
Ethanolic Extract of Artemisia vulgaris Leaf Promotes Apoptotic Cell Death in Non-Small-Cell Lung Carcinoma A549 Cells through Inhibition of the Wnt Signaling Pathway

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Plant Sample and Extract Preparation
2.2.2. Cell Culture
2.2.3. GC-MS Analysis
2.2.4. Cell Viability Assay
2.2.5. Morphological Evaluation
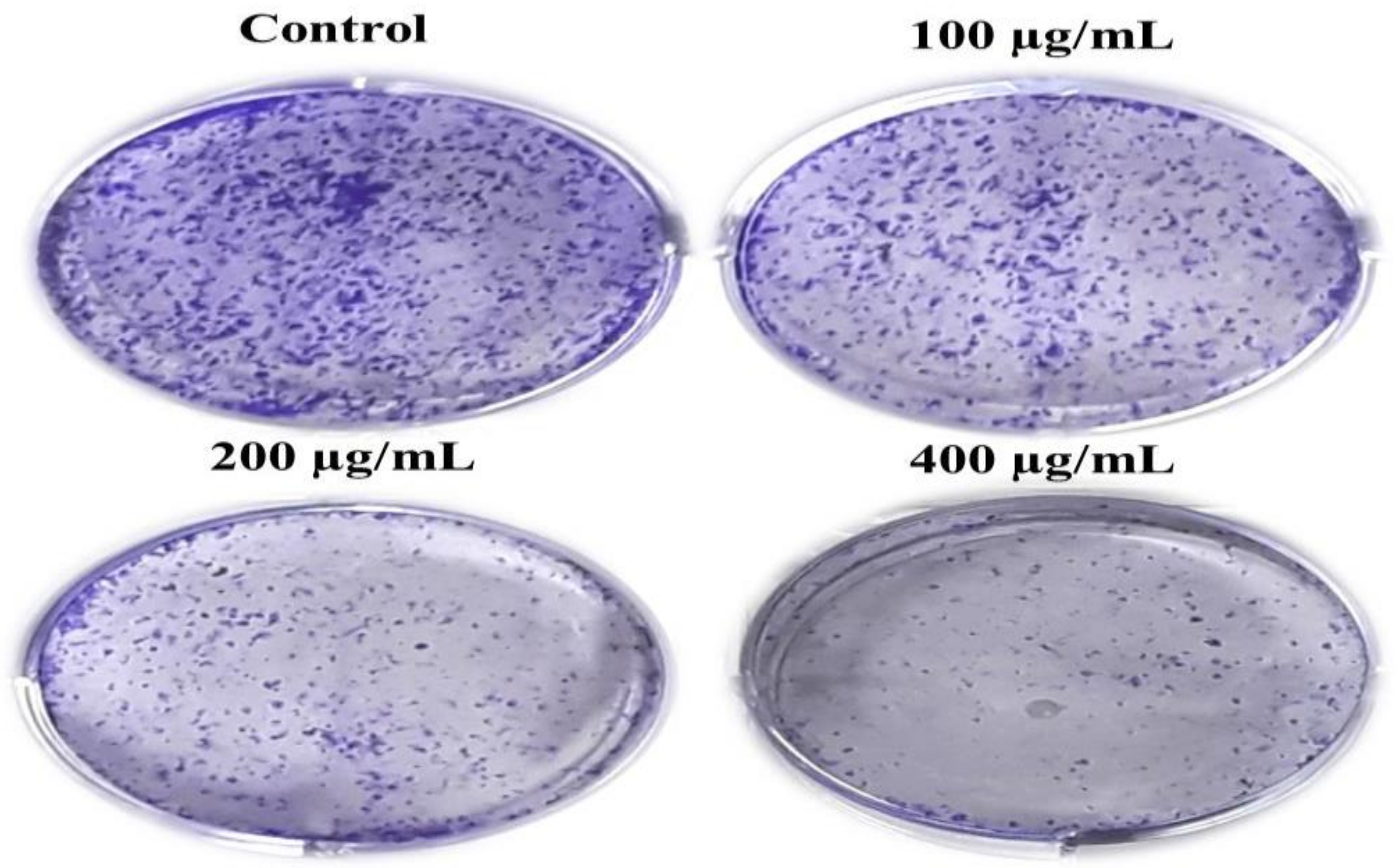
2.2.6. Colony Formation Assay
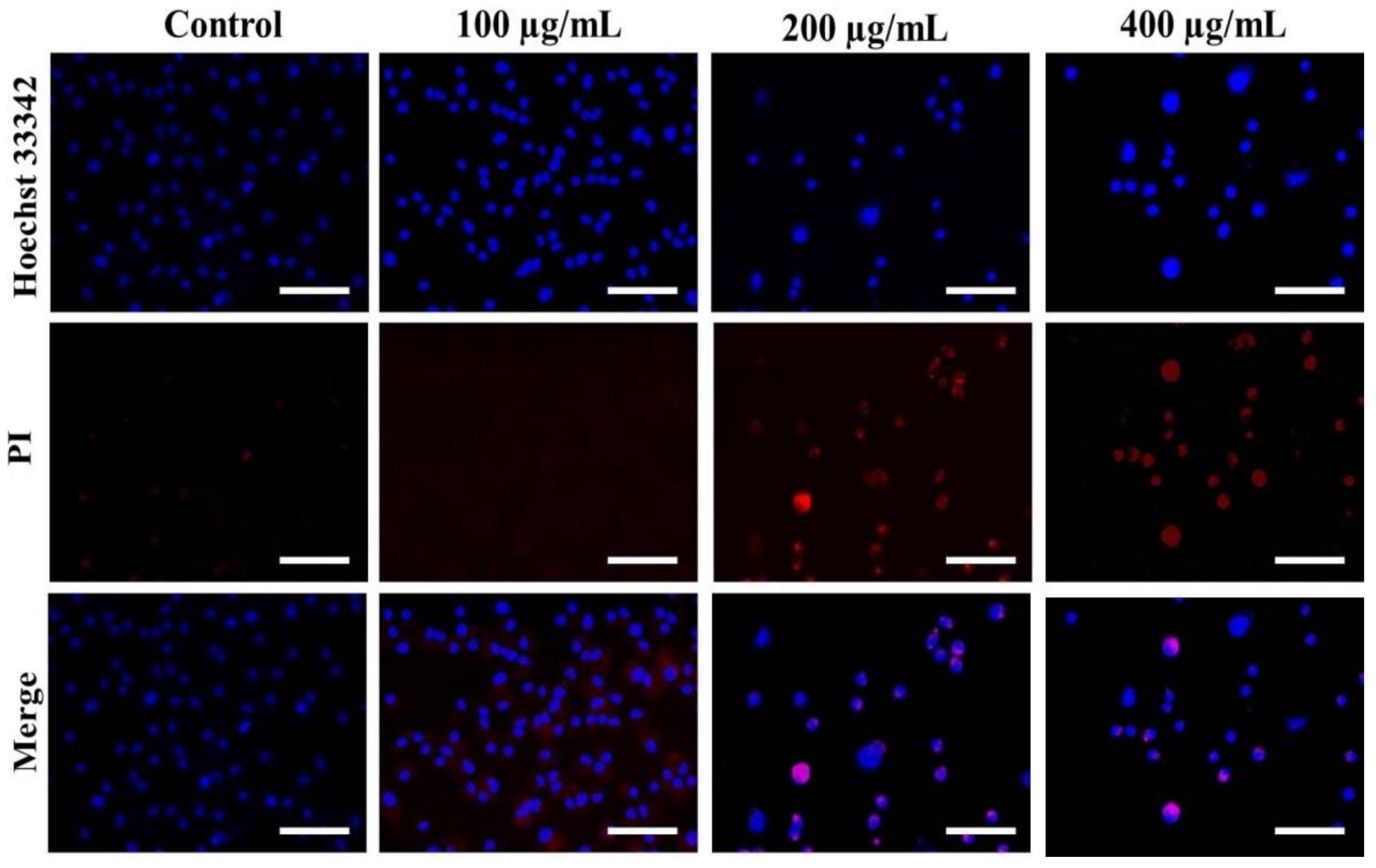
2.2.7. Hoechst/PI Staining
2.2.8. Measurement of Caspase-8, -9, and -3 Activity
2.2.9. Mitochondrial Membrane Potential Quantification
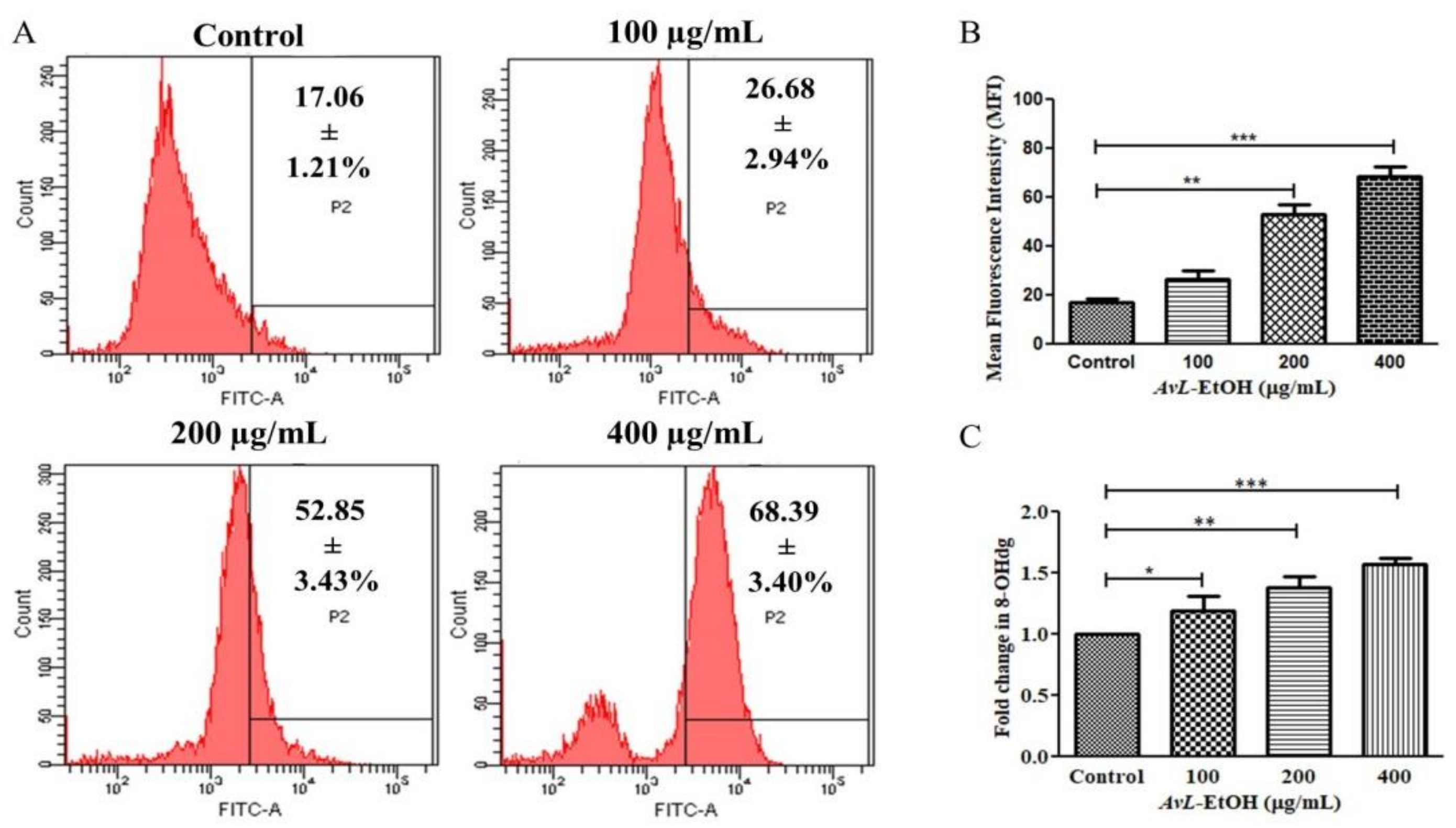
2.2.10. Determination of Intracellular Reactive Oxygen Species Levels
2.2.11. Quantification of Apoptosis
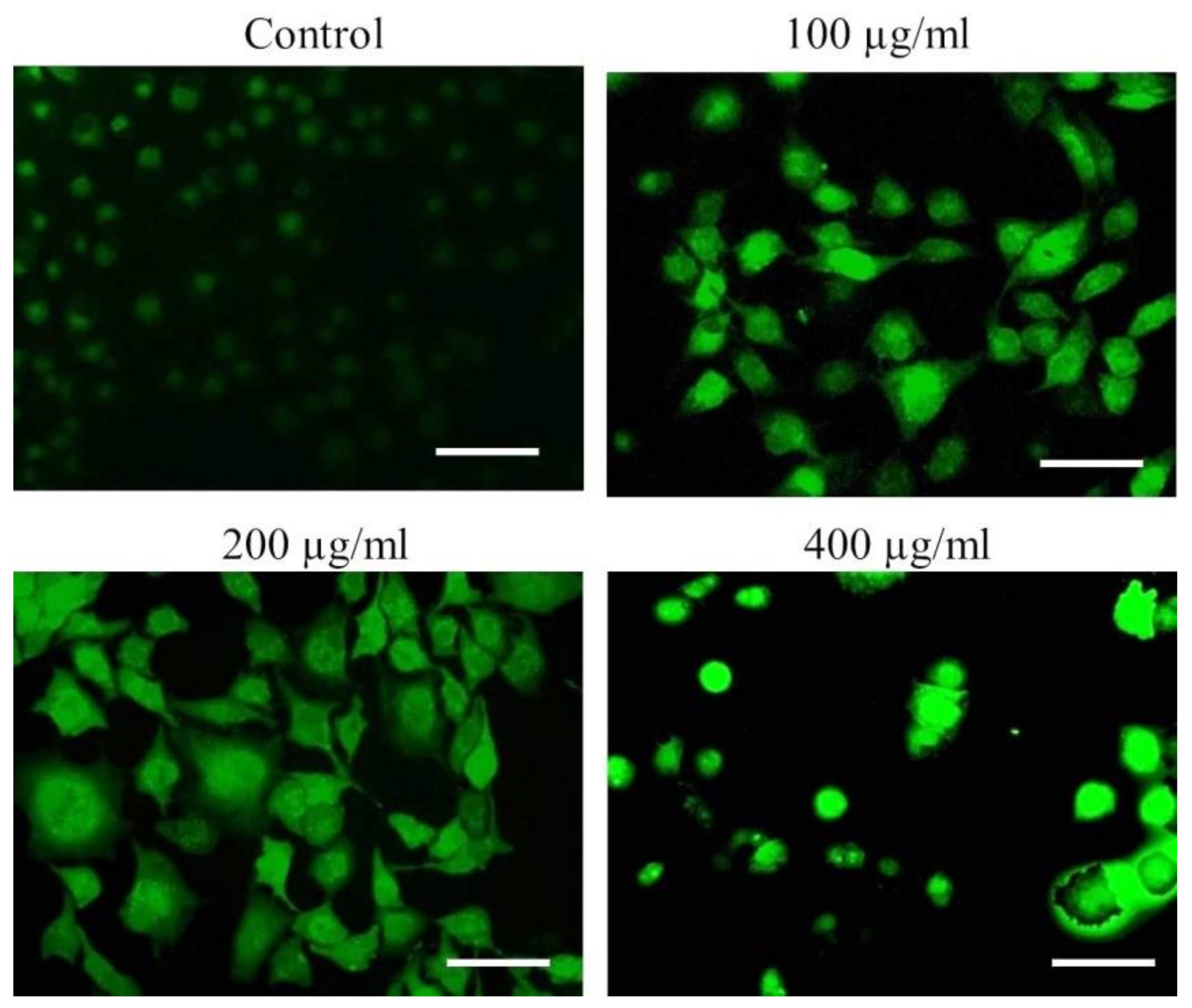
2.2.12. DNA Damage Assay
2.2.13. Quantification of Cytosolic Cytochrome-c Levels
2.2.14. Quantification of PARP Cleavage
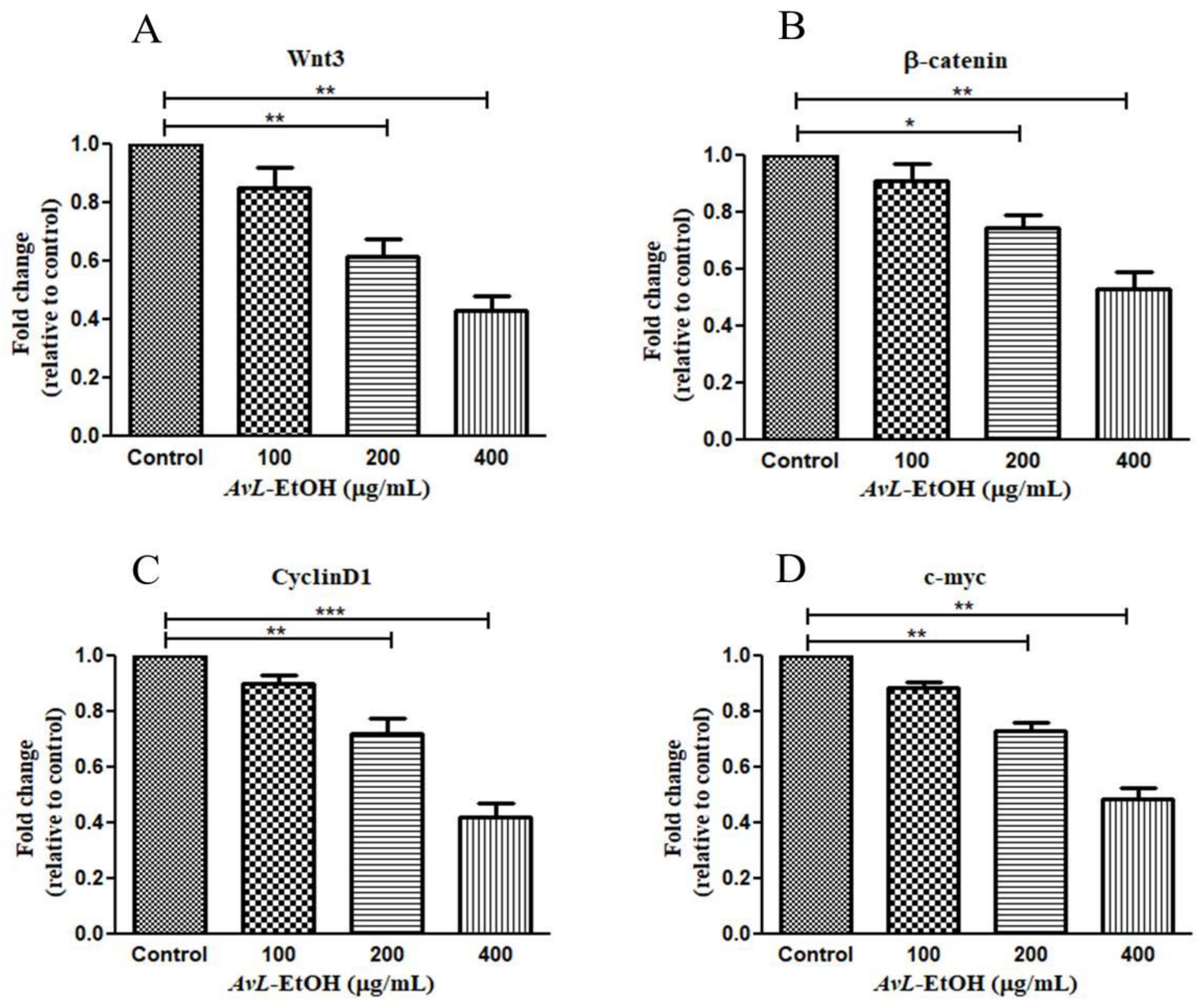
2.2.15. Reverse Transcriptase qPCR Evaluations
2.2.16. Statistical Analysis
3. Results
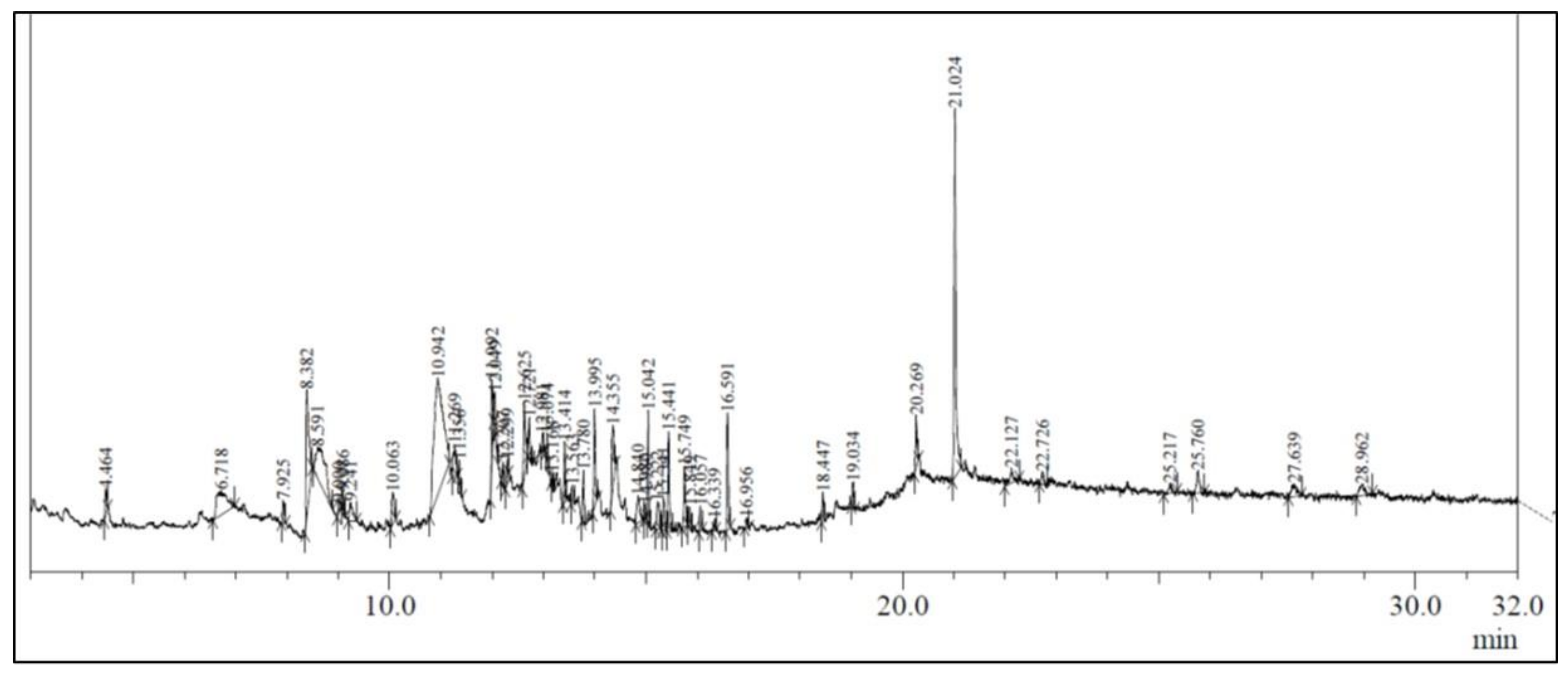
3.1. GC-MS Analysis of Ethanolic Extract of A. vulgaris
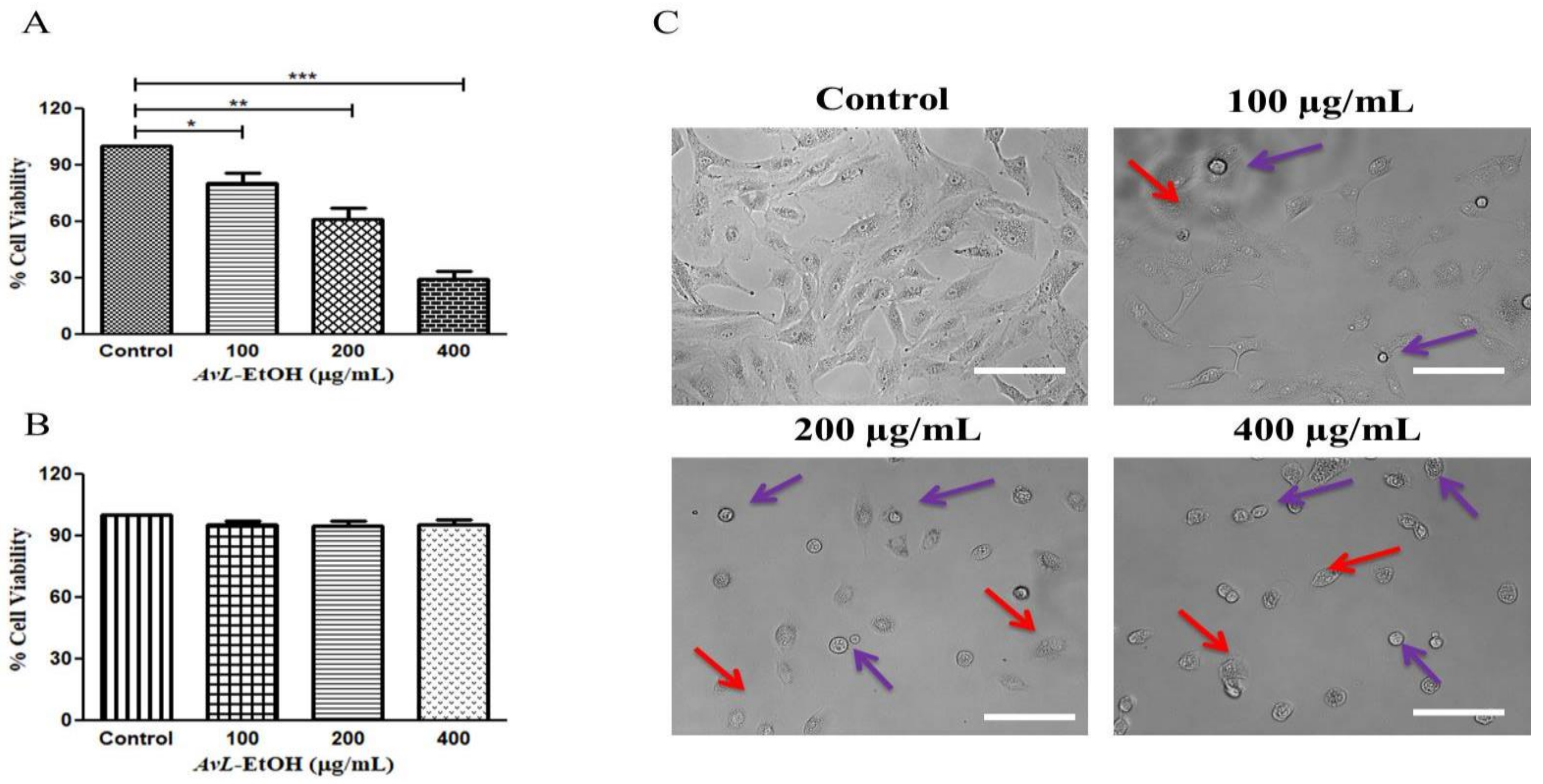
3.2. AvL-EtOH Inhibits the Cell Viability of A549 Cells
3.3. Nuclear Condensation in A549 Cells after AvL-EtOH Exposure
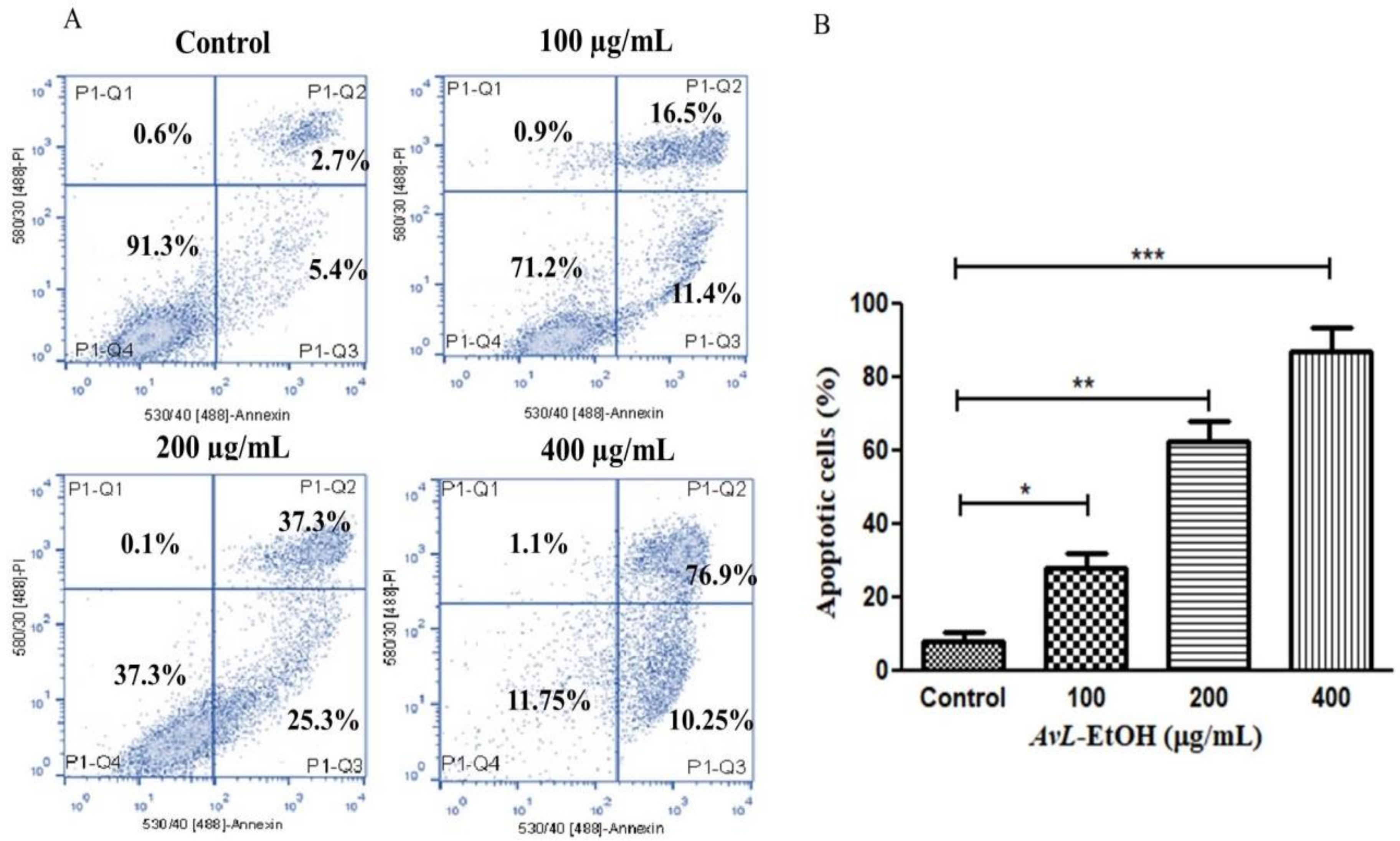
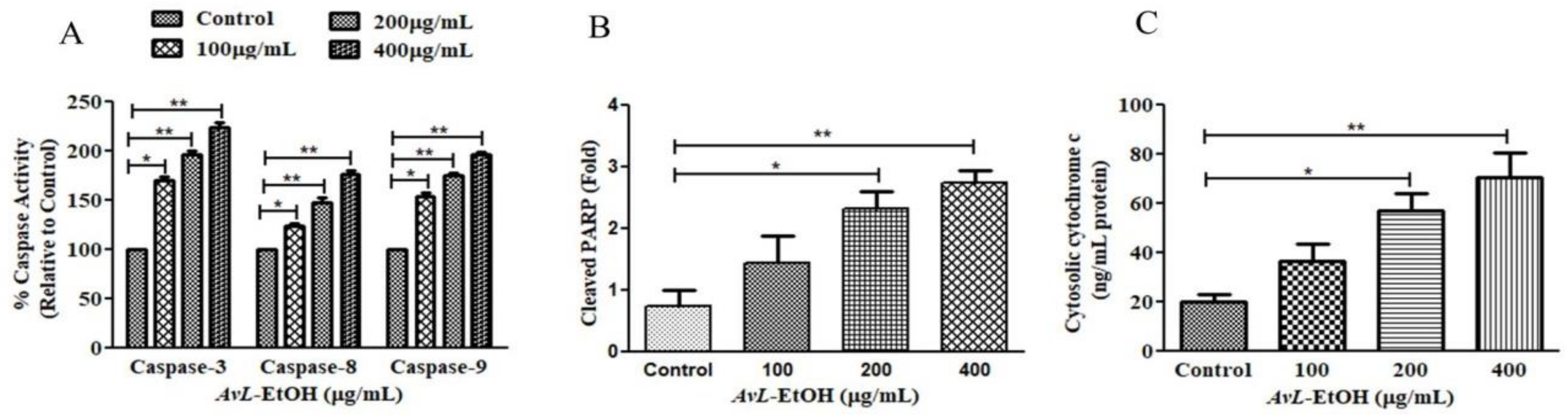
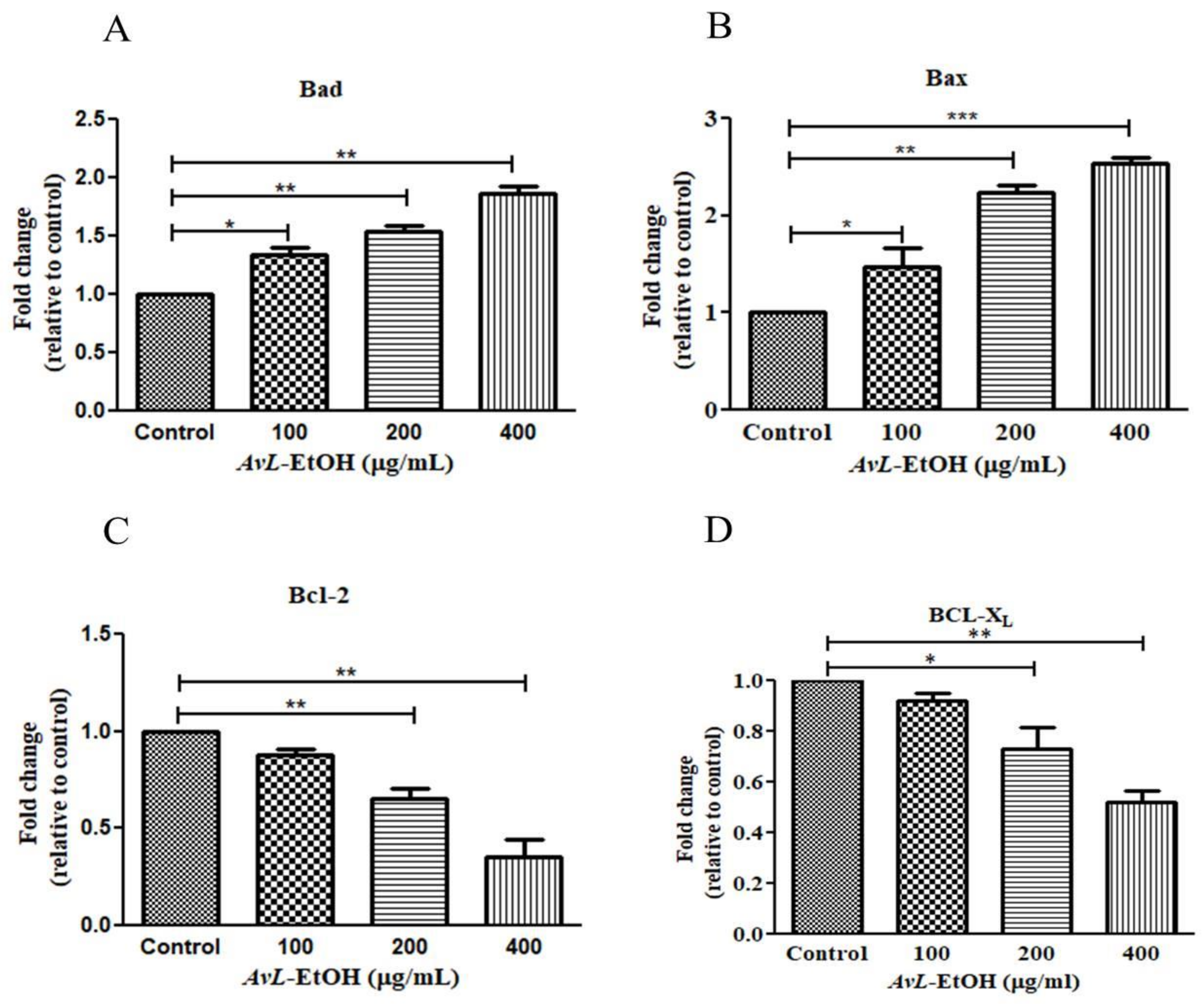
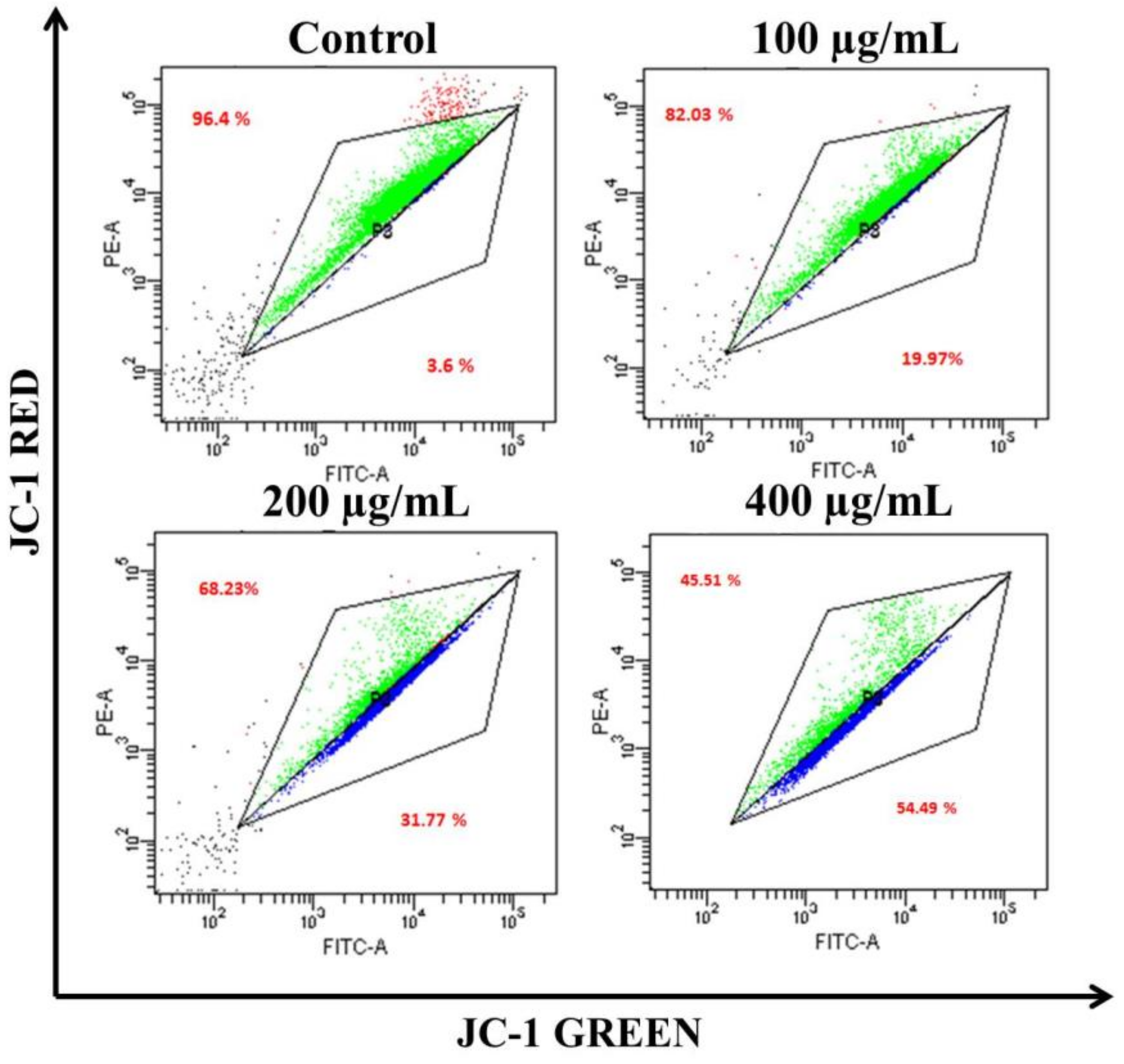
3.4. AvL-EtOH Induced Apoptosis through Activation of Intrinsic as well as Extrinsic Pathway
3.5. Modulation of Wnt/β-Catenin Signaling Pathway by AvL-EtOH
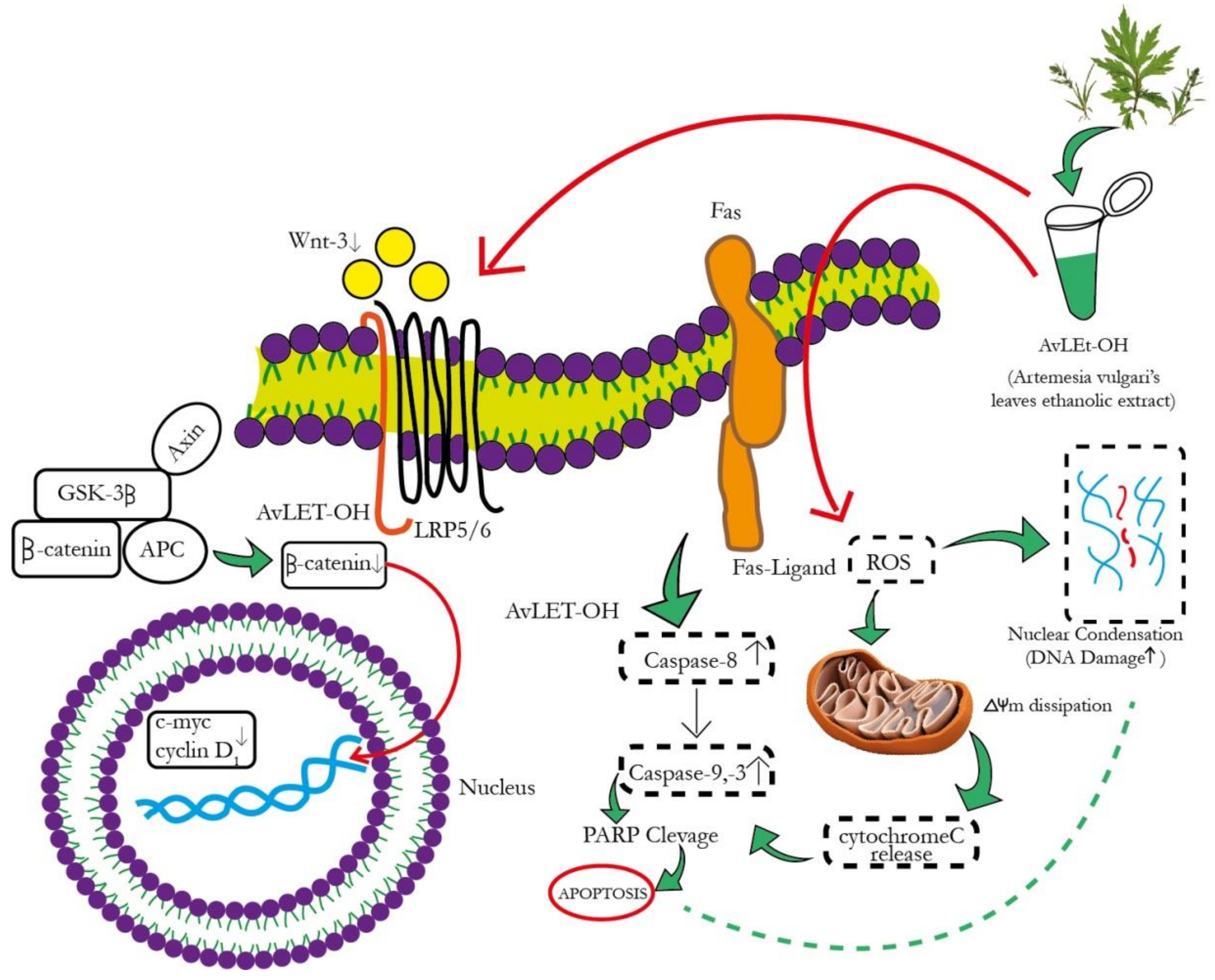
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Cancer Observatory, Cancer Factsheet. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/15-Lung-fact-sheet.pdf (accessed on 25 August 2022).
- Batbold, U.; Liu, J.J. Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators. Appl. Sci. 2022, 12, 4868. [Google Scholar] [CrossRef]
- Hardy, D.; Cormier, J.N.; Xing, Y.; Liu, C.C.; Xia, R.; Du, X.L. Chemotherapy-associated toxicity in a large cohort of elderly patients with non-small cell lung cancer. J. Thorac. Oncol. 2010, 5, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Longley, D.B.; Johnston, P.G. Molecular mechanisms of drug resistance. J. Pathol. 2005, 205, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Bahadur, S.; Harwansh, R.; Biswas, S.; Banerjee, S. Paradigm shift in natural product research: Traditional medicine inspired approaches. Phytochem. Rev. 2016, 16, 803–826. [Google Scholar] [CrossRef]
- Venkatesha, S.H.; Berman, B.M.; Moudgil, K.D. Herbal medicinal products target defined biochemical and molecular mediators of inflammatory autoimmune arthritis. Bioorg. Med. Chem. 2011, 19, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koni, M.; Pinnarò, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int. J. Mol. Sci. 2020, 21, 7697. [Google Scholar] [CrossRef]
- Hafeez, B.B.; Siddiqui, I.A.; Asim, M.; Malik, A.; Afaq, F.; Adhami, V.M.; Saleem, M.; Din, M.; Mukhtar, H. A dietary anthocyanidin delphinidin induces apoptosis of human prostate cancer PC3 cells in vitro and in vivo: Involvement of nuclear factor-kappaB signaling. Cancer Res. 2008, 68, 8564–8572. [Google Scholar] [CrossRef] [Green Version]
- Du, J.H.; Zhang, H.D.; Ma, Z.J.; Ji, K.M. Artesunate induces oncosis-like cell death in vitro and has antitumor activity against pancreatic cancer xenografts in vivo. Cancer Chemother. Pharmacol. 2010, 65, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M.; Shimada, H.; Matsuda, H.; Yamahara, J.; Murakami, N. Bioactive constituents of Chinese natural medicines. I. New sesquiterpene ketones with vasorelaxant effect from Chinese moxa, the processed leaves of Artemisia argyi Levl. et Vant.: Moxartenone and moxartenolide. Chem. Pharm. Bull. 1996, 44, 1656–1662. [Google Scholar] [CrossRef] [Green Version]
- Afsar, S.K.; Kumar, K.R.; Gopal, J.V.; Raveesha, P. Assessment of antiinfammatory activity of Artemisia vulgaris leaves by cotton pellet granuloma method in Wistar albino rats. J. Pharm. Res. 2013, 7, 463–467. [Google Scholar]
- Jakovljević, M.R.; Grujičić, D.; Vukajlović, J.T.; Marković, A.; Milutinović, M.; Stanković, M.; Milošević-Djordjević, O. In vitro study of genotoxic and cytotoxic activities of methanol extracts of Artemisia vulgaris L. and Artemisia alba Turra. S. Afr. J. Bot. 2020, 132, 117–126. [Google Scholar] [CrossRef]
- Ahmad, A.; Tiwari, R.K.; Almeleebia, T.M.; Al Fayi, M.S.; Alshahrani, M.Y.; Ahmad, I.; Abohassan, M.S.; Saeed, M.; Ansari, I.A. Swertia chirayita suppresses the growth of non-small cell lung cancer A549 cells and concomitantly induces apoptosis via downregulation of JAK1/STAT3 pathway. Saudi J. Biol. Sci. 2021, 28, 6279–6288. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Singh, S.; Gupta, C.L.; Bajpai, P. Microglial TLR9: Plausible Novel Target for Therapeutic Regime Against Glioblastoma Multiforme. Cell Mol. Neurobiol. 2021, 41, 1391–1393. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.K.; Bhardwaj, R.; Dwarakanath, B.S.; Varshney, R. Metabolic oxidative stress induced by a combination of 2-DG and 6-AN enhances radiation damage selectively in malignant cells via non-coordinated expression of antioxidant enzymes. Cancer Lett. 2010, 295, 154–166. [Google Scholar] [CrossRef]
- Ahmad, A.; Tiwari, R.K.; Saeed, M.; Ahmad, I.; Ansari, I.A. Glycyrrhizin Mediates Downregulation of Notch Pathway Resulting in Initiation of Apoptosis and Disruption in the Cell Cycle Progression in Cervical Cancer Cells. Nutr Cancer. 2022, 74, 622–639. [Google Scholar] [CrossRef]
- Ahmad, A.; Ansari, I.A. Carvacrol Exhibits Chemopreventive Potential against Cervical Cancer Cells via Caspase-Dependent Apoptosis and Abrogation of Cell Cycle Progression. Anticancer Agents Med. Chem. 2021, 21, 2224–2235. [Google Scholar] [CrossRef]
- Pandey, B.P.; Thapa, R.; Upreti, A. Chemical composition, antioxidant and antibacterial activities of essential oil and methanol extract of Artemisia vulgaris and Gaultheria fragrantissima collected from Nepal. Asian Pac. J. Trop. Med. 2017, 10, 952–959. [Google Scholar] [CrossRef]
- Van Nguyen Thien, T.; Tran, L.T.K.; Nhu, N.T.T.; Duc, T.P.; Do, L.T.M.; Tu, D.D.; That, Q.T. A new eudesmane-type sesquiterpene from the leaves of Artemisia vulgaris. Chem. Nat. Compd. 2018, 54, 66–68. [Google Scholar] [CrossRef]
- Ekiert, H.; Pajor, J.; Klin, P.; Rzepiela, A.; Ślesak, H.; Szopa, A. Significance of Artemisia vulgaris L. (Common Mugwort) in the History of Medicine and Its Possible Contemporary Applications Substantiated by Phytochemical and Pharmacological Studies. Molecules 2020, 25, 4415. [Google Scholar] [CrossRef]
- Saleh, A.M.; Aljada, A.; Rizvi, S.A.; Nasr, A.; Alaskar, A.S.; Williams, J.D. In vitro cytotoxicity of Artemisia vulgaris L. essential oil is mediated by a mitochondria-dependent apoptosis in HL-60 leukemic cell line. BMC Complement. Altern. Med. 2014, 14, 226. [Google Scholar] [CrossRef] [Green Version]
- Vahdati-Mashhadian, N.; Emami, S.; Oghazian, M.; Vosough, R. The cytotoxicity evaluation of seven species of Artemisia on human tumor cell lines. Pharmacologyonline 2009, 1, 229–242. [Google Scholar]
- Rasheed, T.; Bilal, M.; Iqbal, H.M.N.; Li, C. Green biosynthesis of silver nanoparticles using leaves extract of Artemisia vulgaris and their potential biomedical applications. Colloids Surf. B Biointerfaces 2017, 158, 408–415. [Google Scholar] [CrossRef]
- Yip, H.Y.K.; Papa, A. Signaling Pathways in Cancer: Therapeutic Targets, Combinatorial Treatments, and New Developments. Cells 2021, 10, 659. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.; Greer, Y.; Lipkowitz, S.; Takebe, N. Novel Apoptosis-Inducing Agents for the Treatment of Cancer, a New Arsenal in the Toolbox. Cancers 2019, 11, 1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwell, M.; Rahman, P.K. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar]
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Fetrow, C.W.; Avila, J.R. Professional’s Handbook of Complementary & Alternative Medicines; Springhouse Publishing Company: Alphen aan den Rijn, Netherlands, 2001. [Google Scholar]
- Gruenwald, J.; Brendler, T.; Jaenicke, C. Physicians’ Desk Reference (PDR) for Herbal Medicines, 13th ed.; Thomson Medical Economics Company: Montvale, NJ, USA, 2004. [Google Scholar]
- Tang, J.; Zhao, J.J.; Li, Z.H. Ethanol extract of Artemisia sieversiana exhibits anticancer effects and induces apoptosis through a mitochondrial pathway involving DNA damage in COLO-205 colon carcinoma cells. Bangladesh J. Pharmacol. 2015, 10, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Emami, S.A.; Vahdati-Mashhadian, N.; Vosough, R.; Oghazian, M.B. The anticancer activity of five species of Artemisia on Hep2 and HepG2 cell lines. Pharmacologyonline 2009, 3, 327–339. [Google Scholar]
- Sharmila, K.; Padma, P.R. Anticancer activity of Artemisia vulgaris on hepatocellular carcinoma (HepG2) cells. Int. J. Pharm. Pharm. Sci. 2013, 5, 479–483. [Google Scholar]
- Abdelhamed, S.; Yokoyama, S.; Hafiyani, L.; Kalauni, S.K.; Hayakawa, Y.; Awale, S.; Saiki, I. Identification of plant extracts sensitizing breast cancer cells to TRAIL. Oncol. Rep. 2013, 29, 1991–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.C.; Deng, T.; Lehal, R.S.; Kim, J.; Zacksenhaus, E. Identification of tumorsphere- and tumor-initiating cells in HER2/Neu-induced mammary tumors. Cancer Res. 2007, 67, 8671–8681. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The artemisia L. Genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.Q.; Hu, J.; Yang, L.; Tan, R.X. Terpenoids and flavonoids from Artemisia species. Planta Medica 2000, 66, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Turi, C.E.; Shipley, P.R.; Murch, S.J. North American Artemisia species from the subgenus Tridentatae (Sagebrush): A phytochemical, botanical and pharmacological review. Phytochemistry 2014, 98, 9–26. [Google Scholar] [CrossRef]
- Taleghani, A.; Emami, S.A.; Tayarani-Najaran, Z. Artemisia: A promising plant for the treatment of cancer. Bioorganic Med. Chem. 2020, 28, 115180. [Google Scholar] [CrossRef]
- Roos, W.P.; Kaina, B. DNA damage-induced cell death by apoptosis. Trends Mol. Med. 2006, 12, 440–450. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, X.; Tang, M.; Li, L.; Lei, Y.; Cheng, P.; Guo, W.; Zheng, Y.; Wang, W.; Luo, N.; et al. The role of ROS and subsequent DNA-damage response in PUMA-induced apoptosis of ovarian cancer cells. Oncotarget 2017, 8, 23492–23506. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Kim, N.H.; Paul, S.; Jakhar, R.; Han, J.; Kang, S.C. 5-Hydroxy-7-Methoxyflavone Triggers Mitochondrial-Associated Cell Death via Reactive Oxygen Species Signaling in Human Colon Carcinoma Cells. PLoS ONE 2016, 11, e0154525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Lai, H.; Chen, J.; Li, L.; Wong, Y.S.; Chen, T.; Li, X. Natural borneol, a monoterpenoid compound, potentiates selenocystine-induced apoptosis in human hepatocellular carcinoma cells by enhancement of cellular uptake and activation of ROS-mediated DNA damage. PLoS ONE 2013, 8, e63502. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.L.; Sedlic, F.; Bosnjak, Z.; Nilakantan, V. SOD1 and MitoTEMPO partially prevent mitochondrial permeability transition pore opening, necrosis, and mitochondrial apoptosis after ATP depletion recovery. Free Radic. Biol. Med. 2010, 49, 1550–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Wei, J.; Chen, Y.; He, P.; Lin, J.; Tan, S.; Nie, J.; Lu, S.; He, M.; Lu, Z.; et al. Isoorientin from Gypsophila elegans induces apoptosis in liver cancer cells via mitochondrial-mediated pathway. J. Ethnopharmacol. 2016, 187, 187–194. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, X.; Huang, J.; Dong, Q. Wnt signaling regulation of stem-like properties in human lung adenocarcinoma cell lines. Med. Oncol. 2015, 32, 157. [Google Scholar] [CrossRef]
- Stewart, D.J. Wnt signaling pathway in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Sun, P.L.; Li, J.Z.; Jheon, S.; Lee, C.T.; Chung, J.H. Aberrant Wnt1/β-catenin expression is an independent poor prognostic marker of non-small cell lung cancer after surgery. J. Thorac. Oncol. 2014, 6, 716–724. [Google Scholar] [CrossRef] [Green Version]
- David, M.D.; Cantí, C.; Herreros, J. Wnt-3a and Wnt-3 differently stimulate proliferation and neurogenesis of spinal neural precursors and promote neurite outgrowth by canonical signaling. J. Neurosci. Res. 2010, 88, 3011–3023. [Google Scholar] [CrossRef] [Green Version]
- Okoye, U.C.; Malbon, C.C.; Wang, H.Y. Wnt and Frizzled RNA expression in human mesenchymal and embryonic (H7) stem cells. J. Mol. Signal. 2008, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Shi, J.; Huang, Y.; Lai, L. Expression of canonical WNT/β-CATENIN signaling components in the developing human lung. BMC Dev. Biol. 2012, 12, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Sequence | Reverse Sequence |
|---|---|---|
| GAPDH | GAAATCCCATCACCATCTTCCAGG | GAGCCCCAGCCTTCTCCATG |
| Bax | GCCCTTTTGCTTCAGGGTTT | TCCAATGTCCAGCCCATGAT |
| Bad | CCTCAGGCCTATGCAAAAAG | AAACCCAAAACTTCCGATGG |
| Bcl2 | GATTGTGGCCTTCTTTGAG | CAAACTGAGCAGAGTCTTC |
| Mcl-1 | GGACATCAAAAACGAAGACG | GCAGCTTTCTTGGTTTATGG |
| Cyclin D1 | CCGTCCATGCGGAAGATC | GAAGACCTCCTCCTCGCACT |
| c-myc | AGCGACTCTGAGGAGGAACAAG | GTGGCACCTCTTGAGGACCA |
| Wnt3 | CGCTCAGCTATGAACAAGCA | AAAGTTGGGGGAGTTCTCGT |
| β-catenin | TCTGAGGACAAGCCACAAGATTACA | TGGGCACCAATATCAAGTCCAA |
| Bcl-XL | CAGAGCTTTGAACAGGTAG | GCTCTCGGGTGCTGTATTG |
| Peak No. | Retention Time | Area | Area (%) | Compound Identified | Molecular Formula |
|---|---|---|---|---|---|
| 1. | 4.464 | 71,441 | 0.83 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | C6H8O4 |
| 2. | 6.718 | 379,461 | 4.43 | 2,3,6,10,10-Pentamethyl-1-oxa-spiro(4,5)deca-3 | C14H22O |
| 3. | 7.925 | 59,506 | 0.69 | 3-(Pyrrolidin-1-yl)cyclopent-2-en-1-one | C9H13NO |
| 4. | 8.382 | 561,175 | 6.55 | 2H-1-benzopyran-2-one | C9H6O2 |
| 5. | 8.591 | 546,765 | 6.38 | Guanosine | C10H13N5O5 |
| 6. | 9.000 | 41,899 | 0.49 | 4H-1,2,3,6,7,8,9,9A-octahydroquinolizine-1,4,8-D | C9H12D3N |
| 7. | 9.086 | 27,165 | 0.32 | 2,4-Ditert-butylphenyl 5-hydroxypentanoate | C19H30O3 |
| 8. | 9.241 | 104,843 | 1.22 | Decanoic anhydride | C20H38O3 |
| 9. | 10.063 | 128,609 | 1.50 | 1-(4-Methylphenyl)-1H-pyrrole | C11H11N |
| 10. | 10.942 | 1,664,345 | 19.42 | 1,3,4,5-Tetrahydroxycyclohexanecarboxylic acid | C7H12O6 |
| 11. | 11.269 | 124,502 | 1.45 | 3alpha,7beta-Dihydroxy-5beta,6beta-epoxycholestane | C27H46O3 |
| 12. | 11.356 | 40,194 | 0.47 | 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine | C12H15N |
| 13. | 11.992 | 188,369 | 2.20 | Nonylamine, N,N-di(allyl) | C15H29N |
| 14. | 12.207 | 44,211 | 0.52 | (1R,3E,7E,11R)-1,5,5,8-Tetramethyl-12-oxabicyclo [9.1.0]dodeca-3,7-diene | C7H12O6 |
| 15. | 12.299 | 40,009 | 0.47 | 6-Hydroxy-4,4,7a-trimethyl-5,6,7,7a-tetrahydrobenzofuran-2(4H)- | C11H16O3 |
| 16. | 12.625 | 246,632 | 2.88 | Arctiol | C15H26O2 |
| 17. | 12.721 | 96,591 | 1.13 | Neophytadiene | C20H38 |
| 18. | 12.981 | 24,151 | 0.28 | 10,12-Tetradecadienal; Vernaldehyde | C14H24O |
| 19. | 13.074 | 45,010 | 0.53 | 2-Furanmethanol, tetrahydro-;Dihydropanaxacol | C17H28O3 |
| 20. | 13.168 | 23,344 | 0.27 | 3,7,11,15-Tetramethylhexadec-2-en-1-yl acetate; Cetoleic acid | C2 O22H42 |
| 21. | 13.414 | 123,198 | 1.44 | 2-(Dimethylamino)-N-(2,6-dimethylphenyl) acetamide; Lidocaine | C14H22N2O |
| 22. | 13.563 | 51,472 | 0.60 | Limonene dioxide; Diosphenol | C10H16O2 |
| 23. | 13.780 | 116,889 | 1.36 | 1H-Cyclopropa[a]naphthalene, 1a,2,3,5,6,7a,7b-octahydro-1,1,7,7a-tetramethyl-, 1aR-1aα,7α,7aα,7bβ | C15H24 |
| 24. | 13.995 | 239,211 | 2.79 | 5,10-Diethoxy-2,3,7,8-tetrahydro-1H,6H-dipyrrolo[1,2-a:1’,2’-d]pyrazine; Rivastigmine | C14H22N2O2 |
| 25. | 14.355 | 233,920 | 2.73 | Scopoletin | C10H8O4 |
| 26. | 14.840 | 124,027 | 1.45 | Martynoside | C31H40O15 |
| 27. | 14.980 | 29,151 | 0.34 | Geranic acid | C10H16O2 |
| 28. | 15.042 | 224,828 | 2.62 | Deoxyartemisinin | C15H22O4 |
| 29. | 15.222 | 111,627 | 1.30 | Retinal | C20H28O |
| 30. | 15.341 | 88,859 | 1.04 | 5-oxatricyclo[4.4.0.01,4]decan-7-one; 4-Heptyloxyphenol | C13H20O2 |
| 31. | 15.441 | 239,833 | 2.80 | Phytol isomer | C20H40O |
| 32. | 15.749 | 182,147 | 2.13 | Farnesol | C15H26O |
| 33. | 15.849 | 55,741 | 0.65 | Cycloionone | C13H20O |
| 34. | 16.057 | 58,278 | 0.68 | 1H-2-Indenone,2,4,5,6,7,7a-hexahydro-3-(1-methylethyl) | |
| 35. | 16.339 | 25,000 | 0.29 | 1,8-Diazabicyclo[5.4.0]undec-7-en-11-one; Phenoxypropazine | C9H14N2O |
| 36. | 16.591 | 277,133 | 3.23 | Cyperadione; Farnesoic acid | C15H24O2 |
| 37. | 16.956 | 22,939 | 0.27 | 2-dimethylaminoethyl ester; Hexanoic acid | C10H21NO2 |
| 38. | 18.447 | 64,492 | 0.75 | N-cis-11-eicosaenoylethanolamine; 2-(Dimethylamino)ethyl vaccenoate | C22H43NO2 |
| 39. | 19.034 | 48,587 | 0.57 | Phthalic acid | C24H38O4 |
| 40. | 20.269 | 119,663 | 1.40 | 1-Hexacosanol | C26H54O |
| 41. | 21.024 | 1,122,793 | 13.10 | Erucamide; 13-Docosenamide | C22H43NO |
| 42. | 22.127 | 79,068 | 0.92 | Chloroacetic acid | C14H27ClO2 |
| 43. | 22.726 | 42,958 | 0.50 | Octadecamethylcyclononasiloxane | C18H54O9Si9 |
| 44. | 25.217 | 37,974 | 0.44 | Vitamin E | C29H50O2 |
| 46. | 25.760 | 92,327 | 1.08 | 4-(acetyloxy)-1-(1,5-dimethylhexyl)-3A,6,6,12A-tetramethyl-2,3,3A,3B,5A,6,7,8,9,11,12,12A-dodecahydro-1H-cyclopenta[A]cyclopropa[E]phenanthren-7-yl acetate | C34H53DO4 |
| 47. | 27.639 | 102,832 | 1.20 | Stigmasterone | C29H46O |
| 48. | 28.962 | 77,279 | 0.90 | Stearoyldelicone | C33H54O3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, R.K.; Ahmad, A.; Khan, A.F.; Al-Keridis, L.A.; Saeed, M.; Alshammari, N.; Alabdallah, N.M.; Ansari, I.A.; Mujeeb, F. Ethanolic Extract of Artemisia vulgaris Leaf Promotes Apoptotic Cell Death in Non-Small-Cell Lung Carcinoma A549 Cells through Inhibition of the Wnt Signaling Pathway. Metabolites 2023, 13, 480. https://doi.org/10.3390/metabo13040480
Tiwari RK, Ahmad A, Khan AF, Al-Keridis LA, Saeed M, Alshammari N, Alabdallah NM, Ansari IA, Mujeeb F. Ethanolic Extract of Artemisia vulgaris Leaf Promotes Apoptotic Cell Death in Non-Small-Cell Lung Carcinoma A549 Cells through Inhibition of the Wnt Signaling Pathway. Metabolites. 2023; 13(4):480. https://doi.org/10.3390/metabo13040480
Chicago/Turabian StyleTiwari, Rohit Kumar, Afza Ahmad, Ahamad Faiz Khan, Lamya Ahmed Al-Keridis, Mohd Saeed, Nawaf Alshammari, Nadiyah M. Alabdallah, Irfan Ahmad Ansari, and Farina Mujeeb. 2023. "Ethanolic Extract of Artemisia vulgaris Leaf Promotes Apoptotic Cell Death in Non-Small-Cell Lung Carcinoma A549 Cells through Inhibition of the Wnt Signaling Pathway" Metabolites 13, no. 4: 480. https://doi.org/10.3390/metabo13040480
APA StyleTiwari, R. K., Ahmad, A., Khan, A. F., Al-Keridis, L. A., Saeed, M., Alshammari, N., Alabdallah, N. M., Ansari, I. A., & Mujeeb, F. (2023). Ethanolic Extract of Artemisia vulgaris Leaf Promotes Apoptotic Cell Death in Non-Small-Cell Lung Carcinoma A549 Cells through Inhibition of the Wnt Signaling Pathway. Metabolites, 13(4), 480. https://doi.org/10.3390/metabo13040480