



Effect of Grape Seed Proanthocyanidins on Fat Metabolism and Adipocytokines in Obese Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sources
2.2. Experimental Animals, Diets and Design
2.3. Sample Collection and Analyses
2.3.1. Determination of Routine Indices in Rats
2.3.2. Detection of Cytokines and mRNA Expression in Rat Adipose Tissue
2.3.3. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.3.4. Effect of Procyanidins on the Proliferation of 3T3-L1 Preadipocytes
2.3.5. Effect of Proanthocyanidins on Cytokines in Mature Adipocytes
2.3.6. Statistical Analysis
3. Results
3.1. Effect of GSP Supplementation on Rats Weight
3.2. Effect of GSP on Blood Indicators in Rats
3.3. Effect of GSP on Adipose Tissue Cytokine Levels
3.4. Effect of GSP on Cytokine mRNA Expression in Adipose Tissue
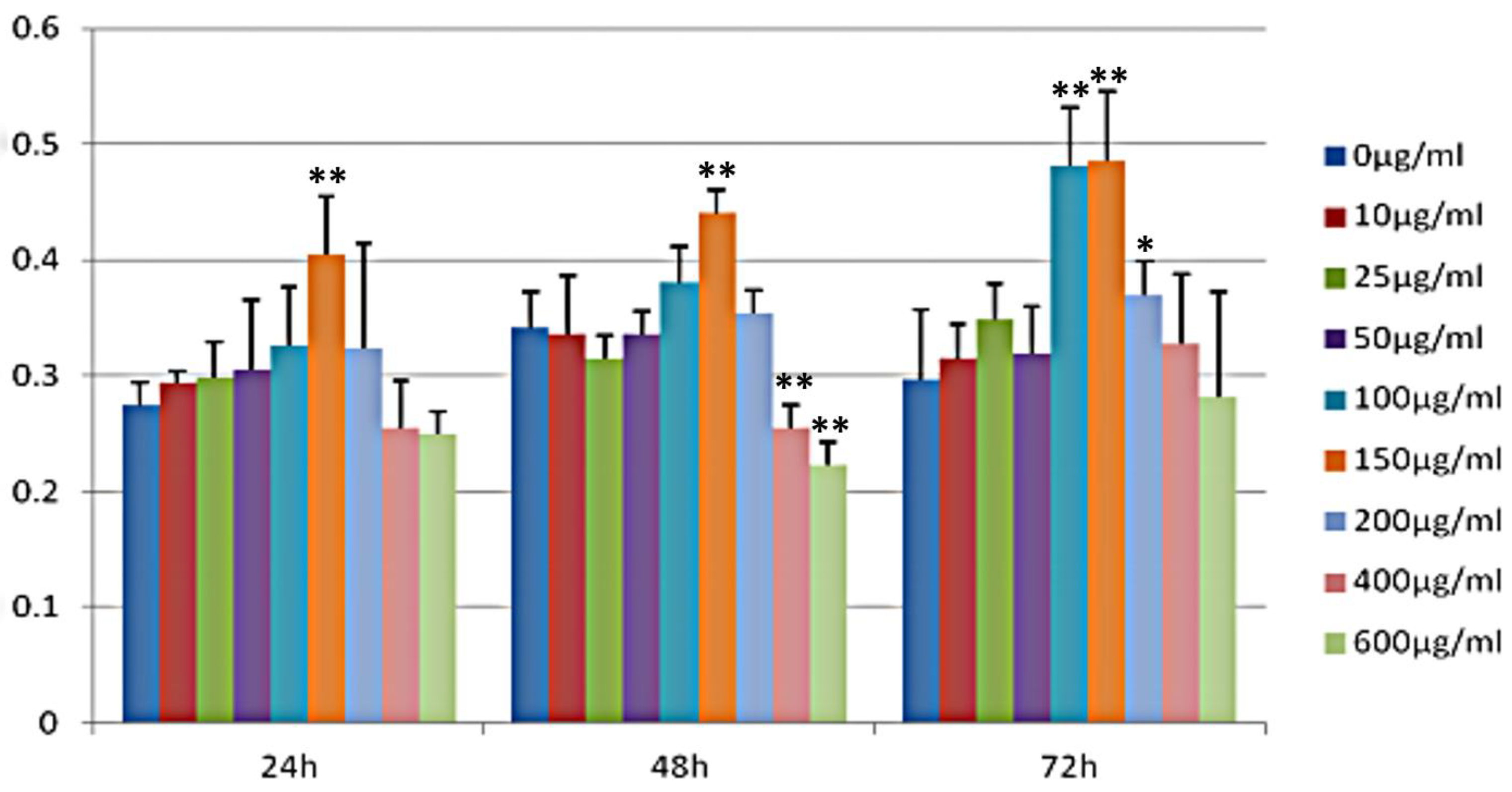
3.5. Effect of GSP on the Proliferation of 3T3-L1 Preadipocytes
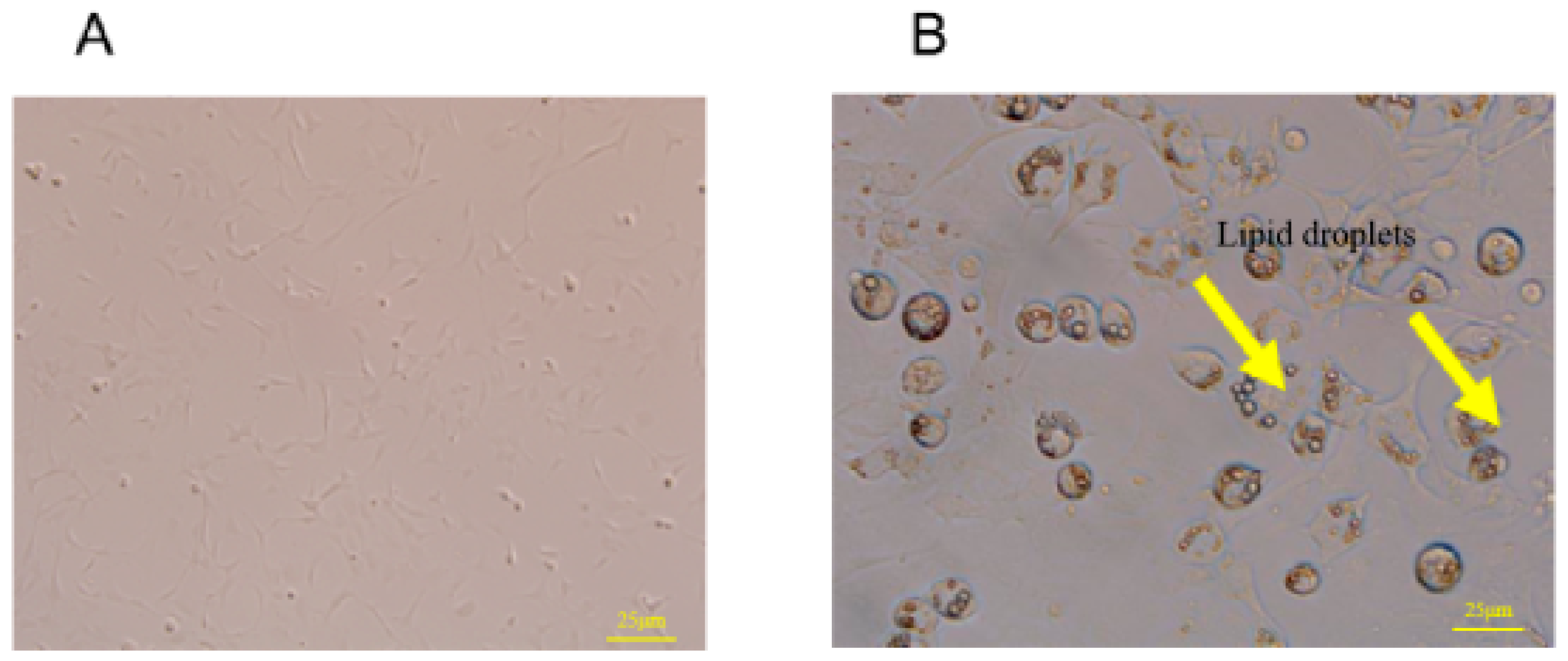
3.6. Morphological Changes of 3T3-L1 Preadipocytes before and after Differentiation
3.7. Morphological Changes in Mature Adipocytes before and after GSP Treatment
3.8. Effect of GSP on Levels of Mature Adipocytokines
3.9. Effect of GSP on mRNA Expression of Mature Adipocyte Factors
4. Discussion
4.1. Body Fat
4.2. Biochemical Indexes
4.3. Adipocytokines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charradi, K.; Mahmoudi, M.; Bedhiafi, T.; Jebari, K.; El May, M.V.; Limam, F.; Aouani, E. Safety evaluation, anti-oxidative and anti-inflammatory effects of subchronically dietary supplemented high dosing grape seed powder (GSP) to healthy rat. Biomed. Pharmacother. 2018, 107, 534–546. [Google Scholar] [CrossRef]
- Gupta, M.; Dey, S.; Marbaniang, D.; Pal, P.; Ray, S.; Mazumder, B. Grape seed extract: Having a potential health benefits. J. Food Sci. Technol. 2020, 57, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.N.; Tonon, R.V.; Mellinger-Silva, C.; Galdeano, M.C.; Iacomini, M.; Santiago, M.C.; Almeida, E.L.; Freitas, S.P. Grape seed pomace as a valuable source of antioxidant fibers. J. Sci. Food Agric. 2019, 99, 4593–4601. [Google Scholar] [CrossRef]
- Sun, H.Y.; Gu, A.X.; Huang, B.Y.; Zhang, T.; Li, J.P.; Shan, A.S. Dietary Grape Seed Proanthocyanidin Alleviates the Liver Injury Induced by Long-Term High-Fat Diets in Sprague Dawley Rats. Front. Vet. Sci. 2022, 9, 959906. [Google Scholar] [CrossRef]
- Piche, M.E.; Tchernof, A.; Despres, J.P. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Nimptsch, K.; Konigorski, S.; Pischon, T. Diagnosis of obesity and use of obesity biomarkers in science and clinical medicine. Metabolism 2019, 92, 61–70. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Kahn, C.R.; Wang, G.; Lee, K.Y. Altered adipose tissue and adipocyte function in the pathogenesis of metabolic syndrome. J. Clin. Investig. 2019, 129, 3990–4000. [Google Scholar] [CrossRef]
- Colom-Pellicer, M.; Rodriguez, R.M.; Navarro-Masip, E.; Bravo, F.I.; Mulero, M.; Arola, L.; Aragones, G. Time-of-day dependent effect of proanthocyanidins on adipose tissue metabolism in rats with diet-induced obesity. Int. J. Obes. 2022, 46, 1394–1402. [Google Scholar] [CrossRef]
- Ferreira, Y.A.M.; Jamar, G.; Estadella, D.; Pisani, L.P. Proanthocyanidins in grape seeds and their role in gut microbiota-white adipose tissue axis. Food Chem. 2023, 404, 134405. [Google Scholar] [CrossRef] [PubMed]
- Gibert-Ramos, A.; Martin-Gonzalez, M.Z.; Crescenti, A.; Salvado, M.J. A Mix of Natural Bioactive Compounds Reduces Fat Accumulation and Modulates Gene Expression in the Adipose Tissue of Obese Rats Fed a Cafeteria Diet. Nutrients 2020, 12, 3251. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Su, L.; Ji, X.; Ma, F.; Yue, Q.; Zhao, C.; Zhang, S.; Sun, X.; Li, K.; Zhao, L. Cistanche promotes the adipogenesis of 3T3-L1 preadipocytes. PLoS ONE 2022, 17, e0264772. [Google Scholar] [CrossRef]
- Yan, H.; Kermouni, A.; Abdel-Hafez, M.; Lau, D.C. Role of cyclooxygenases COX-1 and COX-2 in modulating adipogenesis in 3T3-L1 cells. J. Lipid. Res. 2003, 44, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, K.; Bai, B.R.; Devaraj, S.N. Efficacy of grape seed proanthocyanidins on serum and heart tissue lipids in rats subjected to isoproterenol-induced myocardial injury. Vascul. Pharmacol. 2007, 47, 295–301. [Google Scholar] [CrossRef]
- Liu, M.; Yun, P.; Hu, Y.; Yang, J.; Khadka, R.B.; Peng, X. Effects of Grape Seed Proanthocyanidin Extract on Obesity. Obes. Facts. 2020, 13, 279–291. [Google Scholar] [CrossRef]
- Mildner-Szkudlarz, S.; Bajerska, J. Protective effect of grape by-product-fortified breads against cholesterol/cholic acid diet-induced hypercholesterolaemia in rats. J. Sci. Food Agric. 2013, 93, 3271–3278. [Google Scholar] [CrossRef]
- Song, C.; Zhong, X.; Ge, H.; Liu, M. Research of grape seed powder on serum lipids and hepatic protection effects in high-fat diet rats. J. Hyg. Res. 2013, 42, 991–994. [Google Scholar]
- Guerrero, L.; Margalef, M.; Pons, Z.; Quinones, M.; Arola, L.; Arola-Arnal, A.; Muguerza, B. Serum metabolites of proanthocyanidin-administered rats decrease lipid synthesis in HepG2 cells. J. Nutr. Biochem. 2013, 24, 2092–2099. [Google Scholar] [CrossRef]
- Yin, M.; Zhang, P.; Yu, F.; Zhang, Z.; Cai, Q.; Lu, W.; Li, B.; Qin, W.; Cheng, M.; Wang, H.; et al. Grape seed procyanidin B2 ameliorates hepatic lipid metabolism disorders in db/db mice. Mol. Med. Rep. 2017, 16, 2844–2850. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Jiang, R.; Dong, S.; Xia, L.; Fang, H. Research on Association between Levels of Serum Adiponectin, Hs-CRP, and sICAM-1 and Hypertensive Cerebrovascular Complications. Biomed. Res. Int. 2021, 2021, 4455038. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. Int. J. Mol. Sci. 2020, 21, 2010. [Google Scholar] [CrossRef]
- Satish, M.; Agrawal, D.K. Atherothrombosis and the NLRP3 inflammasome—Endogenous mechanisms of inhibition. Transl. Res. 2020, 215, 75–85. [Google Scholar] [CrossRef]
- Vaid, M.; Singh, T.; Prasad, R.; Kappes, J.C.; Katiyar, S.K. Therapeutic intervention of proanthocyanidins on the migration capacity of melanoma cells is mediated through PGE2 receptors and beta-catenin signaling molecules. Am. J. Cancer Res. 2015, 5, 3325–3338. [Google Scholar]
- Meeran, S.M.; Katiyar, S.K. Proanthocyanidins inhibit mitogenic and survival-signaling in vitro and tumor growth in vivo. Front. Biosci. 2008, 13, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Cline, D.L.; Glavas, M.M.; Covey, S.D.; Kieffer, T.J. Tissue-Specific Effects of Leptin on Glucose and Lipid Metabolism. Endocr. Rev. 2021, 42, 1–28. [Google Scholar] [CrossRef]
- Inoue, T.; Kobayashi, K.; Inoguchi, T.; Sonoda, N.; Fujii, M.; Maeda, Y.; Fujimura, Y.; Miura, D.; Hirano, K.; Takayanagi, R. Reduced expression of adipose triglyceride lipase enhances tumor necrosis factor alpha-induced intercellular adhesion molecule-1 expression in human aortic endothelial cells via protein kinase C-dependent activation of nuclear factor-kappaB. J. Biol. Chem. 2011, 286, 32045–32053. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Wang, T.; Sharma, A.K.; Wolfrum, C. Novel insights into adipose tissue heterogeneity. Rev. Endocr. Metab. Disord. 2022, 23, 5–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 w | 2 w | 4 w | |
|---|---|---|---|
| A | 116.87 ± 7.95 | 183.27 ± 14.91 | 282.32 ± 10.83 b |
| B | 113.87 ± 8.99 | 194.25 ± 8.71 | 299.09 ± 15.98 a |
| C | 111.38 ± 7.81 | 184.52 ± 15.25 | 269.99 ± 11.63 bc |
| D | 109.07 ± 8.29 | 177.72 ± 16.99 | 259.69 ± 13.81 c |
| E | 109.10 ± 6.52 | 180.96 ± 13.41 | 255.44 ± 15.83 c |
| Glu (mmol/L) | TC (mmol/L) | TG (mmol/L) | HDL (mmol/L) | LDL (mmol/L) | |
|---|---|---|---|---|---|
| A | 9.70 ± 0.78 b | 1.43 ± 0.33 b | 0.63 ± 0.17 ab | 165.42 ± 11.26 ab | 55.22 ± 2.75 |
| B | 14.08 ± 3.11 a | 2.27 ± 0.42 a | 0.78 ± 0.11 a | 160.41 ± 3.88 b | 58.17 ± 3.21 |
| C | 10.09 ± 4.14 b | 1.85 ± 0.17 ab | 0.73 ± 0.13 ab | 172.56 ± 6.00 a | 55.37 ± 0.83 |
| D | 9.06 ± 2.37 b | 1.83 ± 0.39 ab | 0.58 ± 0.10 b | 174.89 ± 12.03 a | 57.04 ± 1.70 |
| E | 7.82 ± 3.03 b | 1.75 ± 0.58 b | 0.70 ± 0.12 ab | 176.05 ± 11.45 a | 56.64 ± 2.76 |
| A | B | C | D | E | |
|---|---|---|---|---|---|
| ADPN (μg/L) | 51.35 ± 7.95 | 51.01 ± 8.31 | 55.17 ± 8.77 | 53.96 ± 6.68 | 54.66 ± 9.04 |
| cAMP (nmol/L) | 7.16 ± 0.46 | 7.00 ± 0.66 | 7.01 ± 0.56 | 6.97 ± 0.44 | 6.96 ± 0.33 |
| COX-2 (pg/mL) | 110.77 ± 12.59 ab | 114.45 ± 13.00 a | 106.21 ± 9.71 ab | 98.69 ± 10.77 b | 103.51 ± 12.88 ab |
| IL-6 (ng/L) | 112.45 ± 7.53 ab | 114.84 ± 5.13 a | 106.27 ± 8.96 b | 106.89 ± 6.73 ab | 106.21 ± 5.93 b |
| LEP (μg/L) | 4.53 ± 0.24 | 4.58 ± 0.51 | 4.61 ± 0.24 | 4.41 ± 0.38 | 4.42 ± 0.19 |
| TNF-α (ng/L) | 208.55 ± 5.78 ab | 205.26 ± 5.71 b | 204.73 ± 6.03 b | 214.56 ± 6.40 a | 205.61 ± 5.19 b |
| COX-2 (pg/mL) | LEP (μg/L) | TNF-α (ng/L) | |
|---|---|---|---|
| A | 8.37 ± 1.41 | 5.53 ± 0.57 b | 6.85 ± 0.80 b |
| B | 10.24 ± 1.14 | 7.64 ± 1.01 a | 9.27 ± 0.24 a |
| C | 7.53 ± 0.77 | 7.41 ± 1.05 a | 9.57 ± 0.86 a |
| D | 7.97 ± 0.68 | 5.69 ± 1.05 b | 8.16 ± 0.95 ab |
| E | 8.27 ± 1.18 | 4.93 ± 1.04 b | 6.66 ± 0.52 b |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| ADP (μg/L) | 0 | 35.54 ± 3.54 b | 36.76 ± 3.81 | 29.24 ± 3.80 |
| 100 | 42.40 ± 2.30 ab | 36.85 ± 5.02 | 24.09 ± 3.72 | |
| 150 | 47.87 ± 3.44 a | 39.80 ± 4.10 | 29.72 ± 4.41 | |
| 200 | 50.73 ± 5.04 a | 42.79 ± 5.56 | 33.40 ± 3.46 |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| cAMP (nmol/L) | 0 | 2.02 ± 0.22 b | 2.41 ± 0.56 | 2.13 ± 0.05 |
| 100 | 2.20 ± 0.51 ab | 2.29 ± 0.43 | 2.03 ± 0.53 | |
| 150 | 3.06 ± 0.28 a | 2.86 ± 0.33 | 1.81 ± 0.58 | |
| 200 | 3.03 ± 0.17 a | 2.56 ± 0.42 | 1.61 ± 0.56 |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| COX-2 (pg/mL) | 0 | 1606.46 ± 234.82 b | 1747.08 ± 181.39 | 1479.38 ± 146.28 |
| 100 | 2027.29 ± 145.46 ab | 1850.21 ± 237.05 | 1709.58 ± 282.46 | |
| 150 | 2088.75 ± 256.12 a | 2047.08 ± 310.50 | 1593.96 ± 174.67 | |
| 200 | 1970.00 ± 224.67 ab | 1842.92 ± 145.11 | 1682.50 ± 162.65 |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| IL-6 (ng/L) | 0 | 49.11 ± 8.75 | 42.75 ± 3.19 | 40.50 ± 4.44 |
| 100 | 43.03 ± 1.58 | 38.17 ± 4.33 | 38.88 ± 0.95 | |
| 150 | 52.78 ± 7.84 | 51.44 ± 7.28 | 47.25 ± 6.38 | |
| 200 | 49.82 ± 2.68 | 42.63 ± 3.62 | 45.00 ± 2.95 |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| LEP (μg/L) | 0 | 4.68 ± 0.87 | 4.59 ± 0.44 | 4.62 ± 0.49 |
| 100 | 4.27 ± 0.26 | 4.31 ± 0.11 | 4.76 ± 0.38 | |
| 150 | 4.69 ± 0.57 | 4.19 ± 0.57 | 5.06 ± 0.57 | |
| 200 | 4.71 ± 0.30 | 4.39 ± 0.35 | 4.81 ± 0.26 |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| TNF-α (ng/L) | 0 | 460.39 ± 46.43 | 445.85 ± 37.39 | 438.29 ± 35.47 |
| 100 | 433.45 ± 32.50 | 438.30 ± 16.37 | 402.44 ± 22.67 | |
| 150 | 483.45 ± 34.49 | 441.01 ± 24.68 | 432.29 ± 23.85 | |
| 200 | 474.34 ± 19.37 | 458.64 ± 6.81 | 442.75 ± 47.96 |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| COX-2 (pg/mL) | 0 | 7.61 ± 1.87 | 6.27 ± 1.57 | 9.54 ± 0.61 |
| 100 | 5.40 ± 1.69 | 8.18 ± 1.59 | 6.64 ± 1.59 * | |
| 150 | 7.19 ± 1.22 | 7.39 ± 1.46 | 5.01 ± 0.04 ** | |
| 200 | 6.40 ± 1.07 | 7.08 ± 1.18 | 2.51 ± 0.43 ** |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| LEP (μg/L) | 0 | 14.47 ± 0.14 | 16.13 ± 3.02 | 11.19 ± 1.43 |
| 100 | 5.96 ± 1.89 ** | 11.08 ± 2.70 * | 9.07 ± 2.41 | |
| 150 | 8.69 ± 0.72 ** | 11.11 ± 3.20 * | 4.12 ± 1.53 * | |
| 200 | 3.34 ± 0.00 ** | 7.90 ± 1.05 ** | 1.96 ± 0.85 ** |
| (μg/mL) | Time | |||
|---|---|---|---|---|
| 12 h | 24 h | 48 h | ||
| TNF-α (ng/L) | 0 | 14.14 ± 0.73 | 16.44 ± 3.08 | 8.17 ± 1.74 |
| 100 | 7.79 ± 1.22 ** | 11.74 ± 2.51 | 5.03 ± 1.77 ** | |
| 150 | 10.11 ± 1.57 ** | 11.65 ± 1.19 | 4.80 ± 1.63 ** | |
| 200 | 6.96 ± 1.03 ** | 8.42 ± 1.13 * | 0.43 ± 0.36 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, P.; Fang, L.; Pan, Y.; Jiang, L. Effect of Grape Seed Proanthocyanidins on Fat Metabolism and Adipocytokines in Obese Rats. Metabolites 2023, 13, 568. https://doi.org/10.3390/metabo13040568
Gao P, Fang L, Pan Y, Jiang L. Effect of Grape Seed Proanthocyanidins on Fat Metabolism and Adipocytokines in Obese Rats. Metabolites. 2023; 13(4):568. https://doi.org/10.3390/metabo13040568
Chicago/Turabian StyleGao, Pengxiang, Luoyun Fang, Yucong Pan, and Linshu Jiang. 2023. "Effect of Grape Seed Proanthocyanidins on Fat Metabolism and Adipocytokines in Obese Rats" Metabolites 13, no. 4: 568. https://doi.org/10.3390/metabo13040568
APA StyleGao, P., Fang, L., Pan, Y., & Jiang, L. (2023). Effect of Grape Seed Proanthocyanidins on Fat Metabolism and Adipocytokines in Obese Rats. Metabolites, 13(4), 568. https://doi.org/10.3390/metabo13040568