


Cytotoxic Isopentenyl Phloroglucinol Compounds from Garcinia xanthochymus Using LC-MS-Based Metabolomics


Abstract
:
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Plant Materials
2.3. Extraction and Isolation
2.4. Cell Viability Assay
2.5. UPLC-QTof-MS Analysis
2.6. Metabolomics Data Analysis
3. Results and Discussion
3.1. Cytotoxic Activity Screening of Different Part Extracts of G. xanthochymus
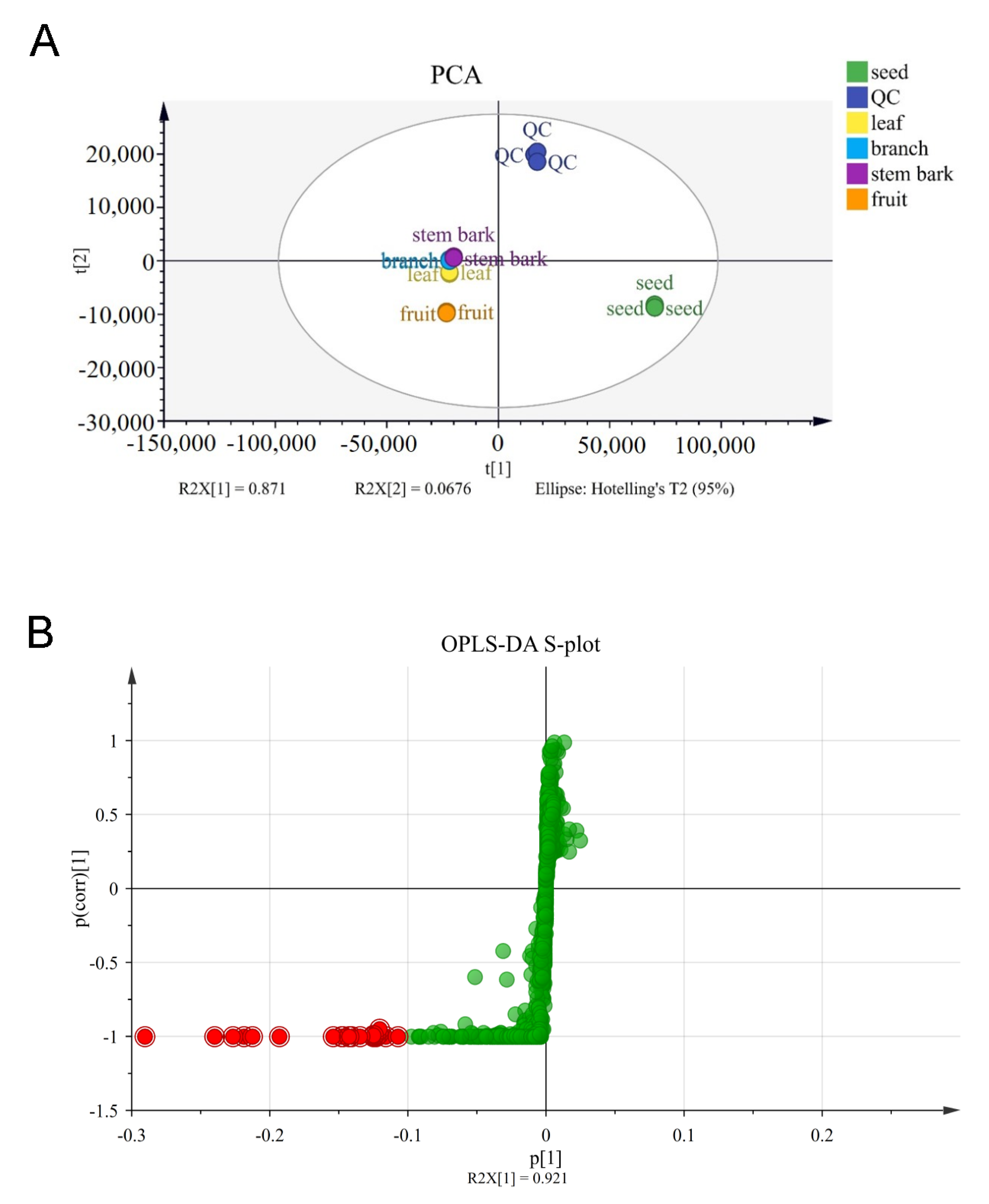
3.2. Metabolites in Different Parts of G. xanthochymus and Marker Compounds’ Identification
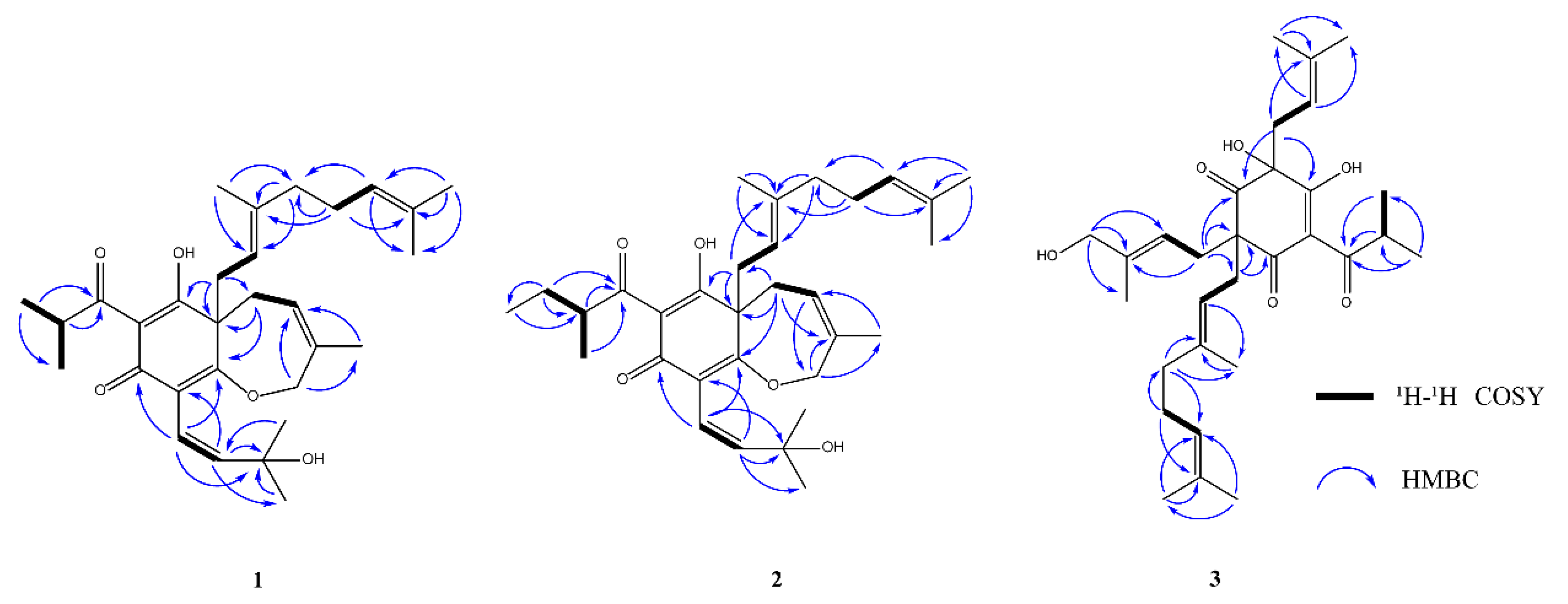
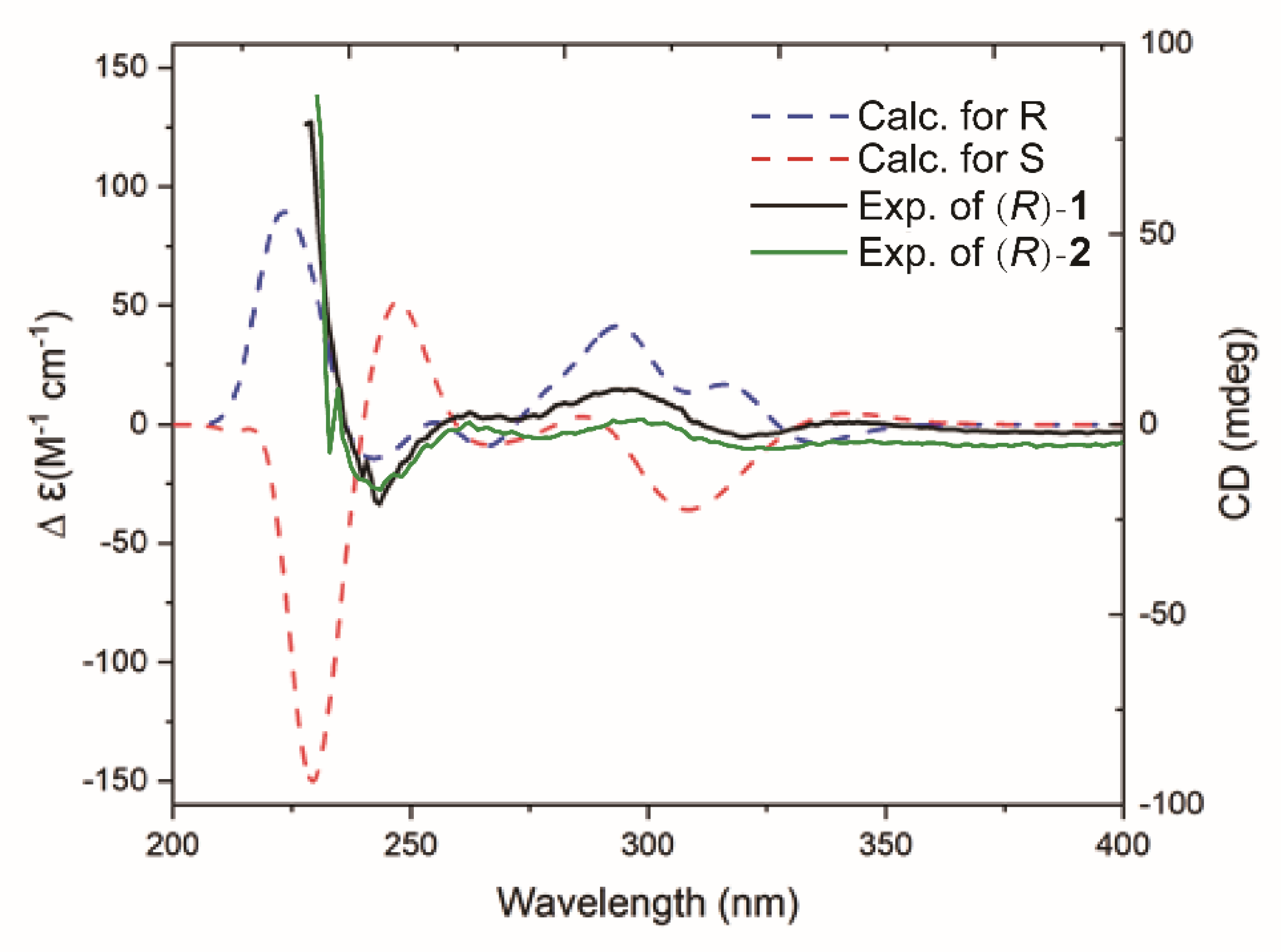
3.3. Isolation and Structure Elucidation of Metabolites from Target Markers
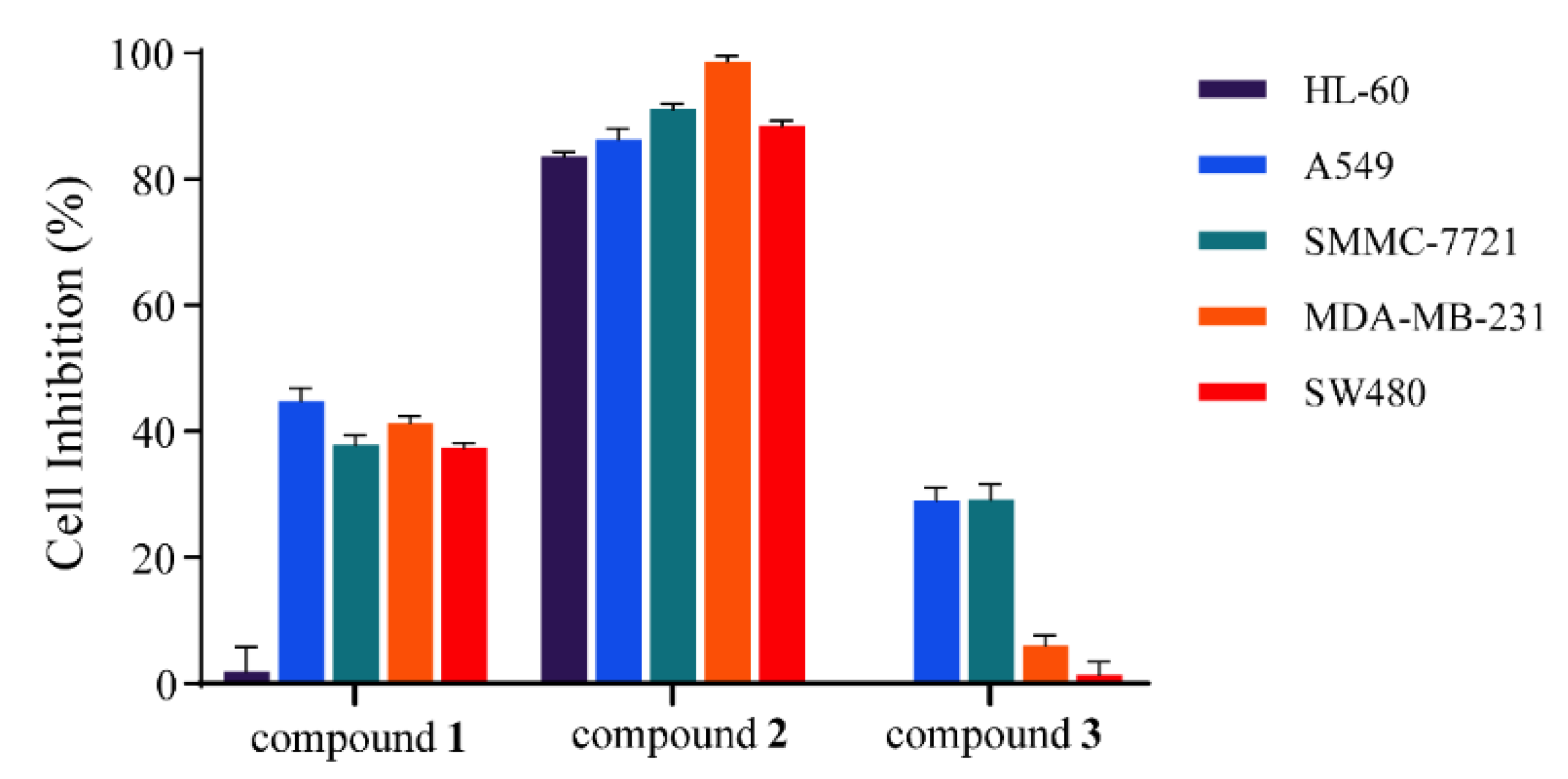
3.4. Cytotoxic Effect of Compounds Isolated from G. xanthochymus Seed
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adley, B. Editorial Note for Journal of Tumor Research and Reports. J. Tumour. Res. Rep. 2021, 6, 1–2. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA-Cancer. J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Xu, J.; Liu, X.; Zhen, Y. Advances and Challenges in the Treatment of Esophageal Cancer. Inst. Med. Biotechnol. Chin. Acad. Med. Sci. Peking Union Med. Coll. 2021, 11, 3379–3392. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Ozleyen, A.; Tumer, T.B.; Adetunji, C.O.; Omari, N.E.; Balahbib, A.; Taheri, Y.; Bouyahya, A.; Martorell, M.; Martins, N.; et al. Natural Products and Synthetic Analogs as a Source of Antitumor Drugs. Biomolecules 2019, 9, 679. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products As Sources of New Drugs Over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Plazas, E.; Casoti, R.; Avila Murillo, M.; Batista Da Costa, F.; Cuca, L.E. Metabolomic Profiling of Zanthoxylum Species: Identification of Anti-Cholinesterase Alkaloids Candidates. Phytochemistry 2019, 168, 112128. [Google Scholar] [CrossRef]
- Olivon, F.; Allard, P.M.; Koval, A.; Righi, D.; Genta-Jouve, G.; Neyts, J.; Apel, C.; Pannecouque, C.; Nothias, L.-F.; Cachet, X.; et al. Bioactive Natural Products Prioritization Using Massive Multi-Informational Molecular Networks. ACS Chem. Biol. 2017, 12, 2644–2651. [Google Scholar] [CrossRef]
- Cox, D.G.; Oh, J.; Keasling, A.; Colson, K.L.; Hamann, M.T. The Utility of Metabolomics in Natural Product and Biomarker Characterization. Biochim. Biophys. Acta 2014, 1840, 3460–3474. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Xiao, C.; Chen, D.; Xiao, Y.; Mei, Z. Rapid Screening and Identification of α-Amylase Inhibitors from Garcinia xanthochymus Using Enzyme-Immobilized Magnetic Nanoparticles Coupled with HPLC and MS. J. Chromatogr. B 2014, 960, 166–173. [Google Scholar] [CrossRef]
- Kulakowski, D.M.; Wu, S.-B.; Balick, M.J.; Kennelly, E.J. Merging Bioactivity with Liquid Chromatography-Mass Spectrometry-Based Chemometrics to Identify Minor Immunomodulatory Compounds from a Micronesian Adaptogen, Phaleria nisidai. J. Chromatogr. A 2014, 1364, 74–82. [Google Scholar] [CrossRef]
- Viswajyothi, K. First Report of Coconut Coreid Bug Paradasynus Rostratus (Distant) on Garcinia xanthochymus. Indian J. Entomol. 2019, 81, 567–569. [Google Scholar] [CrossRef]
- Han, Q.B.; Qiao, C.F.; Song, J.Z.; Yang, N.Y.; Cao, X.W.; Peng, Y.; Yang, D.J.; Chen, S.L.; Xu, H.X. Cytotoxic Prenylated Phenolic Compounds from the Twig Bark of Garcinia xanthochymus. Chem. Biodivers. 2007, 4, 940–946. [Google Scholar] [CrossRef]
- Chen, Y.; Gan, F.; Jin, S.; Liu, H.; Wu, S.J.; Yang, W.T.; Yang, G.Z. Adamantyl Derivatives and Rearranged Benzophenones from Garcinia xanthochymus Fruits. RSC Adv. 2017, 7, 17289–17296. [Google Scholar] [CrossRef]
- Chen, Y.; Fan, H.; Yang, G.Z.; Jiang, Y.; Zhong, F.F.; He, H.W. Two Unusual Xanthones from the Bark of Garcinia xanthochymus. Helv. Chim. Acta 2011, 94, 662–668. [Google Scholar] [CrossRef]
- Hassan, N.K.N.C.; Taher, M.; Susanti, D. Phytochemical Constituents and Pharmacological Properties of Garcinia xanthochymus—A Review. Biomed. Pharmacother. 2018, 106, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.B.; Handique, P.J.; Devi, H.S. Antioxidant Properties, Physico-Chemical Characteristics and Proximate Composition of Five Wild Fruits of Manipur, India. J. Food Sci. Technol. 2015, 52, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Mahfuzur, R.M.; Kawsar, H.; Rokeya, K.M.; Keitaro, I.; Arihiro, I.; Kiyotake, S.; Hisashi, K. Identification and Application of Bioactive Compounds from Garcinia xanthochymus Hook. for Weed Management. Appl. Sci. 2021, 11, 2264. [Google Scholar]
- Joseph, K.S.; Dandin, V.S.; Hosakatte, N.M. Chemistry and Biological Activity of Garcinia xanthochymus: A Review. J. Biol. Act. Prod. Nat. 2016, 6, 173–194. [Google Scholar]
- Li, P.; Yue, G.G.L.; Kwok, H.F.; Long, C.L.; Lau, C.B.S.; Kennelly, E.J. Using Ultra-Performance Liquid Chromatography Quadrupole Time of Flight Mass Spectrometry-Based Chemometrics for the Identification of Anti-angiogenic Biflavonoids from Edible Garcinia Species. J. Agric. Food Chem. 2017, 65, 8348–8355. [Google Scholar] [CrossRef]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an Aqueous Soluble Tetrazolium/Formazan Assay for Cell Growth Assays in Culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vandergheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Yao, X.Y.; Hou, H.Y.; Liang, H.; Chen, K.; Chen, X.B. Raman Spectroscopy Study of Phosphorites Combined with PCA-HCA and OPLS-DA Models. Minerals 2019, 9, 578. [Google Scholar] [CrossRef]
- Surowiec, I.; Fraser, P.D.; Patel, R.; Halket, J.; Bramley, P.M. Metabolomic Approach for the Detection of Mechanically Recovered Meat in Food Products. Food Chem. 2010, 125, 1468–1475. [Google Scholar] [CrossRef]
- Klaiklay, S.; Sukpondma, Y.; Rukachaisirikul, V.; Phongpaichit, S. Friedolanostanes and Xanthones from the Twigs of Garcinia hombroniana. Phytochemistry 2013, 85, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Pailee, P.A.; Kruahong, T.B.; Hongthong, S.A.; Kuhakarn, C.A.; Jaipetch, T.A.; Pohmakotr, M.A.; Jariyawat, S.C.; Suksen, K.C.; Akkarawongsapat, R.D.; Limthongkul, J.D.; et al. Cytotoxic, Anti-HIV–1 and Anti-Inflammatory Activities of Lanostanes from Fruits of Garcinia speciosa. Phytochem. Lett. 2017, 20, 111–118. [Google Scholar] [CrossRef]
- Weng, J.R.; Tsao, L.T.; Wang, J.P.; Wu, R.R.; Lin, C.N. Anti-Inflammatory Phloroglucinols and Terpenoids from Garcinia subelliptica. J. Nat. Prod. 2004, 67, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.W.; Huang, A.M.; Yang, S.C.; Weng, J.R.; Hour, T.C.; Pu, Y.S.; Lin, C.N. Cytotoxic and Antioxidant Constituents from Garcinia subelliptica. Food Chem. 2012, 135, 851–859. [Google Scholar] [CrossRef]
- Yun, Y.S.; Shioura, M.; Hitotsuyanagi, Y.; Yotsumoto, S.; Inoue, H. Garcinielliptone G from Garcinia subelliptica Induces Apoptosis in Acute Leukemia Cells. Molecules 2021, 26, 2422. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Minami, H.; Kuwayama, A. Garsubellins, Polyisoprenylated Phloroglucinol Derivatives from Garcinia subelliptica. Phytochemistry 1998, 49, 853–857. [Google Scholar] [CrossRef]
- Liaw, C.C.; Chang, J.L.; Wang, B.W.; Chen, P.L.; Weng, J.R.; Lin, K.W.; Lin, C.N. Discovering a Racemate Polycyclic Prenylated Acylphloroglucinol with Unprecedented Skeleton by an ESI-LCMS Analytical Approach. Org. Lett. 2019, 21, 857–861. [Google Scholar] [CrossRef]
- Forino, M.; Pace, S.; Chianese, G.; Santagostini, L.; Taglialatela-Scafati, O. Humudifucol and Bioactive Prenylated Polyphenols from Hops (Humulus lupulus cv. “Cascade”). J. Nat. Prod. 2016, 79, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Baggett, S.; Protiva, P.; Mazzola, E.P.; Yang, H.; Ressler, E.T.; Basile, M.J.; Weinstein, I.B.; Kennelly, E.J. Bioactive Benzophenones from Garcinia xanthochymus Fruits. J. Nat. Prod. 2005, 68, 354–360. [Google Scholar] [CrossRef]
- Protiva, P.; Hopkins, M.E.; Baggett, S.; Yang, H.; Lipkin, M.; Holt, P.R.; Kennelly, E.J.; Bernard, W.I. Growth Inhibition of Colon Cancer Cells by Polyisoprenylated Benzophenones is Associated with Induction of the Endoplasmic Reticulum Response. Int. J. Cancer 2008, 123, 687–694. [Google Scholar] [CrossRef]
- Xu, J.; Jin, S.; Gan, F.; Xiong, H.; Mei, Z.; Chen, Y.; Yang, G. Polycyclic Polyprenylated Acylphloroglucinols from Garcinia xanthochymus Fruits Exhibit Antitumor Effects through Inhibition of the STAT3 Signaling Pathway. Food Funct. 2020, 11, 10568–10579. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.H.; Grossman, R.B.; Qiu, Y.F.; Luo, Y.; Lan, T.; Yang, X.W. Polycyclic Polyprenylated Acylphloroglucinols Bearing a Lavandulyl- Derived Substituent from Garcinia xanthochymus Fruits. J. Nat. Prod. 2022, 85, 2845–2855. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.G.; Lin, T.S.; Yu, H.N.; Chen, Y.; Zhan, Z.J. Polyprenylated Xanthones and Benzophenones from the Bark of Garcinia oblongifolia. Helv. Chim. Acta 2012, 95, 1442–1448. [Google Scholar] [CrossRef]
- Wu, S.B.; Long, C.L.; Kennell, E.J. Structural Diversity and Bioactivities of Natural Benzophenones. Nat. Prod. Rep. 2014, 31, 1158–1174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Chiou, C.T.; Cheng, J.J.; Huang, H.C.; Kuo, L.M.Y.; Liao, C.C.; Bastow, K.F.; Lee, K.H.; Kuo, Y.H. Cytotoxic Polyisoprenyl Benzophenonoids from Garcinia subelliptica. J. Nat. Prod. 2010, 73, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Charette, N.; De Saeger, C.; Lannoy, V.; Horsmans, Y.; Leclercq, I.; Starkel, P. Salirasib Inhibits the Growth of Hepatocarcinoma Cell Lines in Vitro and Tumor Growth in Vivo Through Ras and mTOR Inhibition. Mol. Cancer 2010, 9, 256. [Google Scholar] [CrossRef]
- Qiu, G.H.; Xie, X.J.; Xu, F.; Shi, X.H.; Wang, Y.; Deng, L.H. Distinctive Pharmacological Differences between Liver Cancer Cell Lines HepG2 and Hep3B. Cytotechnology 2015, 67, 1–12. [Google Scholar] [CrossRef]
- Hu, Z.X.; Xu, H.C.; Hu, K.; Liu, M.; Li, X.N.; Li, X.R.; Du, X.; Zhang, Y.H.; Puno, P.T.; Sun, H.D. Structurally Diverse Diterpenoids from Isodon pharicus. Org. Chem. Front. 2018, 5, 2379–2389. [Google Scholar] [CrossRef]
- Manohar, S.H.; Naik, P.M.; Patil, L.M.; Karikatti, S.I.; Murthy, H.N. Chemical Composition of Garcinia xanthochymus Seeds, Seed Oil, and Evaluation of Its Antimicrobial and Antioxidant Activity. J. Herbs Spices Med. Plants 2014, 20, 148–155. [Google Scholar] [CrossRef]
- Ruma, K.; Sunil, K.; Prakash, H.S. Antioxidant, Anti-inflammatory, Antimicrobial and Cytotoxic Properties of Fungal Endophytes from Garcinia Species. Int. J. Pharm. Pharm. Sci. 2013, 5, 889–897. [Google Scholar]
- Payamalle, S.; Joseph, K.S.; Bijjaragi, S.C.; Aware, C.; Jadhav, J.P.; Murthy, H.N.J.C.C.P. Anti-Diabetic Activity of Garcinia xanthochymus Seeds. Comp. Clin. Path. 2017, 26, 437–446. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Human Cancer Cell Lines | ||||
|---|---|---|---|---|---|
| HL-60 | A459 | SMMC-7721 | MCF-7 | SW480 | |
| Seed extract | 8.637 ± 0.124 | 3.436 ± 0.169 | 2.666 ± 0.130 | 8.362 ± 0.562 | 9.438 ± 0.290 |
| DDP b | 1.312 ± 0.024 | 12.20 ± 0.22 | 2.527 ± 0.145 | 18.75 ± 0.55 | 15.50 ± 0.99 |
| Taxol b | <0.008 | <0.008 | 0.138 ± 0.008 | <0.008 | <0.008 |
| Marker | RT (min) | Molecular Ions [M+H]+/[M-H]− | Molecular Formula | Identification | ||
|---|---|---|---|---|---|---|
| m/z | M.F. | ppm | ||||
| s1 | 5.613 | 517.3531 | C31H49O6 | 0.4 | C31H48O6 | garcihombronane L [24] |
| 515.3381 | C31H47O6 | 1.6 | ||||
| s2 | 6.595 | 487.3430 | C30H47O5 | 1.4 | C30H46O5 | garciosaterpene E or its isomers [25] |
| 485.3279 | C30H45O5 | 2.5 | ||||
| s3 | 6.692 | 515.3376 | C31H47O6 | 0.6 | C31H46O6 | unknown |
| 513.3221 | C31H45O6 | 1.0 | ||||
| s4 | 6.944 | 485.3291 | C30H45O5 | 4.9 | C30H44O5 | garcinielliptone L or garsubelline A [26,27] |
| 483.3117 | C30H43O5 | 1.4 | ||||
| s5 | 6.996 | 485.3287 | C30H45O5 | 4.1 | C30H45O5 | garcinielliptone F [28] |
| 483.3126 | C30H43O5 | 3.3 | ||||
| s6 | 7.208 | 501.3616 | C31H49O5 | 7.2 | C31H48O5 | unknown |
| 499.3446 | C31H47O5 | 4.6 | ||||
| s7 | 7.495 | 499.3446 | C31H47O5 | 4.6 | C31H46O5 | garsubellin B [29] |
| 497.3280 | C31H45O5 | 2.6 | ||||
| s8 | 7.661 | 499.3456 | C31H47O5 | 6.6 | C31H46O5 | garcinielliptone T [24] |
| 497.3287 | C31H45O5 | 4.0 | ||||
| s9 | 8.310 | 485.3267 | C30H45O5 | 0.0 | C30H44O5 | garcinielliptone L or garsubelline A [30] |
| 483.3115 | C30H43O5 | 1.0 | ||||
| s10 | 8.907 | 499.3428 | C31H47O5 | 1.0 | C31H46O5 | garsubellin E [29] |
| 497.3276 | C31H45O5 | 1.8 | ||||
| s11 | 8.023 | 483.3117 | C30H43O5 | 1.4 | C30H42O5 | unknown |
| 481.2955 | C30H41O5 | 0.2 | ||||
| s12 | 8.660 | 497.3270 | C31H45O5 | 0.6 | C31H44O5 | unknown |
| 495.3113 | C31H43O5 | 0.6 | ||||
| Position | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|
| δH (J in Hz) | δC | δH (J in Hz) | δC | δH (J in Hz) | δC | |
| 1 | 186.9 | 187.0 | 205.7 | |||
| 2 | 106.8 | 106.9 | 62.8 | |||
| 3 | 171.4 | 171.4 | 198.0 | |||
| 4 | 57.8 | 57.8 | 111.2 | |||
| 5 | 196.4 | 196.5 | 192.7 | |||
| 6 | 107.3 | 107.9 | 85.1 | |||
| 7 | 2.50, d (7.1) 2.92, dd (13.5, 10.4) | 35.7 | 2.50, d (6.8) 2.92, ddd (13.5, 10.4, 7.9) | 35.9 | 2.76, dd (13.8, 6.3) 2.86, m | 33.9 |
| 8 | 4.90, dd (10.7, 6.0) | 122.1 | 4.90, dd (9.8, 4.2) | 122.0 | 5.05, t (7.4) | 118.4 |
| 9 | 138.5 | 138.6 | 140.6 | |||
| 10 | 3.78, d (12.0) | 61.8 | 3.77, m | 61.8 | 1.90, m | 40.2 |
| 4.29, d (12.0) | 4.30, dd (14.7, 12.2) | |||||
| 11 | 1.66, s | 22.3 | 1.65, d (1.2) | 22.3 | 1.97, dd (14.9, 7.0) | 26.6 |
| 12 | 2.61, dd (13.7, 7.8) 2.48, d (7.1) | 39.1 | 2.61, dd (13.7, 7.7) 2.48, d (7.2) | 39.0 | 4.97, m | 124.0 |
| 13 | 4.81, t (7.2) | 117.4 | 4.80, dd (8.1, 7.0) | 117.4 | 132.0 | |
| 14 | 139.5 | 139.4 | 1.62, s | 26.1 | ||
| 15 | 1.84, d (6.6) | 40.0 | 1.83, m | 40.0 | 1.52, s | 17.8 |
| 16 | 1.87, m | 26.9 | 1.86, m | 27.1 | 1.63, s | 16.5 |
| 17 | 4.96, dd (6.7, 5.4) | 124.1 | 4.95, m | 124.1 | 2.89, m 2.27, dd (13.0, 6.3) | 40.0 |
| 18 | 131.8 | 131.8 | 4.75, m | 119.7 | ||
| 19 | 1.61, s | 25.9 | 1.61, d (0.7) | 25.9 | 141.6 | |
| 20 | 1.09, d (6.8) | 17.8 | 1.06, d (6.8) | 17.8 | 1.59, s | 21.6 |
| 21 | 1.51, t (3.5) | 16.6 | 1.51, d, (1.9) | 16.6 | 4.07, d (12.2) 3.60, d (12.2) | 60.9 |
| 22 | 6.45, d (10.0) | 114.8 | 6.45, d (10.1) | 114.8 | 2.56, dd, (14.7, 9.2) 2.05, dd (13.7, 6.4) | 37.2 |
| 23 | 5.36, d (10.1) | 123.8 | 5.35, d (10.1) | 123.8 | 4.85, dd (8.0, 6.6) | 115.0 |
| 24 | 81.5 | 81.5 | 137.5 | |||
| 25 | 1.46, s | 28.9 | 1.46, s | 28.9 | 1.43, s | 18.2 |
| 26 | 1.45, s | 29.0 | 1.45, s | 29.0 | 1.63, s | 25.8 |
| 27 | 208.1 | 207.6 | 207.1 | |||
| 28 | 3.92, m | 35.8 | 3.81, dd (13.7, 6.9) | 42.1 | 3.44, m | 35.3 |
| 29 | 1.07, d (6.8) | 19.3 | 1.10, m | 16.4 | 1.27, d (6.8) | 20.5 |
| 30 | 1.12, d (6.8) | 18.8 | 1.33, m | 26.9 | 1.12, d (6.7) | 18.0 |
| 31 | 0.92, t (7.4) | 12.0 | ||||
| Compound | Cancer Cell Lines | ||||
|---|---|---|---|---|---|
| HL-60 | A459 | SMMC-7721 | MDA-MB-231 | SW480 | |
| 2 | 19.02 ± 1.26 | 24.43 ± 0.42 | 15.69 ± 0.57 | 16.29 ± 1.44 | 14.71 ± 0.33 |
| DDP b | 16.07 ± 0.32 | 28.06 ± 2.29 | 24.44 ± 0.75 | 26.69 ± 0.50 | 28.83 ± 0.83 |
| Taxol b | <0.008 | <0.008 | 0.312 ± 0.019 | <0.008 | <0.008 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, F.; Luan, X.; Zhang, J.; Gao, W.; Yan, J.; Li, P. Cytotoxic Isopentenyl Phloroglucinol Compounds from Garcinia xanthochymus Using LC-MS-Based Metabolomics. Metabolites 2023, 13, 258. https://doi.org/10.3390/metabo13020258
Quan F, Luan X, Zhang J, Gao W, Yan J, Li P. Cytotoxic Isopentenyl Phloroglucinol Compounds from Garcinia xanthochymus Using LC-MS-Based Metabolomics. Metabolites. 2023; 13(2):258. https://doi.org/10.3390/metabo13020258
Chicago/Turabian StyleQuan, Fan, Xinbo Luan, Jian Zhang, Wenjie Gao, Jian Yan, and Ping Li. 2023. "Cytotoxic Isopentenyl Phloroglucinol Compounds from Garcinia xanthochymus Using LC-MS-Based Metabolomics" Metabolites 13, no. 2: 258. https://doi.org/10.3390/metabo13020258
APA StyleQuan, F., Luan, X., Zhang, J., Gao, W., Yan, J., & Li, P. (2023). Cytotoxic Isopentenyl Phloroglucinol Compounds from Garcinia xanthochymus Using LC-MS-Based Metabolomics. Metabolites, 13(2), 258. https://doi.org/10.3390/metabo13020258