



Secondary Metabolites Profiling, Antimicrobial and Cytotoxic Properties of Commiphora gileadensis L. Leaves, Seeds, Callus, and Cell Suspension Extracts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Callus Culture Establishment and Maintenance
2.3. Cell Suspension Establishment and Maintenance
2.4. Preparation of Methanolic Extracts
2.5. Chemical Constituents and Molecular Networking Analysis
2.6. Antimicrobial Assays
2.7. Cytotoxicity Assays
3. Results and Discussion
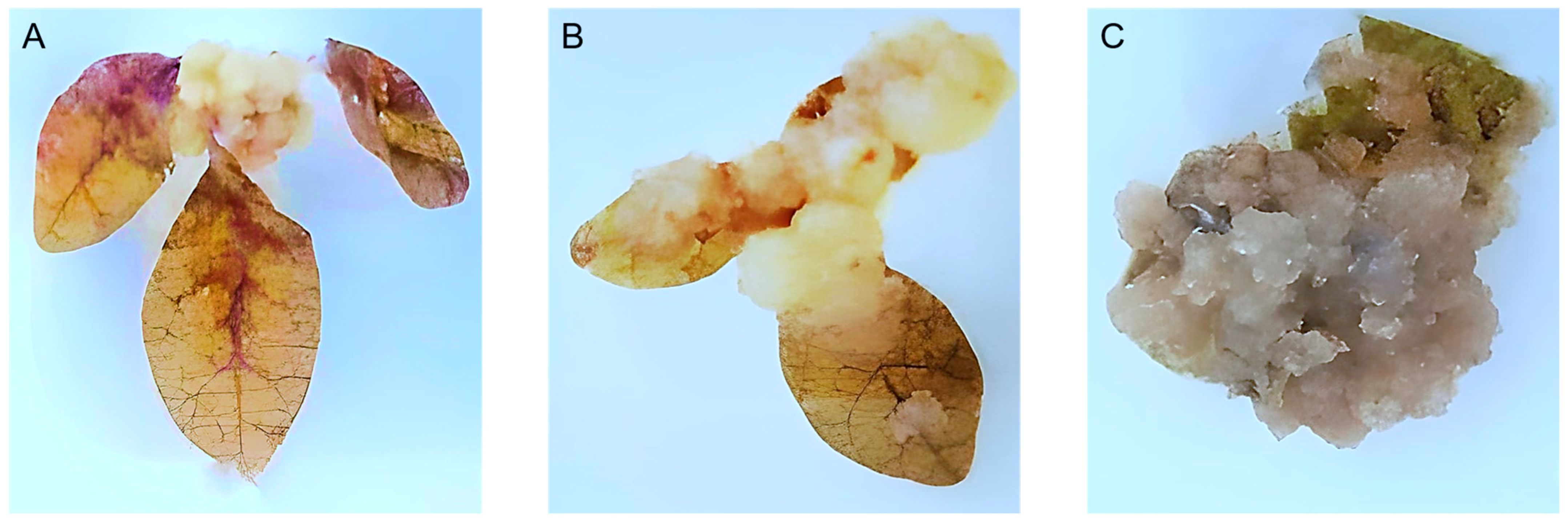
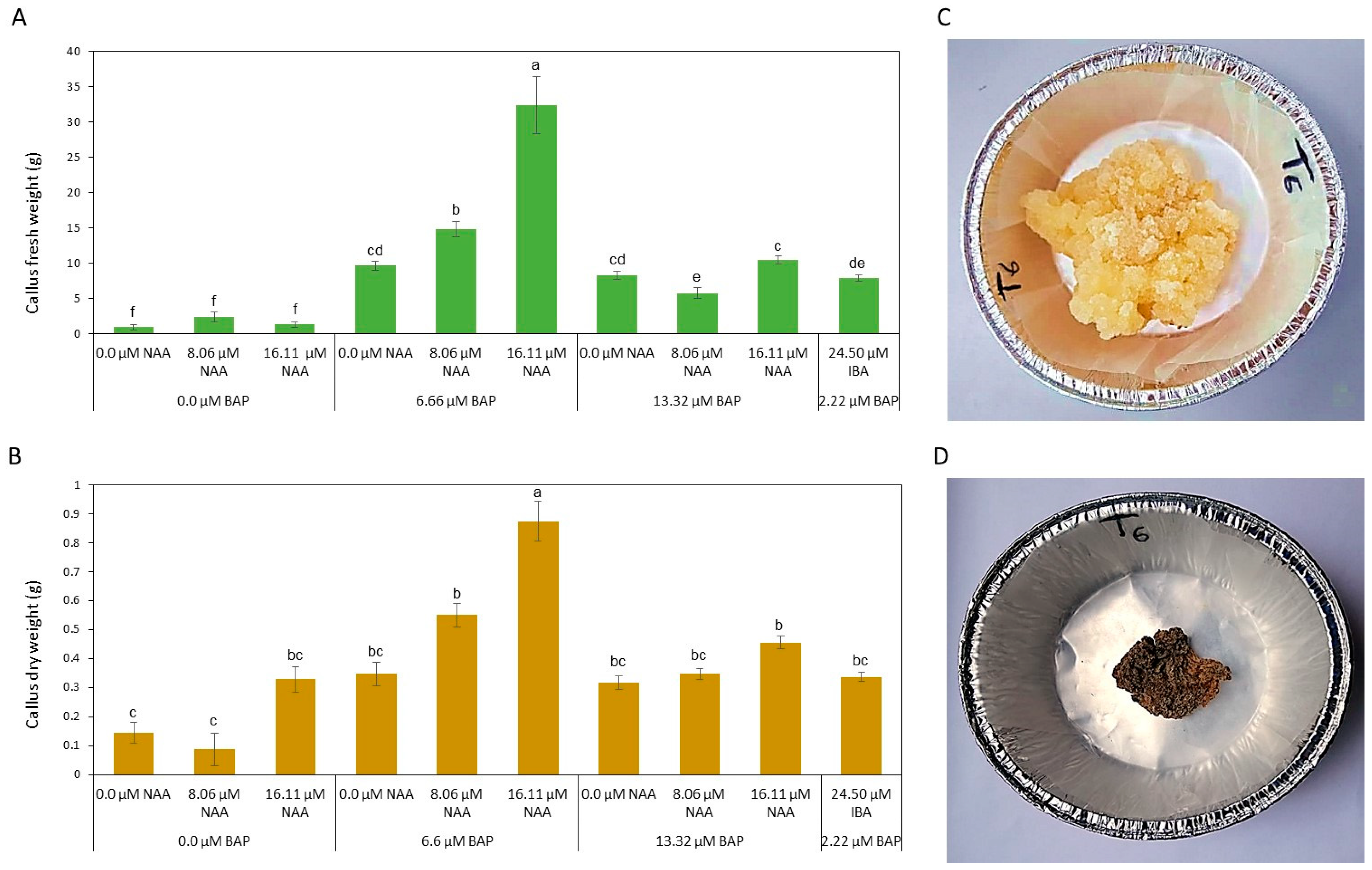
3.1. C. gileadensis Callus Culture
3.2. C. gileadensis Cell Suspension Culture
3.3. Antimicrobial Effects of C. gileadensis Methanolic Extract
3.4. Cytotoxic Effects of C. gileadensis Methanolic Extract
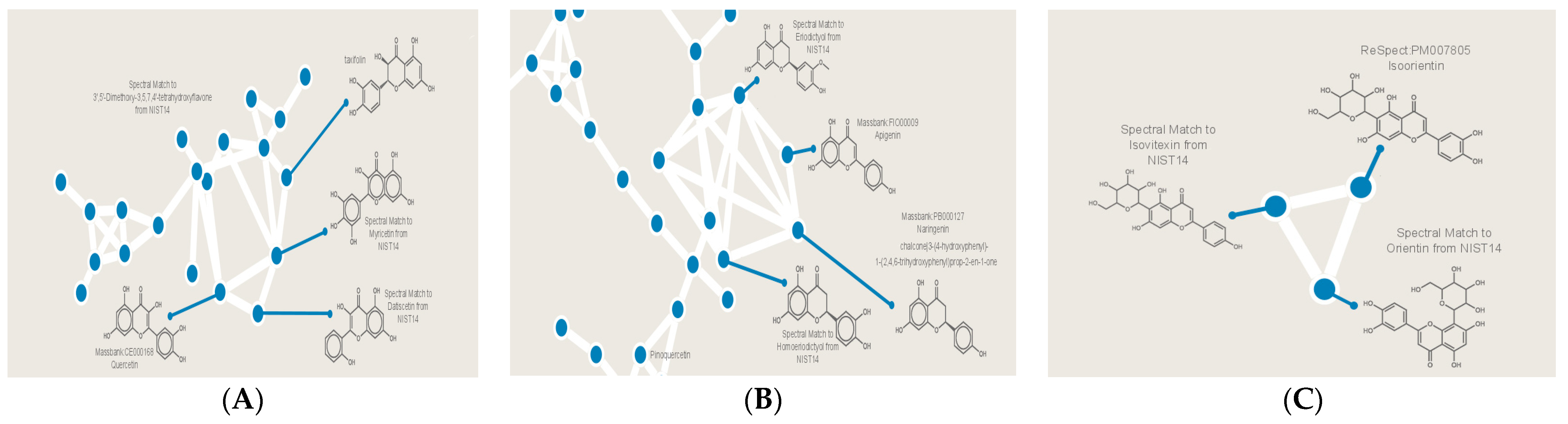
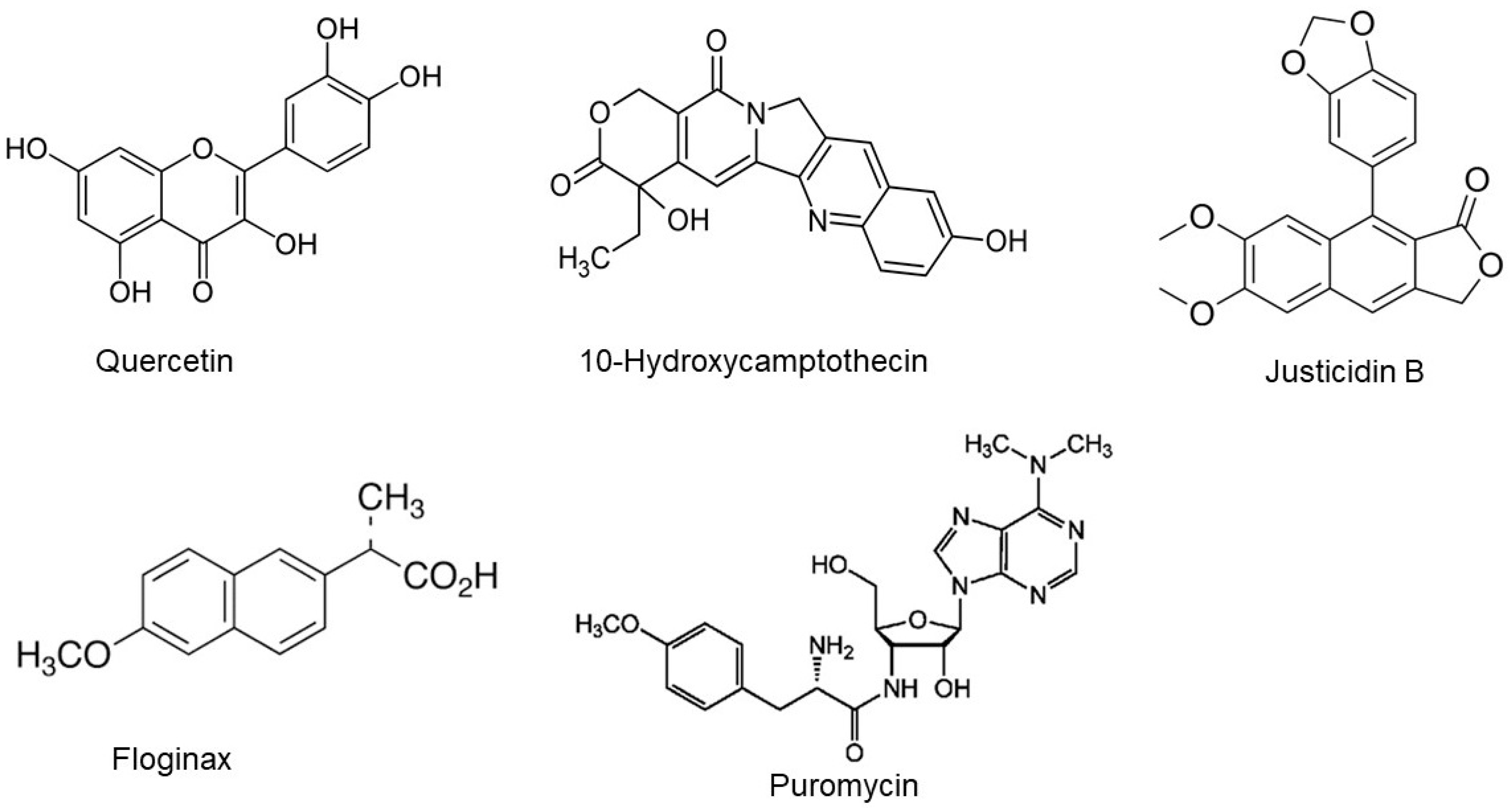
3.5. Secondary Metabolites Profiling of C. gileadensis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shalabi, L.F.; Otaif, F.S. Commiphora Jacq (Burseraceae) in Saudi Arabia, Botanical, Phytochemical and Ethnobotanical Notes. Ecologies 2022, 3, 38–57. [Google Scholar] [CrossRef]
- Yaniv, Z.; Dudai, N. Medicinal and Aromatic Plants of the Middle-East; Springer: Berlin/Heidelberg, Germany, 2014; Volume 2, pp. 67–150. [Google Scholar]
- Hepper, N.F.; Taylor, J.E. Date Palms and Opobalsam in the Madaba Mosaic Map. Palest. Explor. Q. 2004, 136, 35–44. [Google Scholar] [CrossRef]
- Eslamieh, J. Commiphora gileadensis. Cactus Succul. J. 2011, 83, 206–210. [Google Scholar] [CrossRef]
- Dudai, N.; Shachter, A.; Satyal, P.; Setzer, W. Chemical Composition and Monoterpenoid Enantiomeric Distribution of the Essential Oils from Apharsemon (Commiphora gileadensis). Medicines 2017, 4, 66. [Google Scholar] [CrossRef]
- Iluz, D.; Hoffman, M.; Gilboa-Garber, N.; Amar, Z. Medicinal Properties of Commiphora gileadensis. Afr. J. Pharm. Pharmacol. 2010, 4, 516–520. [Google Scholar]
- Shen, T.; Li, G.H.; Wang, X.-N.; Lou, H.-X. The Genus Commiphora: A Review of Its Traditional Uses, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2012, 142, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Amiel, E.; Ofir, R.; Dudai, N.; Soloway, E.; Rabinsky, T.; Rachmilevitch, S. β-Caryophyllene, a Compound Isolated from the Biblical Balm of Gilead (Commiphora gileadensis), Is a Selective Apoptosis Inducer for Tumor Cell Lines. Evid. Based Complement. Alternat. Med. 2012, e872394. [Google Scholar] [CrossRef] [PubMed]
- Alhazmi, A.; Aldairi, A.F.; Alghamdi, A.; Alomery, A.; Mujalli, A.; Obaid, A.A.; Farrash, W.F.; Allahyani, M.; Halawani, I.; Aljuaid, A.; et al. Antibacterial Effects of Commiphora gileadensis Methanolic Extract on Wound Healing. Molecules 2022, 27, 3320. [Google Scholar] [CrossRef]
- Al-Zahrani, S.A.; Bhat, R.S.; Al-Onazi, M.A.; Alwhibi, M.S.; Soliman, D.A.; Aljebrin, N.A.; Al-Suhaibani, L.S.; Al Daihan, S. Anticancer potential of biogenic silver nanoparticles using the stem extract of Commiphora gileadensis against human colon cancer cells. Green Process. Synth. 2022, 11, 435–444. [Google Scholar] [CrossRef]
- Wineman, E.; Douglas, I.; Wineman, V.; Sharova, K.; Jaspars, M.; Meshner, S.; Bentwich, Z.; Cohen, G.; Shtevi, A. Commiphora gileadensis Sap Extract Induces Cell Cycle-Dependent Death in Immortalized Keratinocytes and Human Dermoid Carcinoma Cells. J. Herb. Med. 2015, 5, 199–206. [Google Scholar] [CrossRef]
- Althurwi, H.N.; Salkini, M.A.A.; Soliman, G.A.; Ansari, M.N.; Ibnouf, E.O.; Abdel-Kader, M.S. Wound Healing Potential of Commiphora gileadensis Stems Essential Oil and Chloroform Extract. Separations 2022, 9, 254. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Mohamed, G.A.; Ibrahim, S.R.M.; Koshak, A.E.; Alnashri, I.; Alghamdi, A.; Aljohani, A.; Khairy, A. Commigileadin A: A new triterpenoid from Commiphora gileadensis aerial parts. Pharmacogn. Mag. 2022, 18, 256. [Google Scholar]
- Farid, M.M.; Aboul Naser, A.F.; Salem, M.M.; Ahmed, Y.R.; Emam, M.; Hamed, M.A. Chemical Compositions of Commiphora opobalsamum Stem Bark to Alleviate Liver Complications in Streptozotocin-Induced Diabetes in Rats: Role of Oxidative Stress and DNA Damage. Biomarkers 2022, 27, 671–683. [Google Scholar] [CrossRef]
- Alsherif, E.A. Ecological Studies of Commiphora Genus (Myrrha) in Makkah Region, Saudi Arabia. Heliyon 2019, 5, e01615. [Google Scholar] [CrossRef]
- Twaij, B.M.; Hasan, M.N. Bioactive Secondary Metabolites from Plant Sources: Types, Synthesis, and Their Therapeutic Uses. Int. J. Plant Biol. 2022, 13, 4–14. [Google Scholar] [CrossRef]
- Bouque, V.; Bourgaud, F.; Nguyen, C.; Guckert, A. Production of Daidzein by Callus Cultures of Psoralea species and Comparison with plants. Plant Cell Tissue Organ Cult. 1998, 53, 35–40. [Google Scholar] [CrossRef]
- Tanwar, Y.S.; Mathur, M.; Ramawat, K.G. Morphactin Influences Guggulsterone Production in Callus Cultures of Commiphora wightii. Plant Growth Regul. 2006, 51, 93–98. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Plant Physiol. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Kleigrewe, K.; Almaliti, J.; Tian, I.Y.; Kinnel, R.B.; Korobeynikov, A.; Monroe, E.A.; Duggan, B.M.; Di Marzo, V.; Sherman, D.H.; Dorrestein, P.C.; et al. Combining Mass Spectrometric Metabolic Profiling with Genomic Analysis: A Powerful Approach for Discovering Natural Products from Cyanobacteria. J. Nat. Prod. 2015, 78, 1671–1682. [Google Scholar] [CrossRef]
- Karlsmose, S. Laboratory Protocol: “Susceptibility Testing of Enterobacteriaceae Using Disk Diffusion”. WHO Global Foodborne Infections Network. Protocol 2010, GFNLAB002. Available online: https://antimicrobialresistance.dk/CustomerData/Files/Folders/6-pdf-protocols/57_sop-dd-2010gfnlab002-v2.pdf (accessed on 2 November 2022).
- Motawi, T.; Bustanji, Y.; El-Maraghy, S.; Taha, M.; Al-Ghussein, M. Evaluation of naproxen and cromolyn activities against cancer cells viability, proliferation, apoptosis, p53 and gene expression of survivin and caspase-3. J. Enzyme Inhib. Med. Chem. 2014, 29, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Abu-Rish, E.Y.; Kasabri, V.; Hudaib, M.M.; Mashalla, S.H.; AlAlawi, L.H.; Tawaha, K.; Mohammad, M.K.; Mohamed, Y.S.; Bustanji, Y. Evaluation of Antiproliferative Activity of Some Traditional Anticancer Herbal Remedies from Jordan. Trop. J. Pharm. Res. 2016, 15, 469–474. [Google Scholar] [CrossRef]
- Nikam, T.D.; Ghorpade, R.P.; Nitnaware, K.M.; Ahire, M.L.; Lokhande, V.H.; Chopra, A. Micropropagation and Non-Steroidal Anti-Inflammatory and Anti-Arthritic Agent Boswellic Acid Production in Callus Cultures of Boswellia serrata Roxb. Physiol. Mol. Biol. Plants 2013, 19, 105–116. [Google Scholar] [CrossRef]
- Mishra, S.K.; Kumar, A. Biosynthesis of Guggulsterone in the Callus Culture of Commiphora wightii Arnott. Bhandari (Burseraceae). Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 35–41. [Google Scholar]
- Al Abdallat, A.M.; Sawwan, J.S.; Al Zoubi, B. Agrobacterium tumefaciens-Mediated Transformation of Callus Cells of Crataegus aronia. Plant Cell Tissue Organ Cult. 2010, 104, 31–39. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, X.; Wu, K.; Teixeira da Silva, J.A.; Zeng, S.; Ma, G. Shoot Organogenesis and Plant Regeneration in Cordia Subcordata Lam. In Vitro Cell. Dev. Biol. 2021, 58, 392–398. [Google Scholar] [CrossRef]
- Chatterjee, M.; Chatterjee, S.; Chandra, I. In Vitro Regeneration of Piper longum L. and Comparative RP-HPLC Analysis of Piperine Production of In Vitro and In Vivo Grown Plants. Plant Cell Tissue Organ Cult. 2022, 149, 205–212. [Google Scholar] [CrossRef]
- Daffalla, H.M.; Elsheikh, A.M.; Ali, H.A.; Khalafalla, M.M. Callus Maintenance and Cell Line Selection of Grewia tenax. J. Herbs Spices Med. Plants 2019, 25, 218–235. [Google Scholar] [CrossRef]
- Mathur, M.; Ramawat, K.G. Guggulsterone Production in Cell Suspension Cultures of the Guggul Tree, Commiphora wightii, Grown in Shake-Flasks and Bioreactors. Biotech. Lett. 2007, 29, 979–982. [Google Scholar] [CrossRef]
- Navinraj, S.; Santhanakrishnan, V.P.; Boopathi, N.M.; Balasubramani, V.; Raghu, R.; Rajamani, K.; Vellaikumar, S. Establishment of Cell Suspension Cultures of Azadirachta indica for the Production of Nimbolide. Med. Plants-Int. J. Phytomed. Relat. Ind. 2014, 14, 105–112. [Google Scholar] [CrossRef]
- Mathur, M.; Ramawat, K.G. Improved Guggulsterone Production from Sugars, Precursors, and Morphactin in Cell Cultures of Commiphora Wightii Grown in Shake Flasks and a Bioreactor. Plant Biotechnol. Rep. 2008, 2, 133–136. [Google Scholar] [CrossRef]
- Ozawa, K.; Komamine, A. Establishment of a System of High-Frequency Embryogenesis from Long-Term Cell Suspension Cultures of Rice (Oryza sativa L.). Theor. Appl. Genet. 1989, 77, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.K.; Lu, C.W.; Chang, J.Y.; Lu, C.F.; Tan, C.C.; Huang, L.F. Optimization of the culture medium for recombinant protein production under the control of the αAmy3 promoter in a rice suspension-cultured cell expression system. Plant Cell Tissue Organ Cult. 2018, 132, 383–391. [Google Scholar] [CrossRef]
- Zhu, S.-S.; Qin, D.-P.; Wang, S.-X.; Yang, C.; Li, G.-P.; Cheng, Y.-X. Commipholactam A, a Cytotoxic Sesquiterpenoidal Lactam from Resina commiphora. Fitoterapia 2019, 134, 382–388. [Google Scholar] [CrossRef]
- Dinku, W.; Isaksson, J.; Rylandsholm, F.G.; Bouř, P.; Brichtová, E.; Choi, S.U.; Lee, S.-H.; Jung, Y.-S.; No, Z.S.; Svendsen, J.S.M.; et al. Anti-Proliferative Activity of a Novel Tricyclic Triterpenoid Acid from Commiphora africana Resin against Four Human Cancer Cell Lines. Appl. Biol. Chem. 2020, 63, 16. [Google Scholar] [CrossRef]
- Bustanji, Y.; Taha, M.O.; Yousef, A.-M.; Al-Bakri, A.G. Berberine Potently Inhibits Protein Tyrosine Phosphatase 1B: Investigation by Docking Simulation and Experimental Validation. J. Enzyme Inhib. Med. Chem. 2006, 21, 163–171. [Google Scholar] [CrossRef]
- Assaf, A.M.; Haddadin, R.N.; Aldouri, N.A.; Alabbassi, R.; Mashallah, S.; Mohammad, M.; Bustanji, Y. Anti-Cancer, Anti-Inflammatory and Anti-Microbial Activities of Plant Extracts Used against Hematological Tumors in Traditional Medicine of Jordan. J. Ethnopharmacol. 2013, 145, 728–736. [Google Scholar] [CrossRef]
- Ghorbani, S.; Kosari-Nasab, M.; Mahjouri, S.; Talebpour, A.H.; Movafeghi, A.; Maggi, F. Enhancement of in vitro production of volatile organic compounds by shoot differentiation in Artemisia spicigera. Plants 2021, 10, 208. [Google Scholar] [CrossRef]
- Abu-Darwish, D.; Shibli, R.; Al-Abdallat, A.M. In Vitro Cultures and Volatile Organic Compound Production in Chiliadenus montanus (Vhal.) Brullo. Plants 2022, 11, 1326. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Callus | Cell Suspension | Leaves | Seeds | Tetracycline | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Microbial Species | IZT y | MIC Z | IZT | MIC | IZT | MIC | IZT | MIC | IZT | MIC |
| B. subtilis | - | - | - | - | - | 6.25 | - | - | 18.0 ± 0.20 | 1.56 |
| E. coli | - | - | - | - | - | - | - | - | 10.06 ± 0.17 | 0.39 |
| S. aureus | - | - | - | 6.25 * | 2.50 ± 0.07 ** | 6.25 | 2.30 ± 0.17 | 1.56 | 17.00 ± 0.10 | 0.78 |
| S. epidermidis | - | - | - | 6.25 | 1.50 ± 0.09 | 3.12 | - | 3.12 | 14.00 ± 0.84 | 1.56 |
| Salmonella spp. | - | - | - | - | 1.70 ± 0.14 | 12.50 | - | 12.50 | 22.20 ± 0.66 | 0.78 |
| E. carotovora | - | - | - | - | - | - | - | - | 18.00 ± 0.20 | 1.56 |
| K. pneumoniae | - | - | - | - | - | - | - | - | 17.00 ± 0.10 | 1.56 |
| C. albicans | - | - | - | - | - | 6.25 | - | - | 18.00 ± 0.00 | 6.25 |
| Cell Line | Callus | Cell Suspension | Leaves | Seeds | Doxorubicin |
|---|---|---|---|---|---|
| PanC1 | 488.20 * ± 25.40 ** | NA | 13.60 ± 1.20 | 106.6 ± 6.7 | 1.30 ± 0.09 |
| A549 | 117.50 ± 3.50 | 137.40 ± 9.90 | 1.00 ± 0.50 | 148.7 ± 12.4 | 3.50 ± 0.03 |
| MCF7 | NA *** | NA | 34.20 ± 2.80 | 350.00 ± 32.50 | 5.00 ± 0.79 |
| PC3 | NA | NA | 76.40 ± 4.70 | NA | 1.10 ± 0.05 |
| Fibroblast | Not toxic | Not toxic | 82.50 ± 7.90 | Not toxic | 0.022 ± 0.00 |
| Compound Name | Retention Time | Precursor m/z [M + H]+ | Molecular Formula | Plant Extract |
|---|---|---|---|---|
| Eriodictyol | 14.82 | 287.15 | C15H12O6 | Callus and cell suspension |
| Homoeriodictyol | 13.91 | 303.10 | C16H14O6 | Cell suspension |
| Apigenin | 13.65 | 271.20 | C15H10O5 | Cell suspension |
| Naringenin | 13.55 | 273.12 | C15H12O5 | Cell suspension |
| Pinoquercetin | 5.80 | 317.02 | C16H12O7 | Leaves |
| Laudanosine | 7.81 | 358.13 | C21H27NO4 | Cell suspension, seeds and leaves |
| Catechin Gallate | 12.51 | 443.12 | C22H18O10 | Leaves |
| Isovitexin | 9.52 | 433.10 | C21H20O10 | Cell suspension |
| Isoorientin | 11.42 | 449.08 | C21H20O11 | Cell suspension |
| Orientin | 9.35 | 449.06 | C21H20O11 | Cell suspension |
| Quercetin | 4.96 | 303.05 | C15H10O7 | Leaves |
| 10-Hydroxycamptothecin | 3.74 | 365.15 | C20H16N2O5 | Leaves |
| Justicidin B | 16.91 | 365.15 | C21H16O6 | Callus, cell Suspension and leaves |
| Floginax | 16.15 | 230.96 | C14H14O3 | Callus and leaves |
| Puromycin | 8.32 | 472.17 | C22H29N7O5 | Callus, cell Suspension, seeds and leaves |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Abdallat, A.M.; Adayileh, B.K.; Sawwan, J.S.; Shibli, R.; Al-Qudah, T.S.; Abu-Irmaileh, B.; Albdaiwi, R.N.; Almaliti, J.; Bustanji, Y. Secondary Metabolites Profiling, Antimicrobial and Cytotoxic Properties of Commiphora gileadensis L. Leaves, Seeds, Callus, and Cell Suspension Extracts. Metabolites 2023, 13, 537. https://doi.org/10.3390/metabo13040537
Al-Abdallat AM, Adayileh BK, Sawwan JS, Shibli R, Al-Qudah TS, Abu-Irmaileh B, Albdaiwi RN, Almaliti J, Bustanji Y. Secondary Metabolites Profiling, Antimicrobial and Cytotoxic Properties of Commiphora gileadensis L. Leaves, Seeds, Callus, and Cell Suspension Extracts. Metabolites. 2023; 13(4):537. https://doi.org/10.3390/metabo13040537
Chicago/Turabian StyleAl-Abdallat, Ayed M., Batool K. Adayileh, Jamal S. Sawwan, Rida Shibli, Tamara S. Al-Qudah, Bashaer Abu-Irmaileh, Randa N. Albdaiwi, Jehad Almaliti, and Yasser Bustanji. 2023. "Secondary Metabolites Profiling, Antimicrobial and Cytotoxic Properties of Commiphora gileadensis L. Leaves, Seeds, Callus, and Cell Suspension Extracts" Metabolites 13, no. 4: 537. https://doi.org/10.3390/metabo13040537
APA StyleAl-Abdallat, A. M., Adayileh, B. K., Sawwan, J. S., Shibli, R., Al-Qudah, T. S., Abu-Irmaileh, B., Albdaiwi, R. N., Almaliti, J., & Bustanji, Y. (2023). Secondary Metabolites Profiling, Antimicrobial and Cytotoxic Properties of Commiphora gileadensis L. Leaves, Seeds, Callus, and Cell Suspension Extracts. Metabolites, 13(4), 537. https://doi.org/10.3390/metabo13040537