Crops; Rao, G.P., Lava Kumar, P., Holguin-Pena, R.J., Eds.; Stadium Press LLC: Houston, TX, USA, 2008; pp. 97–123. [Google Scholar]
- Barnabas, A.D.; Radhakrishnan, G.K.; Ramakrishnan, U. Characterization of a begomovirus causing horsegram yellow mosaic disease in India. Eur. J. Plant Pathol. 2010, 127, 41–51. [Google Scholar] [CrossRef]
- Bashir, M.; Ahmad, Z.; Mansoor, S. Occurance and distribution of viral diseases of mungbean and mashbean in Punjab, Pakistan. Pak. J. Bot. 2006, 38, 1341–1351. [Google Scholar]
- Sharma, S.R.; Varma, A. Control of yellow mosaic of mungbean through insecticides and oil. J. Entomol. Res. 1982, 6, 130–136. [Google Scholar]
- Alam, A.M.; Somta, P.; Jompuk, C.; Chatwachirawong, P.; Srinives, P. Evaluation of mungbean genotypes based on yield stability and reaction to mungbean yellow mosaic virus disease. Plant Pathol. J. 2014, 30, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Muniyappa, V.; Reddy, H.R. Studies on the yellow mosaic disease of horsegram (Dolichos biflorus L.)—Virus vector relationships. Mysore J. Agric. Sci. 1976, 10, 605–610. [Google Scholar]
- Qazi, J.; Ilyas, M.; Mansoor, S.; Briddo, R.W. Legume yellow mosaic viruses: Genetically isolated begomoviruses. Mol. Plant Pathol. 2007, 8, 343–348. [Google Scholar] [CrossRef]
- Parimala, K.; Meenakumari, K.V.S.; Sudhakar, R.; Durga, K.K. Screening of horsegram genotypes against yellow mosaic virus and powdery mildew diseases. Indian J. Pl. Protect. 2011, 39, 160. [Google Scholar]
- Durga, K.K.; Varma, V.S.; Reddy, A.V.V. Sources of resistance to wilt and YMV in horse gram. J. Glob. Biosci. 2014, 3, 280–284. [Google Scholar]
- Bandi, H.R.K.; Rao, K.N.; Vamsi Krishna, K.; Srinivasulu, K. Screening of blackgram (Vigna mungo l. Hepper) germplasm for resistance to mungbean yellow mosaic virus under rice fallow situation. Bull. Environ. Pharmacol. Life Sci. 2018, 7, 125–128. [Google Scholar]
- Warth, B.; Parich, A.; Bueschl, C.; Schoefbeck, D.; Neumann, N.K.N.; Kluger, B.; Schuster, K.; Krska, R.; Adam, G.; Lemmens, M.; et al. GC–MS based targeted metabolic profiling identifies changes in the wheat metabolome following deoxynivalenol treatment. Metabolomics 2015, 11, 722–738. [Google Scholar] [CrossRef] [Green Version]
- Maravi, D.K.; Kumar, S.; Sahoo, L. NMR-Based Metabolomic Profiling of Mungbean Infected with Mungbean Yellow Mosaic India Virus. Appl. Biochem. Biotechnol. 2022, 194, 5808–5826. [Google Scholar] [CrossRef]
- Vinci, G.; Cozzolino, V.; Mazzei, P.; Monda, H.; Spaccini, R.; Piccol, A. An alternative to mineral phosphorus fertilizers: The combined effects of Trichoderma harzianum and compost on Zea mays, as revealed by 1 H NMR and GC-MS metabolomics. PLoS ONE 2018, 13, e0209664. [Google Scholar] [CrossRef] [Green Version]
- Alice, D.; Nadarajan, N. Pluses: Screening Techniques and Assessment for Disease Resistance; All India Coordinated Research Project on MULLaRP; Tamil Nadu Agricultural University: Coimbatore, India; Kasturi Graphics and Printers: Coimbatore, India, 2007. [Google Scholar]
- Wheeler, B.E.J. An Introduction to Plant Disease; John Wiley: London, UK, 1969; p. 301. [Google Scholar]
- Rouhibakhsh, A.; Priya, J.; Periasamy, M.; Haq, Q.M.I.; Malathi, V.G. An improved DNA isolation method and PCR protocol for efficient detection of multicomponents of begomovirus in legumes. J. Virol. Methods 2008, 147, 37–42. [Google Scholar] [CrossRef]
- Rojas, M.R.; Gilbertson, R.L.; Russell, D.R.; Maxwell, D.P. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis. 1993, 77, 340–347. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Scalabrin, E.; Radaelli, M.; Rizzato, G.; Bogani, P.; Buiatti, M.; Gambaro, A.; Capodaglio, G. Metabolomic analysis of wild and transgenic Nicotiana langsdorffii plants exposed to abiotic stresses: Unraveling metabolic responses. Anal. Bioanal. Chem. 2015, 407, 6357–6368. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Sawada, Y.; Nakabayashi, R.; Sato, S.; Hirakawa, H.; El-Sayed, M.; Hirai, M.Y.; Saito, K.; Yamauchi, N.; Shigyo, M. Integrating transcriptome and target metabolome variability in doubled haploids of Allium cepa for abiotic stress protection. Mol. Breed. 2015, 35, 195. [Google Scholar] [CrossRef] [Green Version]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmuller, E.; Dormann, P.; Gibon, Y.; Stitt, M.; Willmitzer, L.; et al. GMD@CSBDB: The Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Prema, G.U.; Rangaswamy, K.T. Field Evaluation of horsegram germplasm/ genotypes against horsegram yellow mosaic virus (HgYMV) disease and biological transmission of horse gram yellow mosaic virus to different leguminous hosts through white flies. Int. J. Agric. Sci. 2017, 9, 4934–4939. [Google Scholar]
- Kaushik, U.; Aeri, V.; Mir, S.R. Cucurbitacins—An insight into medicinal leads from nature. Pharmacogn. Rev. 2015, 9, 12–18. [Google Scholar]
- Metcalf, R.L.; Metcalf, R.A.; Rhodes, A.M. Cucurbitacins as kairomones for diabroticite beetles. Proc. Natl. Acad. Sci. USA 1980, 77, 3769–3772. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, R.Y.; Vaagenes, P.; Breivik, T.; Fonnum, F.; Opstad, P.K. Glycine—An important neurotransmitter and cytoprotective agent. Acta Anaesthesiol Scand. 2005, 49, 1108–1116. [Google Scholar] [CrossRef]
- Hulbert, P.B.; Beuding, E.; Robinson, C.H. Structure and antisehistosomal activity in the nitrofuran series. J. Med. Chem. 1973, 16, 72. [Google Scholar] [CrossRef]
- Moghayedi, M.; Ahmadzadeh, H.; Ghazvini, K.H.; Goharshadi, E.K. Neglected antibacterial activity of ethylene glycol as a common solvent. Microb. Pathog. 2017, 107, 457–461. [Google Scholar] [CrossRef]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two classes of plant antibiotics: Phytoalexins versus “phytoanticipins”. Plant Cell 1994, 6, 1191–1192. [Google Scholar] [CrossRef]
- Wittstock, U.; Burow, M. Glucosinolate breakdown in Arabidopsis: Mechanism, regulation and biological significance. Arab. Book 2010, 8, e0134. [Google Scholar] [CrossRef] [Green Version]
- Tierens, K.F.; Thomma, B.P.; Brouwer, M.; Schmidt, J.; Kistner, K.; Porzel, A.; Mauch-Mani, B.; Cammue, B.P.; Broekaert, W.F. Study of the role of antimicrobial glucosinolate-derived isothiocyanates in resistance of Arabidopsis to microbial pathogens. Plant Physiol. 2001, 125, 1688–1699. [Google Scholar] [CrossRef] [Green Version]
- Bhat, Z.S.; Rather, M.A.; Maqbool, M.; Hafiz, U.L.L.; Yousuf, S.K.; Ahmad, Z. α-pyrones: Small molecules with versatile structural diversity reflected in multiple pharmacological activities-an update. Biomed. Pharmacother. 2017, 91, 265–277. [Google Scholar] [CrossRef]
- Ahmad, I.; Shagufta. Sulfones: An important class of organic compounds with diverse biological activities. Int. J. Pharm. Pharm. Sci. 2015, 7, 19–27. [Google Scholar]
- Almajan, G.L.; Barbuceanu, S.F.; Almajan, E.R.; Draghici, C.; Saramet, G. Synthesis, characterization and antibacterial activity of some triazole Mannich bases carrying diphenylsulfone moieties. Eur. J. Med. Chem. 2009, 44, 3083–3089. [Google Scholar] [CrossRef]
- Belyagoubi-Benhammou, N.; Belyagoubi, L.; Gismondi, A.; Di Marco, G.; Canini, A.; Bekkara, F.A. GC/MS analysis, and antioxidant and antimicrobial activities of alkaloids extracted by polar and apolar solvents from the stems of Anabasis articulata. Med. Chem. Res. 2019, 28, 754–767. [Google Scholar] [CrossRef]
- Singh, A.; Palariya, D.; Dhami, A.; Prakash, O.; Kumar, R.; Rawat, D.S.; Pant, A.K. Biological activities and phytochemical analysis of Zanthoxylum armatum dc. leaves and bark extracts collected from Kumaun region, Uttarakhand, India. J. Med. Herbs Ethnomed. 2020, 6, 1–10. [Google Scholar] [CrossRef]
- Patnaik, B.R.; Rout, D.N.; Mahapatra, G.N. Studies of oxazole part I: Synthesis and chlorination of some new 2-amino-4-substituted oxazoles and their use as fungicides. J. Indian Chem. Soc. 1978, 55, 264. [Google Scholar] [CrossRef]
- Hussein, H.M.; Ubaid, J.M.; Hameed, I.H. Insecticidal Activity of Methanolic Seeds Extract of Ricinus communis on Adults of Callosobruchus maculatus (Coleoptera: Brauchidae) and Analysis of its Phytochemical Composition. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 1385–1397. [Google Scholar]
- Morkunas, I.; Gmerek, J. Sucrose-stimulated accumulation of isoflavonoids as a defence response of lupine to Fusarium oxysporum. Plant Physiol. Bioch. 2005, 43, 363–373. [Google Scholar] [CrossRef]
- Bolouri-Moghaddam, M.R.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Herbers, K.; Meuwly, P.; Frommer, W.B.; Métraux, J.P.; Sonnewald, U. Systemic acquired resistance mediated by the ectopic expression of invertase: Possible hexose sensing in the secretory pathway. Plant Cell 1996, 8, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Faivre-Rampant, O.; Thomas, J.; Allègre, M.; Morel, J.B.; Tharreau, D.; Nottéghem, J.L.; Lebrun, M.H.; Schaffrath, U.; Piffanelli, P. Characterization of the model system rice-Magnaporthefor the study of nonhost resistance in cereals. New Phytol. 2008, 180, 899–910. [Google Scholar] [CrossRef]
- Kano, A.; Gomi, K.; Yamasaki-Kokudo, Y.; Satoh, M.; Fukumoto, T.; Ohtani, K.; Tajima, S.; Izumori, K.; Tanaka, K.; Ishida, Y.; et al. A rare sugar, d-allose, confers resistance to rice bacterial blight with upregulation of defense-related genes in Oryza sativa. Phytopathology 2010, 100, 85–90. [Google Scholar] [CrossRef]
- Samuels, L.; Kunst, L.; Jetter, R. Sealing plant surfaces: Cuticular wax formation by epidermal cells. Annu. Rev. Plant Bio. 2008, 59, 683–707. [Google Scholar] [CrossRef] [Green Version]
- Baena-González, E.; Sheen, J. Convergent energy and stress signalling. Trends Plant Sci. 2008, 13, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Dracheva, L.V.; Dorozhko, E.V.; Avramchuk, O.A.; Korotkova, E.I.; Ryzhkova, E.P.; Hao, L.; Danilova, I.V. Voltammetric study of the antioxidant activity of propionic acid bacteria in liquid cultures. Mosc. Univ. Biol. Sci. Bull. 2009, 64, 157–160. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, P.; Marwaha, R.K.; Narasimhan, B. Synthesis, antimicrobial evaluation and QSAR studies of propionic acid derivatives. Arab. J. Chem. 2017, 10, S881–S893. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Deng, S.; Wang, Y.; Li, P.; Tang, L.; Pang, Y. Specific inhibition of acyl-coa oxidase-1 by an acetylenic acid improves hepatic lipid and reactive oxygen species (ROS) metabolism in rats fed a high fat diet. J. Biol. Chem. 2017, 292, 3800–3809. [Google Scholar] [CrossRef] [Green Version]
- Kasuga, N.C.; Sugie, A.; Nomiya, K. Syntheses, structures and antimicrobial activities of water-soluble silver(i)–oxygen bonding complexes with chiral and racemic camphanic acid (HCA) ligands. Dalton Trans. 2004, 21, 3732–3740. [Google Scholar] [CrossRef]
- Seidel, V.; Taylor, P.W. In vitro activity of extracts and constituents of Pelagonium against rapidly growing mycobacteria. Int. J. Antimicrob. Agents 2004, 23, 613–619. [Google Scholar] [CrossRef]
- Agoramoorthy, G.; Chandrasekaran, M.; Venkatesalu, V.; Hsu, M.J. Antibacterial and antifungal activities of fatty acid methyl esters of the blind-your-eye mangrove from India. Braz. J. Microbiol. 2007, 38, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Kravchuk, Z.; Vicedo, B.; Flors, V.; Camañes, G.; González-Bosch, C.; García-Agustín, P. Priming for JA-dependent defenses using hexanoic acid is an effective mechanism to protect Arabidopsis against B. cinerea. J. Plant Physiol. 2011, 168, 359–366. [Google Scholar] [CrossRef]
- Kumar, P.P.; Rao, M.R.K.; Elizabeth, A.A.; Prabhu, K. GC MS analysis of one ayurvedic medicine sahacharadi kashayam. Int. J. Pharm. 2018, 10, 31214–31230. [Google Scholar]
- Radhakrishnan, R.; Lee, I.J. Penicillium–sesame interactions: A remedy for mitigating high salinity stress effects on primary and defense metabolites in plants. Environ. Exp. Bot. 2015, 116, 47–60. [Google Scholar] [CrossRef]
- McManus, M.C. Mechanisms of bacterial resistance to antimicrobial agents. AM J. Health Syst. Ph. 1997, 54, 1420–1433. [Google Scholar] [CrossRef]
- Chakraborty, A.; Brantne, A.; Mukainaka, T.; Nobukuni, Y.; Kuchide, M.; Konoshima, T.; Tokuda, H.; Nishino, H. Cancer chemopreventive activity of Achyranthes aspera leaves on Epstein–Barr virus activation and two-stage mouse skin carcinogenesis. Cancer Lett. 2002, 177, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bellincampi, D.; Cervone, F.; Lionetti, V. Plant cell wall dynamics and wall-related susceptibility in plant–pathogen interactions. Front. Plant Sci. 2014, 5, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Disease Symptom | Category |
|---|---|---|
| 0 | No plants showing symptoms. | Immune |
| 1 | No visible symptoms or minute yellow specks on leaves. | Free |
| 2 | Small yellow spots with restricted spread covering 0.1% to 5% leaf area. | Highly resistant (HR) |
| 3 | Mottling of leaves covering 6% to 10% leaf area. | Resistant (R) |
| 4 | Yellow of mottling leaves covering 11% to 15% leaf area. | Moderately resistant (MR) |
| 5 | Yellow of mottling leaves covering 15% to 20% leaf area. | Moderately susceptible (MS) |
| 6 | Yellow coloration of 21%–30% leaves and yellow pods. | Susceptible (S) |
| 7 | Pronounced yellow mottling and discoloration of leaves and pods, reduction in leaf size, and stunting of the plant covering 30% to 50% of the foliage. | |
| 8 | Severe yellow discoloration of entirety of leaves covering above 50% to 75% of foliage, stunting of plants, and reduction in pod formation. | Highly susceptible (HS) |
| 9 | Severe yellowing of leaves covering above foliage, stunting of plants, and no pod formation. |
| S. No. | Genotype | Kharif 2018 | Rabi 2018 | ||||
|---|---|---|---|---|---|---|---|
| PDI | Grade | Range of PYL * | PDI | Grade | Range of PYL | ||
| Highly resistant (HR) | |||||||
| 1 | PLS 6002 | 1.2 | 2 | -- | 1.2 | 2 | 1.2–2.0 |
| Moderately resistant (MR) | |||||||
| 1 | PLS6039 | 12.5 | 4 | -- | 12.5 | 4 | 7.60–9.40 |
| 2 | PLS6008 | 12.5 | 4 | -- | 12.5 | 4 | 8.30–10.2 |
| 3 | PLS6013 | 12.5 | 4 | -- | 12.5 | 4 | 10.2–11.6 |
| Moderately susceptible (MS) | |||||||
| 1 | PLS 6004 | 20.0 | 5 | -- | 20.5 | 5 | 23.1–32.5 |
| 2 | PLS 6183 | 19.0 | 5 | -- | 19.5 | 5 | 32.3–35.6 |
| 3 | PLS 6006 | 19.0 | 5 | -- | 19.0 | 5 | 34.2–38.9 |
| Susceptible (S) | |||||||
| 1 | 2448984 | 49.0 | 7 | -- | 49.5 | 7 | 42.1–46.5 |
| 2 | PLS 6081 | 43.0 | 7 | -- | 44.0 | 7 | 49.8–46.7 |
| 3 | PLS 6046 | 42.0 | 7 | -- | 43.0 | 7 | 53.5–61.8 |
| Highly susceptible (HS) | |||||||
| 1 | PLS 6194 | 72.0 | 9 | -- | 73.0 | 9 | 75.0–89.4 |
| Susceptible check (SC) | |||||||
| 1 | HG 22: SC | 49.0 | 7 | -- | 49.0 | 7 | 46.9–62.7 |
| Moderately resistant checks (MRC) | |||||||
| 1 | CRIDA 1-18R | 12.50 | 4 | -- | 12.50 | 4 | 12.8–12.4 |
| 2 | PAIYUR 2 | 12.25 | 4 | -- | 12.25 | 4 | 10.1–10.9 |
| S. No | Class | Peak Area (%) | |||
|---|---|---|---|---|---|
| SC | HS | RC | HR | ||
| 1 | Alcohol | 0.142 | 1.156 | 0.286 | 0.661 |
| 2 | Aldehyde | 0.002 | 0.175 | 0.004 | 1.993 |
| 3 | Alkane | 0.212 | 8.035 | 3.145 | 15.30 |
| 4 | Alkene | - | 0.164 | - | 0.667 |
| 5 | Amide | - | 0.144 | - | - |
| 6 | Amine | - | - | - | 0.161 |
| 7 | Amino acids | - | - | - | 0.151 |
| 8 | Aromatic oxazole | - | - | - | 0.163 |
| 9 | Azide | - | - | - | 0.146 |
| 10 | Benzene | 0.113 | 0.354 | 0.143 | 0.561 |
| 11 | Carboxylic acid | 0.684 | 2.421 | 0.268 | 3.658 |
| 12 | Cyclic azines | - | 0.119 | - | - |
| 13 | Cyclic carboxylic ester | - | 0.766 | - | - |
| 14 | Ester | - | 0.204 | - | - |
| 15 | Ether | - | 0.119 | - | - |
| 16 | Glucosinolate | - | - | - | 0.412 |
| 17 | Glycoside | 4.289 | 17.914 | - | 6.277 |
| 18 | Hetero cyclic dioxin | - | 0.209 | - | - |
| 19 | Heterocyclic organo oxygen | - | - | - | 0.406 |
| 20 | Isoquinoline | - | - | - | 0.203 |
| 21 | Ketone | - | 2.381 | 0.152 | 1.991 |
| 22 | Lanostane skeleton | - | - | - | 0.158 |
| 23 | Sugars | 21.124 | 53.648 | 22.684 | 59.226 |
| 24 | Organo sulphur compound | - | 0.393 | - | 0.274 |
| 25 | Phosphonic acid | - | 0.248 | - | - |
| 26 | Triene | - | 0.304 | - | - |
| S. No. | Class | Biomolecule | HS | HR | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SI Value | RSI Value | RT | % Area | SI Value | RSI Value | RT | % Area | |||
| 1 | Alcohol | 2-Ethyl-1-hexanol | 813 * | 899 * | 4.664 | 0.125 | 786 | 886 * | 4.664 | 0.227 |
| 2,4,7,9-Tetramethyl-5-decyn-4,7-diol | 747 | 818 * | 10.106 | 0.151 | - | - | - | - | ||
| Galactitol | 407 | 585 | 12.522 | 0.562 | - | - | - | - | ||
| Phenol | 645 | 727 ** | - | - | - | - | ||||
| 9,10-Secocholesta-5,7,10(19)-triene-3,24,25-triol | - | - | - | - | 439 | 485 | 3.599 | 0.163 | ||
| 1,2-Ethanediol | - | - | - | - | 557 | 834 * | ||||
| 2 | Aldehyde | Nonanal | 801 * | 857 * | 5.814 | 0.171 | 806 * | 849 * | 5.810 | 0.169 |
| 2,2-Dimethyl-5-[2-(2-trimethylsilylethoxymethoxy)-propyl]-[1,3]dioxolane-4-carboxaldehyde | - | - | - | - | 632 | 675 | 23.492 | 1.824 | ||
| 3 | Alkane | 4-Methyl(trimethylene)silyloxyoctane | 621 | 640 | 25.942 | 0.173 | - | - | - | - |
| 1-Methyl-1-(3-phenylprop-2-enyl)oxy-1-silacyclobutane | 431 | 539 | 12.692 | 0.299 | - | - | - | - | ||
| 3,7,11,14,18-Pentaoxa-2,19-disilaeicosane | 651 | 653 | 13.718 | 2.182 | 643 | 649 | 14.368 | 0.594 | ||
| 1-Methyl-1-n-octyloxy-1-silacyclobutane | 536 | 649 | 24.652 | 0.110 | 620 | 660 | 24.417 | 0.214 | ||
| 3,7,11,15,18-Pentaoxa-2,19-disilaeicosane | 641 | 667 | - | - | - | - | ||||
| (2S,2′S)-2,2′-Bis[1,4,7,10,13-pentaoxacyclopentadecane] | 466 | 597 | 31.284 | 0.110 | - | - | - | - | ||
| 1-(Methoxymethoxy)-3-methyl-3-hydroxybutane | - | - | - | - | 569 | 699 | 4.264 | 0.407 | ||
| 17α,21β-28,30-Bisnorhopane | - | - | - | - | 479 | 526 | 30.710 | 0.120 | ||
| 4 | Alkene | 3(E)-Hydroxyimino-1,2,2,5,6,7-hexamethylbicyclo[3.2.0]hept-6-ene | 465 | 598 | 26.823 | 0.164 | - | - | - | - |
| 9-Octadecene | - | - | - | - | 464 | 493 | 3.694 | 0.134 | ||
| 3-Methoxyhex-1-ene | - | - | - | - | 560 | 761 ** | 4.469 | 0.136 | ||
| H-Indene | - | - | - | - | 513 | 539 | 23.897 | 0.397 | ||
| 5 | Amide | Deoxyspergualin | 559 | 592 | 4.264 | 0.144 | - | - | - | - |
| 6 | Amine | 2-Oxazolamine | - | - | - | - | 517 | 542 | 5.529 | 0.161 |
| 7 | Amino acids | Glycine | - | - | - | - | 406 | 453 | 11.627 | 0.151 |
| 8 | Aromatic oxazole compound | Benzoxazole | - | - | - | - | 603 | 846 * | 4.584 | 0.163 |
| 9 | Azide | Pyridine-4-carbohydrazide | - | - | - | - | 690 | 712 ** | 6.540 | 0.146 |
| 10 | Benzene | Benzene | 678 | 821 * | 7.490 | 0.354 | 684 | 805 * | 7.480 | 0.561 |
| 9-Octadecenoic acid | 495 | 519 | 26.042 | 0.611 | - | - | - | - | ||
| 5,8,11,14-Eicosatetraynoic acid | 449 | 466 | 13.933 | 0.148 | - | - | - | - | ||
| 1,2-Benzenedicarboxylic acid | 599 | 827 * | 19.195 | 0.108 | - | - | - | - | ||
| n-Hexadecanoic acid | 755 | 817 * | 21.191 | 0.272 | 857 * | 866 * | 21.186 | 1.342 | ||
| 9,12-Octadecadiynoic acid | 523 | 532 | 26.373 | 0.908 | - | - | - | - | ||
| Butanoic acid | 614 | 794 ** | 30.714 | 0.258 | - | - | - | - | ||
| Hexanoic acid | - | - | - | - | 578 | 747 ** | 3.829 | 0.263 | ||
| 6-Benzoylhexanoic acid | - | - | - | - | 544 | 647 | 5.274 | 0.169 | ||
| 11 | Carboxylic acid | cis-11-Eicosenoic acid | - | - | - | - | 487 | 548 | 9.486 | 0.142 |
| Tetradecanoic acid | - | - | - | - | 723 ** | 776 ** | 17.144 | 0.205 | ||
| Pentadecanoic acid | - | - | - | - | 625 | 688 | 19.195 | 0.175 | ||
| Heptadecanoic acid | - | - | - | - | 686 | 736 ** | 23.072 | 0.120 | ||
| Octadecanoic acid | - | - | - | - | 720 ** | 757 ** | 24.902 | 0.281 | ||
| 4,7,7-Trimethyl-3-oxo-2-oxabicyclo[2.2.1]heptane-1-carboxylic acid | - | - | - | - | 519 | 560 | 26.828 | 0.539 | ||
| Propanoic acid | - | - | - | - | 479 | 513 | 26.968 | 0.157 | ||
| Hexadecanoic acid | - | - | - | - | 492 | 507 | 27.238 | 0.123 | ||
| 10,12-Tricosadiynoic acid | - | - | - | - | 552 | 573 | 29.214 | 0.142 | ||
| 12 | Cyclic azines | 4H-Pyrazole | 411 | 549 | 10.791 | 0.119 | - | - | - | - |
| 13 | Cyclic carboxylic ester | 3-Deoxy-d-mannoic lactone | 622 | 749 ** | 14.423 | 0.766 | - | - | - | - |
| 14 | Ester | Methyl 2,2-dimethyl-3,6,9,12,15,18,21-heptaoxa-2-silatricosan-23-oate | 492 | 547 | 12.342 | 0.204 | - | - | - | - |
| 15 | Ether | Ether | 518 | 729 ** | 29.229 | 0.119 | - | - | - | - |
| 16 | Glucosinolate | Glucobrassicin | - | - | - | - | 490 | 498 | 24.472 | 0.412 |
| 17 | Glycoside | Galactopyranoside | 442 | 510 | 23.686 | 0.394 | - | - | - | - |
| α-D-Glucopyranoside | 879 * | 882 * | 13.462 | 17.520 | - | - | - | - | ||
| Methyl-α-D-thiogalactoside | - | - | - | - | 801 * | 875 * | 13.303 | 4.565 | ||
| Methyl 3-O-acetyl-2,4,6-tri-O-ethyl-α-d-mannopyranoside | - | - | - | - | 628 | 665 | 13.698 | 0.904 | ||
| Octyl-α-D-glucopyranoside | - | - | - | - | 464 | 539 | 23.707 | 0.579 | ||
| α-D-Galactopyranoside | - | - | - | - | 552 | 572 | 24.652 | 0.229 | ||
| 18 | Hetero cyclic dioxin | 4H-1,3-Benzodioxin | 708 ** | 741 ** | 6.540 | 0.209 | - | - | - | - |
| 19 | Heterocyclic organo oxygen | 2-Vinylfuran | - | - | - | - | 777 ** | 852 * | 3.919 | 0.406 |
| 20 | Isoquinoline | Papaveroline | - | - | - | - | 578 | 672 | 31.085 | 0.203 |
| 4′-Chlorobutyrophenone | 554 | 641 | 5.269 | 0.168 | - | - | - | - | ||
| 4H-Pyran-4-one | 714 | 882 * | 6.395 | 0.201 | - | - | - | - | ||
| 6-Amino-1-α-d-ribofuranosylimidazo[4,5-c]pyridin-4(5H)-one | 474 | 527 | 6.680 | 0.117 | - | - | - | - | ||
| 8,8-Dimethyl-3-(4-methyl-5-oxo-2,5-dihydrofuran-2-yloxymethylene)-3a,4,6,7,8,8b-hexahydro-3H-indeno[1,2-b]furan-2,5-dione | 485 | 516 | 7.535 | 0.421 | - | - | - | - | ||
| 5,6-Epoxy-6-methyl-2-heptanone | 529 | 662 | 10.001 | 0.164 | - | - | - | - | ||
| 21 | Ketone | Propan-1-one | ||||||||
| 2-Azetidinone | ||||||||||
| 4-Carbomethoxy-4-[2-(2-carbomethoxyvinyl)benzyl]-3-methoxy-2,5-cyclohexadien-1-one | ||||||||||
| Dihydroxyacetone | ||||||||||
| 2H-Pyran-2-one | ||||||||||
| 12-Oxatricyclo [4.4.3.0(1,6)] tridecane-3,11-dione | ||||||||||
| Androstan-3-one | ||||||||||
| 22 | Lanostane skeleton | Cucurbitacin b | - | - | - | - | 413 | 476 | 16.614 | 0.158 |
| 23 | Sugars | α-D-Glucopyranose | 753 | 882 * | 26.282 | 0.832 | 653 | 680 | 26.523 | 2.834 |
| Rhamnose | 695 | 759 ** | 3.824 | 0.197 | - | - | - | - | ||
| D-Allose | 722 | 826 * | 11.527 | 0.619 | ||||||
| 3-O-Methyl-d-glucose | 753 ** | 768 ** | 15.863 | 44.605 | 742 ** | 765 ** | 15.994 | 38.406 | ||
| 1,6-Anhydro-2,3-O-isopropylidene-α-D-mannopyranose | 639 | 679 | 23.491 | 0.920 | - | - | - | - | ||
| Melezitose | 729 ** | 731 ** | 10.521 | 1.813 | 658 | 673 | 35.883 | 14.636 | ||
| Maltose | 675 | 687 | 25.647 | 4.051 | - | - | - | - | ||
| α-D-Galactopyranose | 532 | 562 | 24.487 | 0.269 | 619 | 661 | 24.417 | 0.214 | ||
| 24 | Organo sulphur compound | Diphenyl sulfone | 827 * | 862 * | 20.285 | 0.393 | 799 | 831 * | 20.270 | 0.274 |
| 25 | Phosphonic acid | Phosphonic acid | 789 | 830 * | 3.914 | 0.248 | - | - | - | - |
| 26 | Triene | 4-Hexyl-1-(7-methoxycarbonylheptyl)bicyclo[4.4.0]deca-2,5,7-triene | 452 | 496 | 26.618 | 0.304 | - | - | - | - |
| S. No. | Class | Biomolecule | Chemical Formula | MW (g/mol) | HS | SC | HR | RC |
|---|---|---|---|---|---|---|---|---|
| 1 | Alcohol | 2-Ethyl-1-hexanol | C8H18O | 130.23 | √ | √ | √ | √ |
| 2,4,7,9-Tetramethyl-5-decyn-4,7-diol | C14H26O2 | 226.35 | √ | √ | - | - | ||
| Galactitol | C6H14O6 | 182.17 | √ | √ | - | - | ||
| Phenol | C6H6O | 94.11 | √ | √ | - | - | ||
| 9,10-Secocholesta-5,7,10(19)-triene-3,24,25-triol | C27H44O3 | 416.60 | - | - | √ | √ | ||
| 1,2-Ethanediol | (C2H4O)nH2O | 62.07 | √ | √ | √ | √ | ||
| 2 | Aldehyde | Nonanal | C9H18O | 142.24 | √ | - | √ | - |
| 2,2-Dimethyl-5-[2-(2-trimethylsilylethoxymethoxy)-propyl]-[1,3]dioxolane-4-carboxaldehyde | C15H30O5Si | 318.48 | √ | - | √ | - | ||
| 3 | Alkane | 4-Methyl(trimethylene)silyloxyoctane | C12H26OSi | 214.42 | √ | √ | - | - |
| 1-Methyl-1-(3-phenylprop-2-enyl)oxy-1-silacyclobutane | C13H18OSi | 218.36 | √ | √ | - | - | ||
| 3,7,11,14,18-Pentaoxa-2,19-disilaeicosane | C17H40O5Si2 | 380.70 | √ | √ | √ | √ | ||
| 1-Methyl-1-n-octyloxy-1-silacyclobutane | C12H26OSi | 214.41 | √ | √ | √ | √ | ||
| 3,7,11,15,18-Pentaoxa-2,19-disilaeicosane | C17H40O5Si2 | 380.7 | √ | √ | - | - | ||
| (2S,2′S)-2,2′-Bis[1,4,7,10,13-pentaoxacyclopentadecane] | C20H38O10 | 438.00 | √ | √ | - | - | ||
| 1-(Methoxymethoxy)-3-methyl-3-hydroxybutane | C7H16O3 | 148.20 | - | - | √ | √ | ||
| 17α,21β-28,30-Bisnorhopane | C29H50 | 398.71 | - | - | √ | √ | ||
| 4 | Alkene | 3(E)-Hydroxyimino-1,2,2,5,6,7-hexamethylbicyclo[3.2.0]hept-6-ene | C13H21ON | 207.00 | √ | - | - | - |
| 9-Octadecene | C18H36 | 252.50 | - | - | √ | - | ||
| 3-Methoxyhex-1-ene | C7H14O | 114.19 | - | - | √ | - | ||
| H-Indene | C9H8 | 116.16 | - | - | √ | - | ||
| 5 | Amide | Deoxyspergualin | C17H37N7O3 | 387.50 | √ | √ | - | |
| 6 | Amine | Z-Oxazolamine | C8H14N2O | 154.21 | - | - | √ | √ |
| 7 | Amino acids | Glycine | C2H5NO2 | 75.07 | - | - | √ | √ |
| 8 | Aromatic oxazole compound | Benzoxazole | C7H5NO | 119.12 | - | - | √ | √ |
| 9 | Azide | Pyridine-4-carbohydrazide | C12H14Cl3N6O2Ti | 428.50 | - | - | √ | √ |
| 10 | Benzene | Benzene | C6H6 | 78.11 | √ | √ | √ | √ |
| 11 | Carboxylic acid | 9-Octadecenoic acid | C18H34O2 | 282.50 | √ | √ | - | - |
| 5,8,11,14-Eicosatetraynoic acid | C20H24O2 | 296.40 | √ | √ | - | - | ||
| 1,2-Benzenedicarboxylic acid | C16H20O4 | 276.33 | √ | √ | - | - | ||
| n-Hexadecanoic acid | C16H32O2 | 256.42 | √ | √ | √ | √ | ||
| 9,12-Octadecadiynoic acid | C18H28O2 | 276.40 | √ | √ | - | - | ||
| Butanoic acid | C10H22O2Si | 202.37 | √ | √ | - | - | ||
| Hexanoic acid | C6H12O2 | 1.08 | - | - | √ | √ | ||
| 6-Benzoylhexanoic acid | C13H16O3 | 220.26 | - | - | √ | √ | ||
| cis-11-Eicosenoic acid | C20H38O2 | 310.50 | - | - | √ | √ | ||
| Tetradecanoic acid | C14H28O2 | 229.36 | - | - | √ | √ | ||
| Pentadecanoic acid | C15H30O2 | 242.40 | - | - | √ | √ | ||
| Heptadecanoic acid | C17H34O2 | 270.50 | - | - | √ | √ | ||
| Octadecanoic acid | C18H36O2 | 284.50 | - | - | √ | √ | ||
| 4,7,7-Trimethyl-3-oxo-2-oxabicyclo[2.2.1]heptane-1-carboxylic acid | C12H26OSi | 214.42 | - | - | √ | √ | ||
| Propanoic acid | C3H6O2 | 74.08 | - | - | √ | √ | ||
| Hexadecanoic acid | C16H32O2 | 256.42 | - | - | √ | √ | ||
| 10,12-Tricosadiynoic acid | C23H38O2 | 346.50 | - | - | √ | √ | ||
| 12 | Cyclic azines | 4H-Pyrazole | C3H4N2 | 68.08 | √ | - | - | - |
| 13 | Cyclic carboxylic ester | 3-Deoxy-d-mannoic lactone | C6H10O5 | 162.14 | √ | - | - | - |
| 14 | Ester | Methyl 2,2-dimethyl-3,6,9,12,15,18,21-heptaoxa-2-silatricosan-23-oate | C18H38O9Si | 426.60 | √ | - | - | - |
| 15 | Ether | Ether | C4H10O | 74.12 | √ | - | - | - |
| 16 | Glucosinolate | Glucobrassicin | C16H19N2O9S2- | 447.46 | - | - | - | √ |
| 17 | Glycoside | Galactopyranoside | C7H14O6 | 194.18 | √ | √ | - | - |
| α-D-Glucopyranoside | C7H14O6 | 194.18 | √ | √ | - | - | ||
| Methyl-α-D-thiogalactoside | C7H14O6 | 194.18 | - | - | √ | - | ||
| Methyl 3-O-acetyl-2,4,6-tri-O-ethyl-α-d-mannopyranoside | C13H22O8 | 306.31 | - | - | √ | - | ||
| Octyl-α-D-glucopyranoside | C14H28O6 | 292.37 | - | - | √ | - | ||
| α-D-Galactopyranoside | C12H15NO8 | 301.25 | - | - | √ | - | ||
| 18 | Hetero cyclic dioxin | 4H-1,3-Benzodioxin | C8H8O2 | 136.15 | √ | √ | - | - |
| 19 | Heterocyclic organo oxygen | 2-Vinylfuran | C6H6O | 94.11 | - | - | √ | √ |
| 20 | Isoquinoline | Papaveroline | C16H14BrNO4 | 364.19 | √ | √ | ||
| 21 | Ketone | 4′-Chlorobutyrophenone | C10H11ClO | 182.64 | √ | - | - | - |
| 4H-Pyran-4-one | C5H4O2 | 96.08 | √ | - | - | - | ||
| 6-Amino-1-α-d-ribofuranosylimidazo[4,5-c]pyridin-4(5H)-one | C19H14N2O2 | 302.3 | √ | - | - | - | ||
| 8,8-Dimethyl-3-(4-methyl-5-oxo-2,5-dihydrofuran-2-yloxymethylene)-3a,4,6,7,8,8b-hexahydro-3H-indeno[1,2-b]furan-2,5-dione | C39H40O12 | 700.73 | √ | - | - | - | ||
| 5,6-Epoxy-6-methyl-2-heptanone | C8H14O | 142.20 | √ | - | - | - | ||
| Propan-1-one | C12H15NO2 | 205.25 | √ | - | - | - | ||
| 2-Azetidinone | C3H5NO | 71.08 | √ | - | - | - | ||
| 4-Carbomethoxy-4-[2-(2-carbomethoxyvinyl)benzyl]-3-methoxy-2,5-cyclohexadien-1-one | C20H20O6 | 356.37 | √ | - | - | - | ||
| Dihydroxyacetone | C3H6O3 | 90.08 | - | - | √ | √ | ||
| 2H-Pyran-2-one | C5H4O2 | 96.08 | - | - | √ | √ | ||
| 12-Oxatricyclo [4.4.3.0(1,6)] tridecane-3,11-dione | C12H16O3 | 208.25 | - | - | √ | √ | ||
| Androstan-3-one | C19H30O | 274.40 | - | - | √ | √ | ||
| 22 | Lanostane skeleton | Cucurbitacin b | C64H90O16 | 1115.40 | - | - | √ | √ |
| 23 | Sugars | α-D-Glucopyranose | C6H12O | 180.16 | √ | √ | √ | √ |
| Rhamnose | C6H12O5 | 164.07 | √ | √ | - | - | ||
| D-Allose | C6H12O6 | 180.16 | - | - | √ | √ | ||
| 3-O-Methyl-d-glucose | C7H14O6 | 194.18 | √ | √ | √ | √ | ||
| 1,6-Anhydro-2,3-O-isopropylidene-á-D-mannopyranose | C9H14O5 | 202.20 | √ | √ | - | - | ||
| Melezitose | C12H22O11 | 342.30 | √ | √ | √ | √ | ||
| Maltose | C12H22O11 | 342.30 | √ | √ | - | - | ||
| α-D-Galactopyranose | C26H42O11 | 530.60 | √ | √ | √ | √ | ||
| 24 | Organo sulphur compound | Diphenyl sulfone | C12H10O2S | 218.27 | √ | √ | √ | √ |
| 25 | Phosphonic acid | Phosphonic acid | H3PO3 | 80.98 | √ | √ | - | - |
| 26 | Triene | 4-Hexyl-1-(7-methoxycarbonylheptyl)bicyclo[4.4.0]deca-2,5,7-triene | C25H40O2 | 372.60 | √ | √ | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajaprakasam, S.; Shanmugavel, P.; Chockalingam, V.; Jegadeesan, S.; Latha, T.K.S.; Ananthan, S.N.; Muthurajan, R.; Kanagarajan, S. Comparative Metabolomic Profiling of Horse Gram (Macrotyloma uniflorum (Lam.) Verdc.) Genotypes for Horse Gram Yellow Mosaic Virus Resistance. Metabolites 2023, 13, 165. https://doi.org/10.3390/metabo13020165
Rajaprakasam S, Shanmugavel P, Chockalingam V, Jegadeesan S, Latha TKS, Ananthan SN, Muthurajan R, Kanagarajan S. Comparative Metabolomic Profiling of Horse Gram (Macrotyloma uniflorum (Lam.) Verdc.) Genotypes for Horse Gram Yellow Mosaic Virus Resistance. Metabolites. 2023; 13(2):165. https://doi.org/10.3390/metabo13020165
Chicago/Turabian StyleRajaprakasam, Sudhagar, Priyanka Shanmugavel, Vanniarajan Chockalingam, Souframanien Jegadeesan, Tnpalayam Krishnaswamy Sukirtha Latha, Saravanan Naaganoor Ananthan, Raveendran Muthurajan, and Selvaraju Kanagarajan. 2023. "Comparative Metabolomic Profiling of Horse Gram (Macrotyloma uniflorum (Lam.) Verdc.) Genotypes for Horse Gram Yellow Mosaic Virus Resistance" Metabolites 13, no. 2: 165. https://doi.org/10.3390/metabo13020165
APA StyleRajaprakasam, S., Shanmugavel, P., Chockalingam, V., Jegadeesan, S., Latha, T. K. S., Ananthan, S. N., Muthurajan, R., & Kanagarajan, S. (2023). Comparative Metabolomic Profiling of Horse Gram (Macrotyloma uniflorum (Lam.) Verdc.) Genotypes for Horse Gram Yellow Mosaic Virus Resistance. Metabolites, 13(2), 165. https://doi.org/10.3390/metabo13020165