


Enhanced Antioxidant and Anticancer Potential of Artemisia carvifolia Buch Transformed with rol A Gene


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Germination and DNA Barcoding
2.2. Bacterial Strains and Plasmids
2.3. Transformation and Regeneration
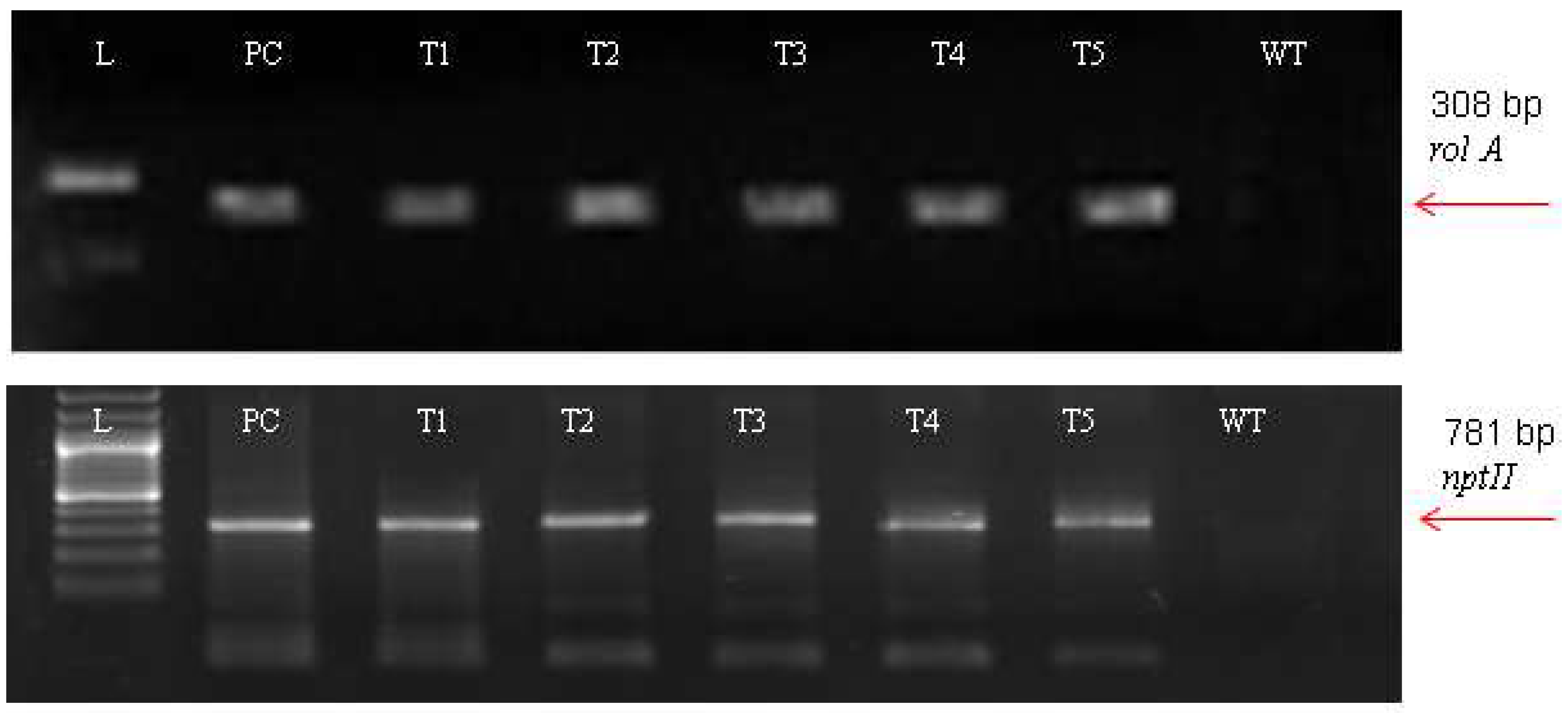
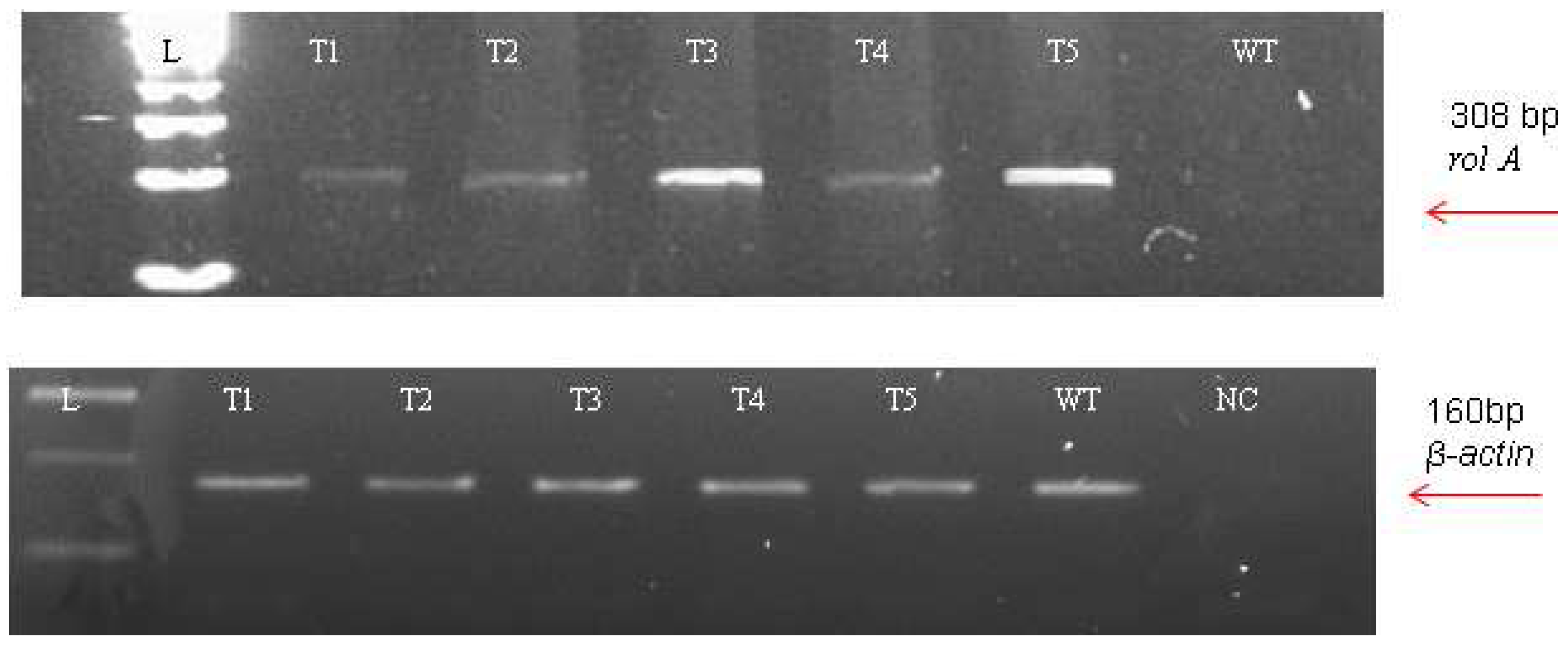
2.4. Molecular Analysis
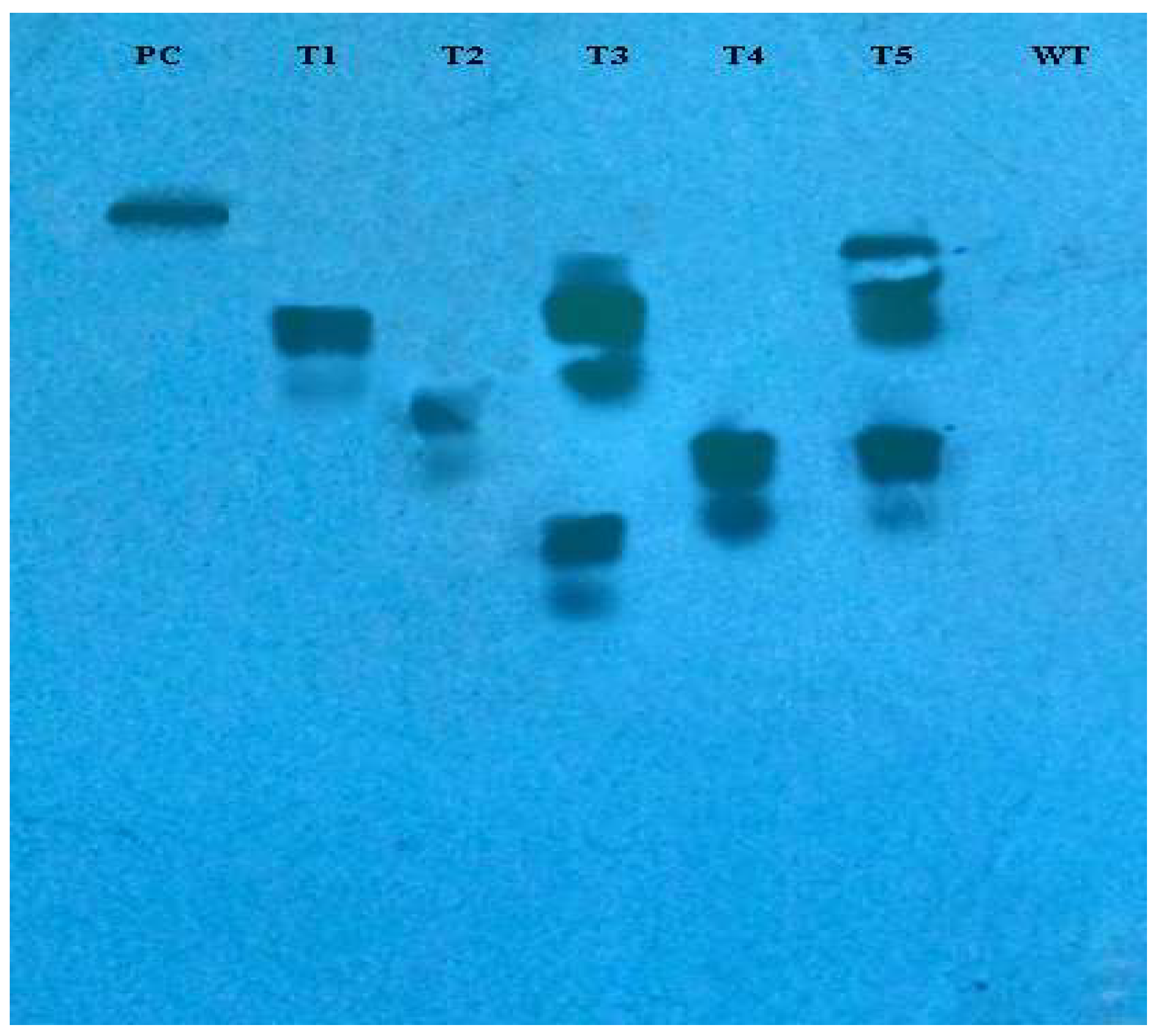
2.5. Southern Blotting
2.6. Evaluation of Flavonoids through an HPLC-DAD System
2.7. Analysis of Genes Involved in the Flavonoid Biosynthetic Pathway Using Real-Time qPCR
2.8. Antioxidant Potential Measurement
2.8.1. Total Phenolic Content Measurement
2.8.2. Total Flavonoid Content Measurement
2.8.3. Total Antioxidant Capacity Measurement
2.8.4. Total Reducing Power Measurement
2.9. DPPH Free Radical Scavenging Assay
2.10. Measurement of Anticancerous Activity through MTT Assay
2.11. Statistical Analysis
3. Results
3.1. DNA Barcoding for Identification of Plant
3.2. Transformation and Regeneration
3.3. Molecular Analysis
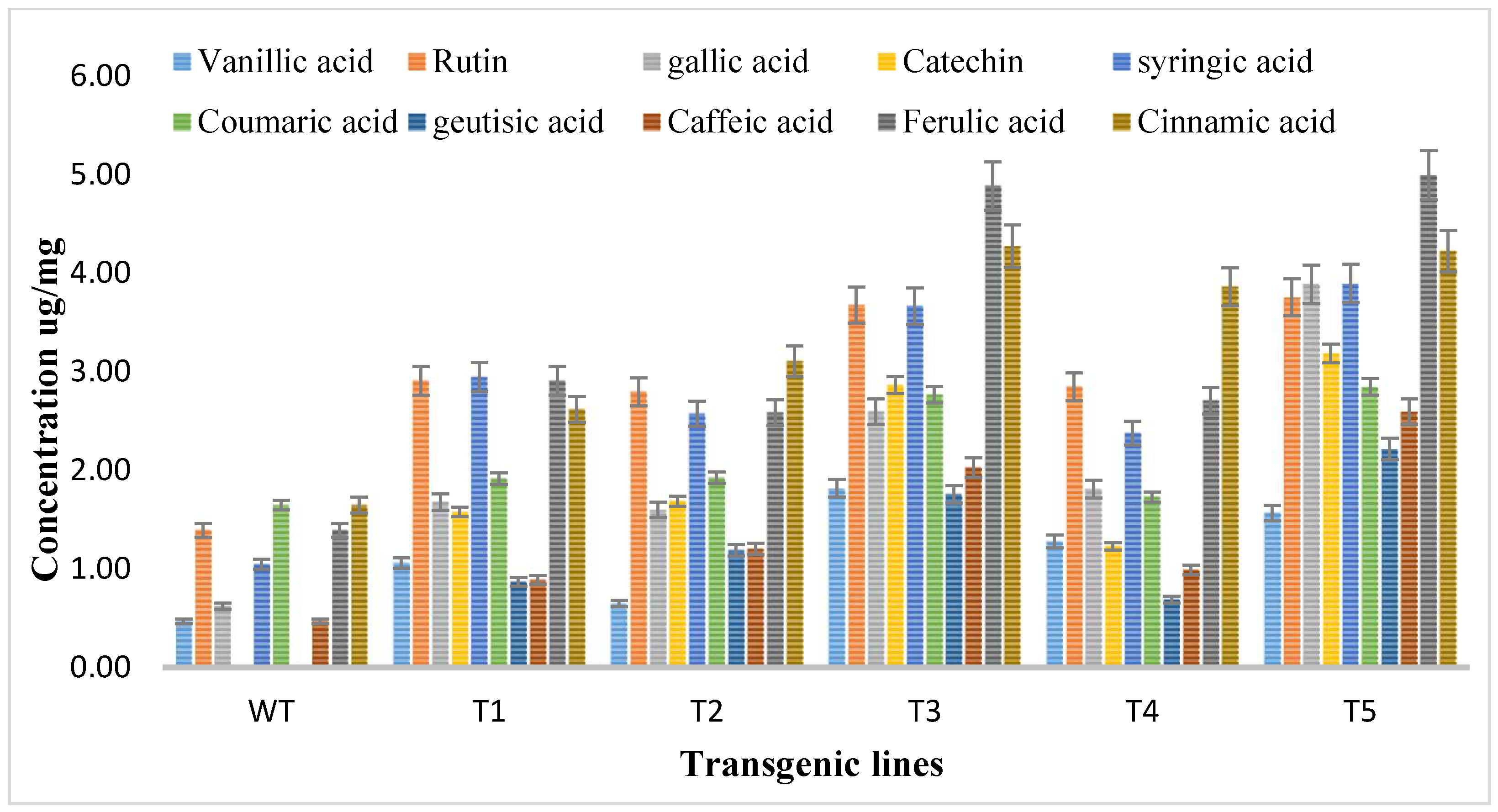
3.4. HPLC-DAD-Based Quantification of Flavonoids
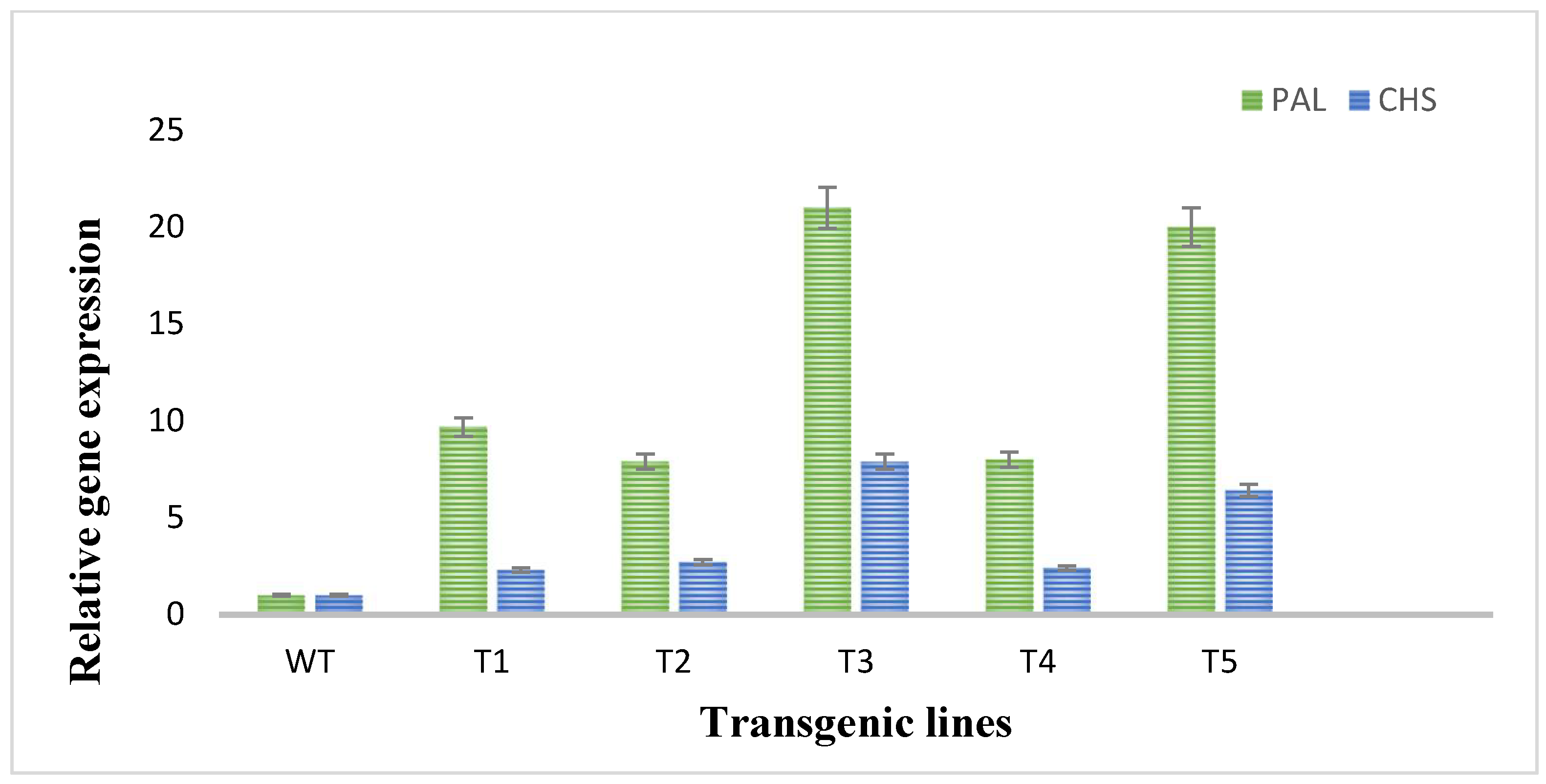
3.5. Expression Analysis of Flavonoid Biosynthetic Pathway Genes through Real-Time qPCR
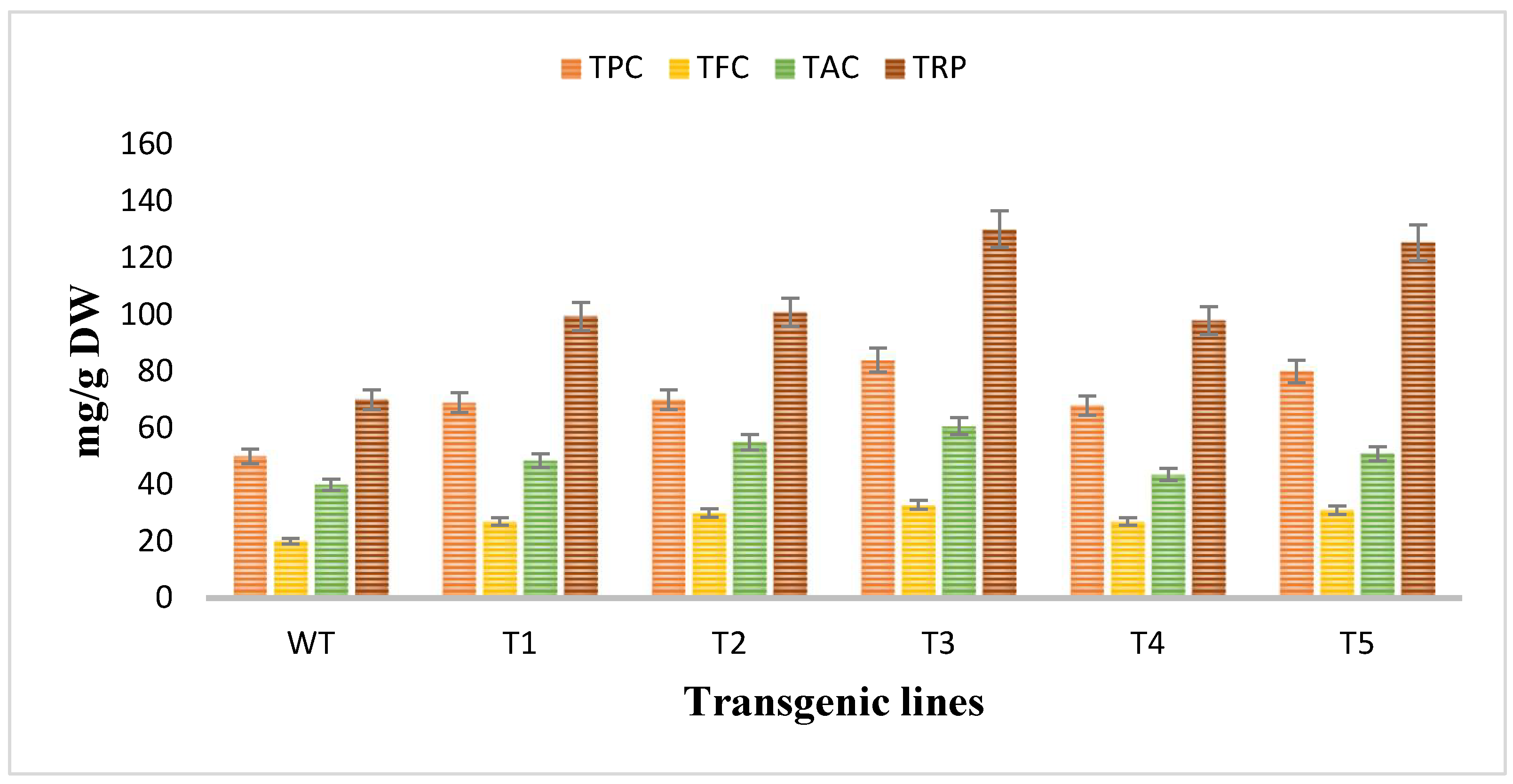
3.6. Analyzation of the Antioxidant Potential of Rol Gene Transformed and Untransformed A. carvifolia
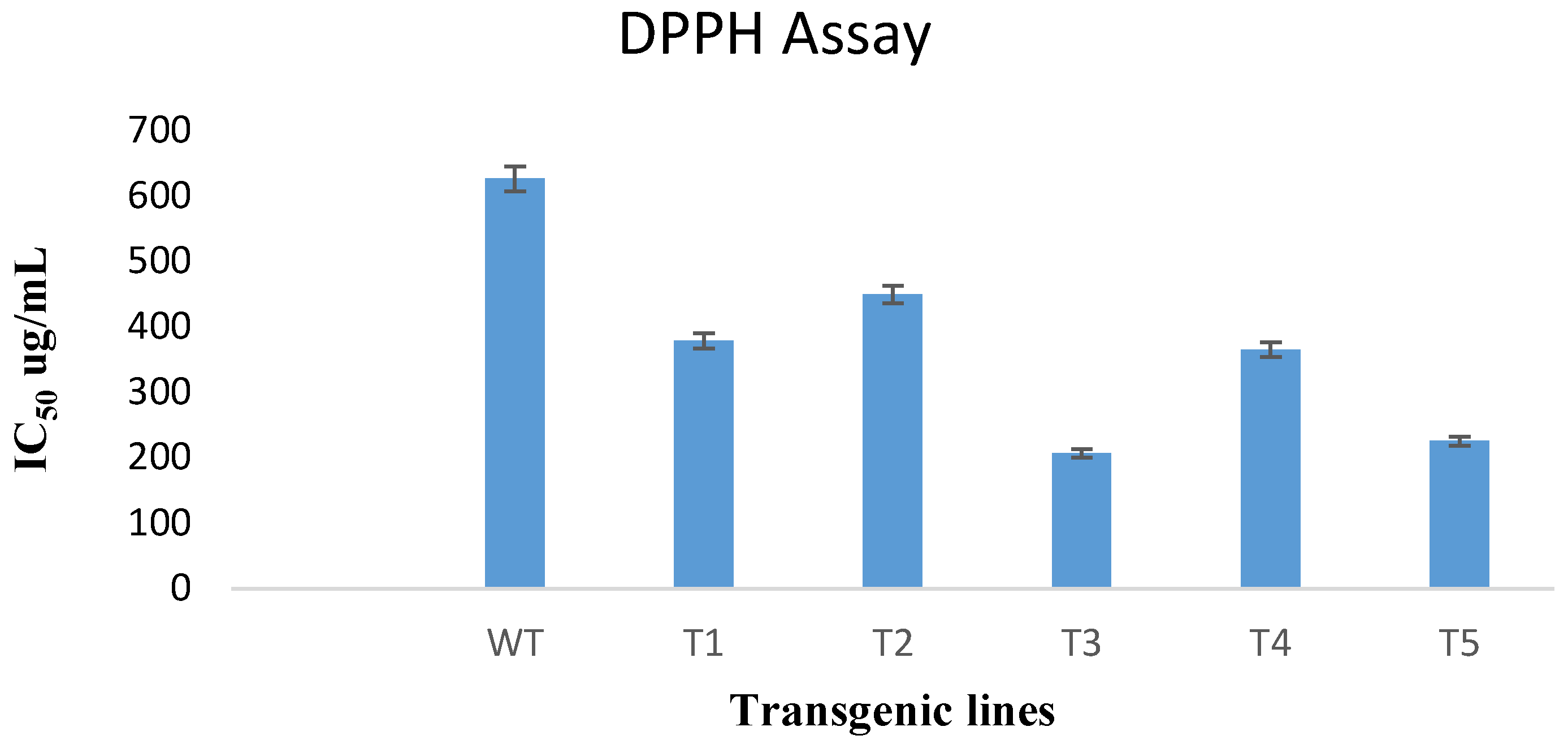
3.7. DPPH Free Radical Scavenging Assay
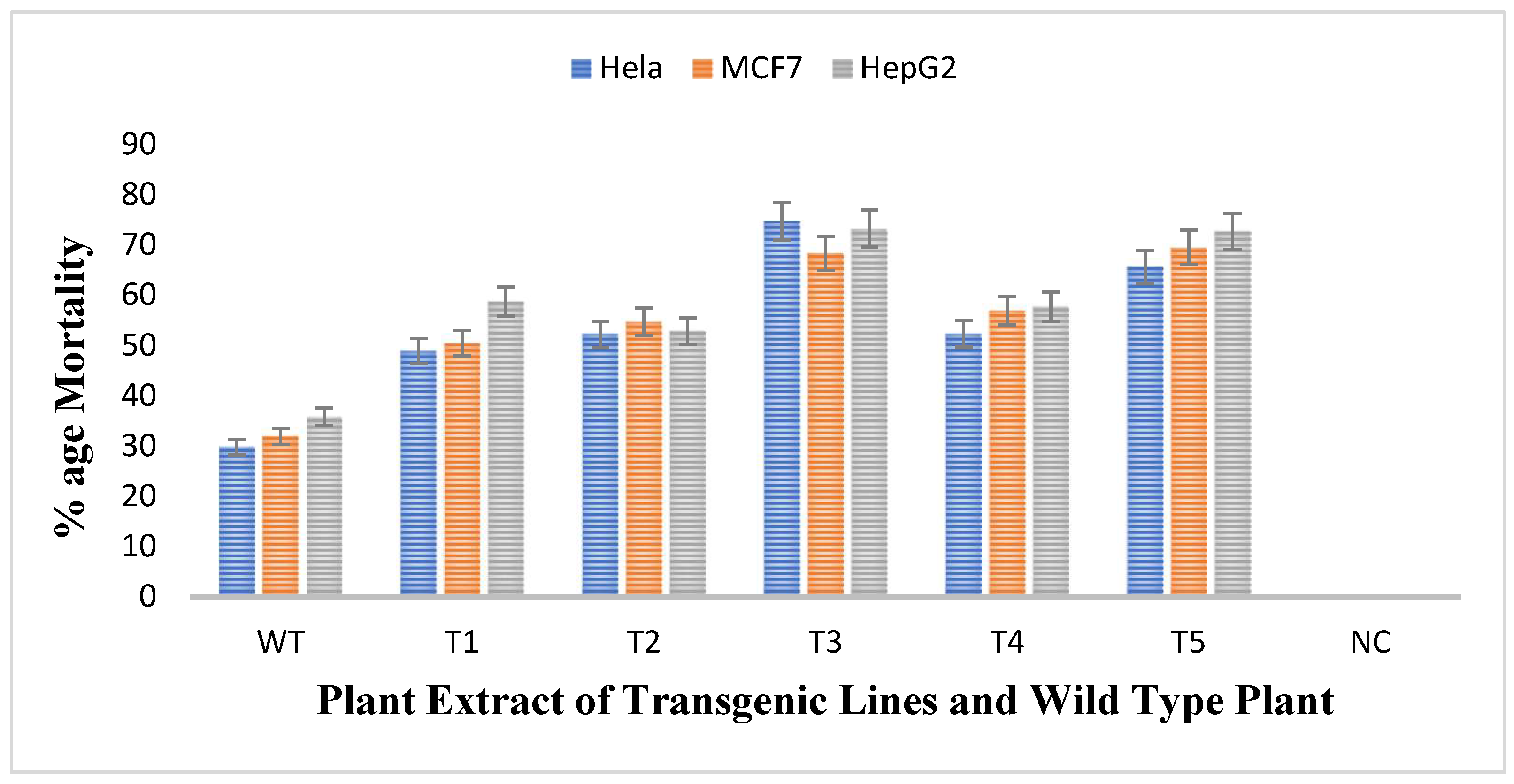
3.8. Measurement of Anticancerous Activity against Cancer Cell Lines through MTT Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussain, A. The genus Artemisia (Asteraceae): A review on its ethnomedicinal prominence and taxonomy with emphasis on foliar anatomy, morphology, and molecular phylogeny. Proc. Pak. Acad. Sci. B Life Environ. Sci. 2020, 57, 1–28. [Google Scholar]
- Abad Martínez, M.J.; Bedoya del Olmo, L.M.; Apaza Ticona, L.N.; Bermejo Benito, P. The Artemisia L. Genus: A Review of Bioactive Essential Oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Hayat, M.Q.; Sahreen, S.; ul Ain, Q.; Bokhari, S.A. Pharmacological promises of genus Artemisia (Asteraceae): A review: Pharmacological promises of genus Artemisia. Proc. Pak. Acad. Sci. B Life Environ. Sci. 2017, 54, 265–287. [Google Scholar]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Cavaiuolo, M.; Cocetta, G.; Ferrante, A. The antioxidants changes in ornamental flowers during development and senescence. Antioxidants 2013, 2, 132–155. [Google Scholar] [CrossRef] [Green Version]
- Iwashina, T. Flavonoid function and activity to plants and other organisms. Biol. Sci. Space 2003, 17, 24–44. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, L.; Routaboul, J.-M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Du, H.; Wang, J.; Pu, Y.; Yang, C.; Yan, R.; Yang, H.; Cheng, H.; Yu, D. Multiplex CRISPR/Cas9-mediated metabolic engineering increases soya bean isoflavone content and resistance to soya bean mosaic virus. Plant Biotechnol. J. 2020, 18, 1384–1395. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Man, C.; Xie, Y.; Yan, J.; Chu, J.; Huang, J. A crucial role of GA-regulated flavonol biosynthesis in root growth of Arabidopsis. Mol. Plant 2019, 12, 521–537. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.F.; Luthria, D.L.; Sasaki, T.; Heyerick, A. Flavonoids from Artemisia annua L. as antioxidants and their potential synergism with artemisinin against malaria and cancer. Molecules 2010, 15, 3135–3170. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Verma, P.K.; Singh, G. Total phenolic, flavonoids and tannin contents in different extracts of Artemisia absinthium. J. Complement. Med. Res. 1970, 1, 101. [Google Scholar] [CrossRef]
- Hajdú, Z.; Martins, A.; Orbán-Gyapai, O.; Forgo, P.; Jedlinszki, N.; Máthé, I.; Hohmann, J. Xanthine oxidase-inhibitory activity and antioxidant properties of the methanol extract and flavonoids of Artemisia asiatica. Rec. Nat. Prod. 2014, 8, 299–302. [Google Scholar]
- Qnais, E.; Raad, D.; Bseiso, Y. Analgesic and anti-inflammatory effects of an extract and flavonoids from Artemisia Herba-Alba and their mechanisms of action. Neurophysiology 2014, 46, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Benedito, V.A.; Modolo, L.V. Introduction to Metabolic Genetic Engineering for the Production of Valuable Secondary Metabolites in in vivo and in vitro Plant Systems. Recent Pat. Biotechnol. 2014, 8, 61–75. [Google Scholar] [CrossRef]
- Bulgakov, V.P. Functions of rol genes in plant secondary metabolism. Biotechnol. Adv. 2008, 26, 318–324. [Google Scholar] [CrossRef]
- Filippini, F.; Rossi, V.; Marin, O.; Trovato, M.; Costantino, P.; Mark Downey, P.; Lo Schiavo, F.; Terzi, M. A plant oncogene as a phosphatase. Nature 1996, 379, 499–500. [Google Scholar] [CrossRef]
- Filippini, F.; Schiavo, F.L.; Terzi, M.; Costantino, P.; Trovato, M. The plant oncogene rolB alters binding of auxin to plant cell membranes. Plant Cell Physiol. 1994, 35, 767–771. [Google Scholar] [CrossRef]
- Kiselev, K.; Dubrovina, A.; Veselova, M.; Bulgakov, V.; Fedoreyev, S.; Zhuravlev, Y.N. The rolB gene-induced overproduction of resveratrol in Vitis amurensis transformed cells. J. Biotechnol. 2007, 128, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Shkryl, Y.N.; Veremeichik, G.N.; Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. Individual and combined effects of the rolA, B, and C genes on anthraquinone production in Rubia cordifolia transformed calli. Biotechnol. Bioeng. 2008, 100, 118–125. [Google Scholar] [CrossRef]
- Bulgakov, V.; Tchernoded, G.; Mischenko, N.; Shkryl, Y.N.; Glazunov, V.; Fedoreyev, S.; Zhuravlev, Y.N. Effects of Ca2+ channel blockers and protein kinase/phosphatase inhibitors on growth and anthraquinone production in Rubia cordifolia callus cultures transformed by the rolB and rolC genes. Planta 2003, 217, 349–355. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Veselova, M.; Tchernoded, G.; Kiselev, K.; Fedoreyev, S.; Zhuravlev, Y.N. Inhibitory effect of the Agrobacterium rhizogenes rolC gene on rabdosiin and rosmarinic acid production in Eritrichium sericeum and Lithospermum erythrorhizon transformed cell cultures. Planta 2005, 221, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Dilshad, E.; Cusido, R.M.; Estrada, K.R.; Bonfill, M.; Mirza, B. Genetic transformation of Artemisia carvifolia Buch with rol genes enhances artemisinin accumulation. PLoS ONE 2015, 10, e0140266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilshad, E.; Cusido, R.M.; Palazon, J.; Estrada, K.R.; Bonfill, M.; Mirza, B. Enhanced artemisinin yield by expression of rol genes in Artemisia annua. Malar. J. 2015, 14, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazón, J.; Cusidó, R.; Roig, C.; Pinol, M. Expression of the rolC gene and nicotine production in transgenic roots and their regenerated plants. Plant Cell Rep. 1998, 17, 384–390. [Google Scholar]
- Ahmed, I.; Islam, M.; Arshad, W.; Mannan, A.; Ahmad, W.; Mirza, B. High-quality plant DNA extraction for PCR: An easy approach. J. Appl. Genet. 2009, 50, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Yunheng, J. psbA-trnH sequence analysis from chloroplast on medicinal plants of Artemisia. Chin. Agric. Sci. Bull. 2009, 25, 46–49. [Google Scholar]
- Spena, A.; Schmülling, T.; Koncz, C.; Schell, J. Independent and synergistic activity of rol A, B and C loci in stimulating abnormal growth in plants. EMBO J. 1987, 6, 3891–3899. [Google Scholar] [CrossRef]
- Kiani, B.H.; Safdar, N.; Mannan, A.; Mirza, B. Comparative Artemisinin analysis in Artemisia dubia transformed with two different Agrobacteria harbouring rol ABC genes. Plant Omics 2012, 5, 386–391. [Google Scholar]
- Sanbrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: Laurel Hollow, NY, USA, 1989; Volume 11, p. 31. [Google Scholar]
- Zafar, S.; Dilshad, E.; Ismail, H.; Rizvi, C.B.; Mirza, B. Rol genes enhance content of artemisinin and other secondary metabolites in Shennong hybrid of Artemisia annua. Chin. Herb. Med. 2019, 11, 209–215. [Google Scholar] [CrossRef]
- Dilshad, E.; Ismail, H.; Haq, I.-u.; Cusido, R.M.; Palazon, J.; Ramirez-Estrada, K.; Mirza, B. Rol genes enhance the biosynthesis of antioxidants in Artemisia carvifolia Buch. BMC Plant Biol. 2016, 16, 125. [Google Scholar] [CrossRef] [Green Version]
- Ul-Haq, I.; Ullah, N.; Bibi, G.; Kanwal, S.; Ahmad, M.S.; Mirza, B. Antioxidant and cytotoxic activities and phytochemical analysis of Euphorbia wallichii root extract and its fractions. Iran. J. Pharm. Res. IJPR 2012, 11, 241. [Google Scholar] [PubMed]
- Dilshad, E.; Zafar, S.; Ismail, H.; Waheed, M.T.; Cusido, R.M.; Palazon, J.; Mirza, B. Effect of rol genes on polyphenols biosynthesis in Artemisia annua and their effect on antioxidant and cytotoxic potential of the plant. Appl. Biochem. Biotechnol. 2016, 179, 1456–1468. [Google Scholar] [CrossRef] [PubMed]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction--antioxidative activities of products of browning reaction prepared from glucosamine. Eiyogaku Zasshi Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Rani, R. Agrobacterium Mediated Transformation and Secondary Metabolite Analysis of Ajuga bracteosa Wall. ex. Benth. Ph.D. Thesis, Quaid-I-Azam University, Islamabad, Pakistan, 2015. [Google Scholar]
- Schmülling, T.; Schell, J.; Spena, A. Promoters of the rolA, B, and C genes of Agrobacterium rhizogenesare differentially regulated in transgenic plants. Plant Cell 1989, 1, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Sinkar, V.P.; Pythoud, F.; White, F.F.; Nester, E.W.; Gordon, M.P. rolA locus of the Ri plasmid directs developmental abnormalities in transgenic tobacco plants. Genes Dev. 1988, 2, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, T.; Spena, A. The plant oncogenes rolA, B, and C from Agrobacterium rhizogenes. In Agrobacterium Protocols; Springer: Berlin/Heidelberg, Germany, 1995; pp. 207–222. [Google Scholar]
- Dehio, C.; Grossmann, K.; Schell, J.; Schmülling, T. Phenotype and hormonal status of transgenic tobacco plants overexpressing the rolA gene of Agrobacterium rhizogenes T-DNA. Plant Mol. Biol. 1993, 23, 1199–1210. [Google Scholar] [CrossRef]
- Zia, M.; Mirza, B.; Malik, S.A.; Chaudhary, M.F. Expression of rol genes in transgenic soybean (Glycine max L.) leads to changes in plant phenotype, leaf morphology, and flowering time. Plant Cell Tissue Organ Cult. (PCTOC) 2010, 103, 227–236. [Google Scholar] [CrossRef]
- Ismail, H.; Dilshad, E.; Waheed, M.T.; Sajid, M.; Kayani, W.K.; Mirza, B. Transformation of Lactuca sativa L. with rol C gene results in increased antioxidant potential and enhanced analgesic, anti-inflammatory and antidepressant activities in vivo. 3 Biotech 2016, 6, 215. [Google Scholar] [CrossRef] [Green Version]
- Kiani, B.H. Transfer of Rol Genes and Evaluation of Artemisinin Synthesis in Transgenic Artemisia annua L. and Artemisia Dubia Wall. Ph.D. Thesis, Quaid-i-Azam University, Islamabad, Pakistan, 2012. [Google Scholar]
- Schmülling, T.; Fladung, M.; Grossmann, K.; Schell, J. Hormonal content and sensitivity of transgenic tobacco and potato plants expressing single rol genes of Agrobacterium rhizogenes T-DNA. Plant J. 1993, 3, 371–382. [Google Scholar] [CrossRef]
- Bensaddek, L.; Villarreal, M.L.; Fliniaux, M.-A. Induction and growth of hairy roots for the production of medicinal compounds. Electron. J. Integr. Biosci. 2008, 3, 2–9. [Google Scholar]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Production of glucosinolates, phenolic compounds and associated gene expression profiles of hairy root cultures in turnip (Brassica rapa ssp. rapa). 3 Biotech 2016, 6, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.A.; Elkelish, A.; Elansary, H.O.; Ali, H.M.; Elshikh, M.; Witczak, J.; Ahmad, M. Genetic transformation and hairy root induction enhance the antioxidant potential of Lactuca serriola L. Oxidative Med. Cell. Longev. 2017, 2017, 5604746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijveldt, R.J.; Van Nood, E.; Van Hoorn, D.E.; Boelens, P.G.; Van Norren, K.; Van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spies, L.; Koekemoer, T.; Sowemimo, A.; Goosen, E.; Van de Venter, M. Caspase-dependent apoptosis is induced by Artemisia afra Jacq. ex Willd in a mitochondria-dependent manner after G2/M arrest. S. Afr. J. Bot. 2013, 84, 104–109. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.L.; Charles, S.V. Antioxidant, antibacterial and cytotoxic activities of essential oils and ethanol extracts of selected South East Asian herbs. J. Med. Plants Res. 2011, 5, 5284–5290. [Google Scholar]
- Singh, N.P.; Ferreira, J.F.; Park, J.S.; Lai, H.C. Cytotoxicity of ethanolic extracts of Artemisia annua to Molt-4 human leukemia cells. Planta Med. 2011, 77, 1788–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Standard Flavonoids | Signal Wavelength (nm) | Retention Time (min) |
|---|---|---|---|
| 1 | Vanillic acid | 257 | 9.116 |
| 2 | Rutin | 257 | 12.649 |
| 3 | Gallic acid | 279 | 3.679 |
| 4 | Catechin | 279 | 7.003 |
| 5 | syringic acid | 279 | 9.76 |
| 6 | Coumaric acid | 279 | 13.79 |
| 7 | Geutisic acid | 325 | 7.433 |
| 8 | Caffeic acid | 325 | 9.252 |
| 9 | Ferulic acid | 325 | 12.642 |
| 10 | Cinnanic acid | 325 | 13.79 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | Prob. |
|---|---|---|---|---|---|
| Polyphenols | 9 | 117.4 | 13.04 | 54.64 | 0.0000 |
| Transgenic lines | 5 | 120.2 | 24.04 | 1592 | 0.0000 |
| Interaction | 45 | 20.15 | 0.4478 | 2933 | 0.0000 |
| Residual (error) | 120 | 0.9835 | 0.008196 | ||
| Total | 179 | 258.7 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | Prob. |
|---|---|---|---|---|---|
| Interaction | 15 | 3330 | 222.0 | 68.68 | 0.0000 |
| Transgenic lines | 5 | 7898 | 1580 | 488.8 | 0.0000 |
| Antioxidant assays | 3 | 56270 | 18760 | 5804 | 0.0000 |
| Residual (error) | 48 | 155.1 | 3.232 | ||
| Total | 71 | 67660 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | Prob. |
|---|---|---|---|---|---|
| Interaction | 15 | 472.6 | 31.51 | 17.32 | 0.0000 |
| Transgenic lines | 5 | 3281 | 656.2 | 360.7 | 0.0000 |
| Concentrations | 3 | 22130 | 7376 | 4055 | 0.0000 |
| Residual (error) | 48 | 87.33 | 1.819 | ||
| Total | 71 | 25970 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | Prob. |
|---|---|---|---|---|---|
| Cancer cell lines | 2 | 202.2 | 101.1 | 118.2 | 0.0000 |
| Transgenic lines | 6 | 34110 | 5684 | 6645 | 0.0000 |
| Interaction | 12 | 291.4 | 24.28 | 28.38 | 0.0000 |
| Residual (error) | 42 | 35.93 | 0.8554 | ||
| Total | 62 | 34640 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.N.; Dilshad, E. Enhanced Antioxidant and Anticancer Potential of Artemisia carvifolia Buch Transformed with rol A Gene. Metabolites 2023, 13, 351. https://doi.org/10.3390/metabo13030351
Khan AN, Dilshad E. Enhanced Antioxidant and Anticancer Potential of Artemisia carvifolia Buch Transformed with rol A Gene. Metabolites. 2023; 13(3):351. https://doi.org/10.3390/metabo13030351
Chicago/Turabian StyleKhan, Amna Naheed, and Erum Dilshad. 2023. "Enhanced Antioxidant and Anticancer Potential of Artemisia carvifolia Buch Transformed with rol A Gene" Metabolites 13, no. 3: 351. https://doi.org/10.3390/metabo13030351
APA StyleKhan, A. N., & Dilshad, E. (2023). Enhanced Antioxidant and Anticancer Potential of Artemisia carvifolia Buch Transformed with rol A Gene. Metabolites, 13(3), 351. https://doi.org/10.3390/metabo13030351