

Dietary Coated Sodium Butyrate Ameliorates Hepatic Lipid Accumulation and Inflammation via Enhancing Antioxidative Function in Post-Peaking Laying Hens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animals and Diet
2.2. Sample Collection
2.3. Assessment of Liver Injury
2.4. Liver Lipid Profile
2.5. Determination of Antioxidant Capacity
2.6. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.7. Statistical Analysis
3. Results
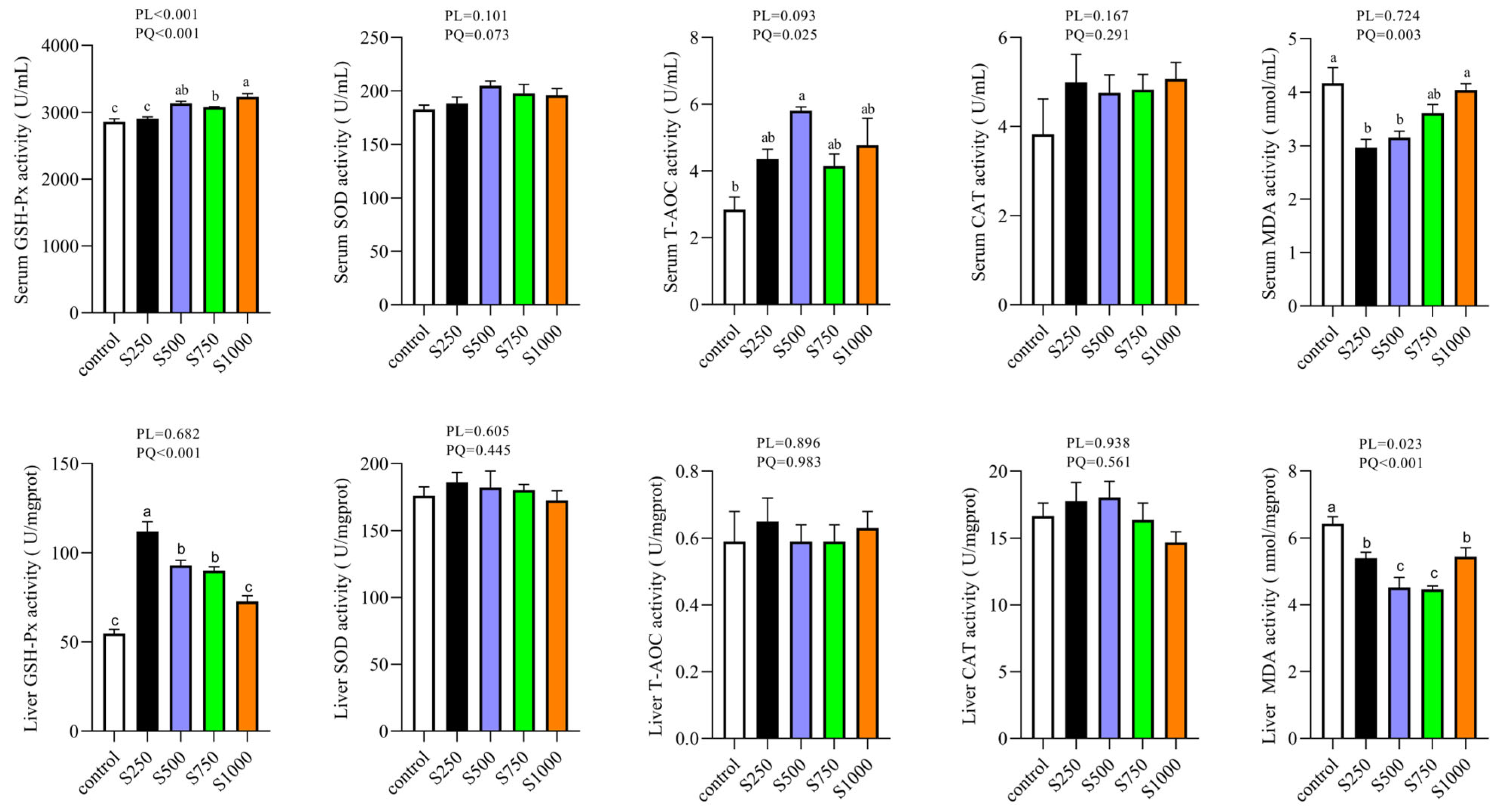
3.1. CSB Enhanced Serum and Hepatic Antioxidant Properties
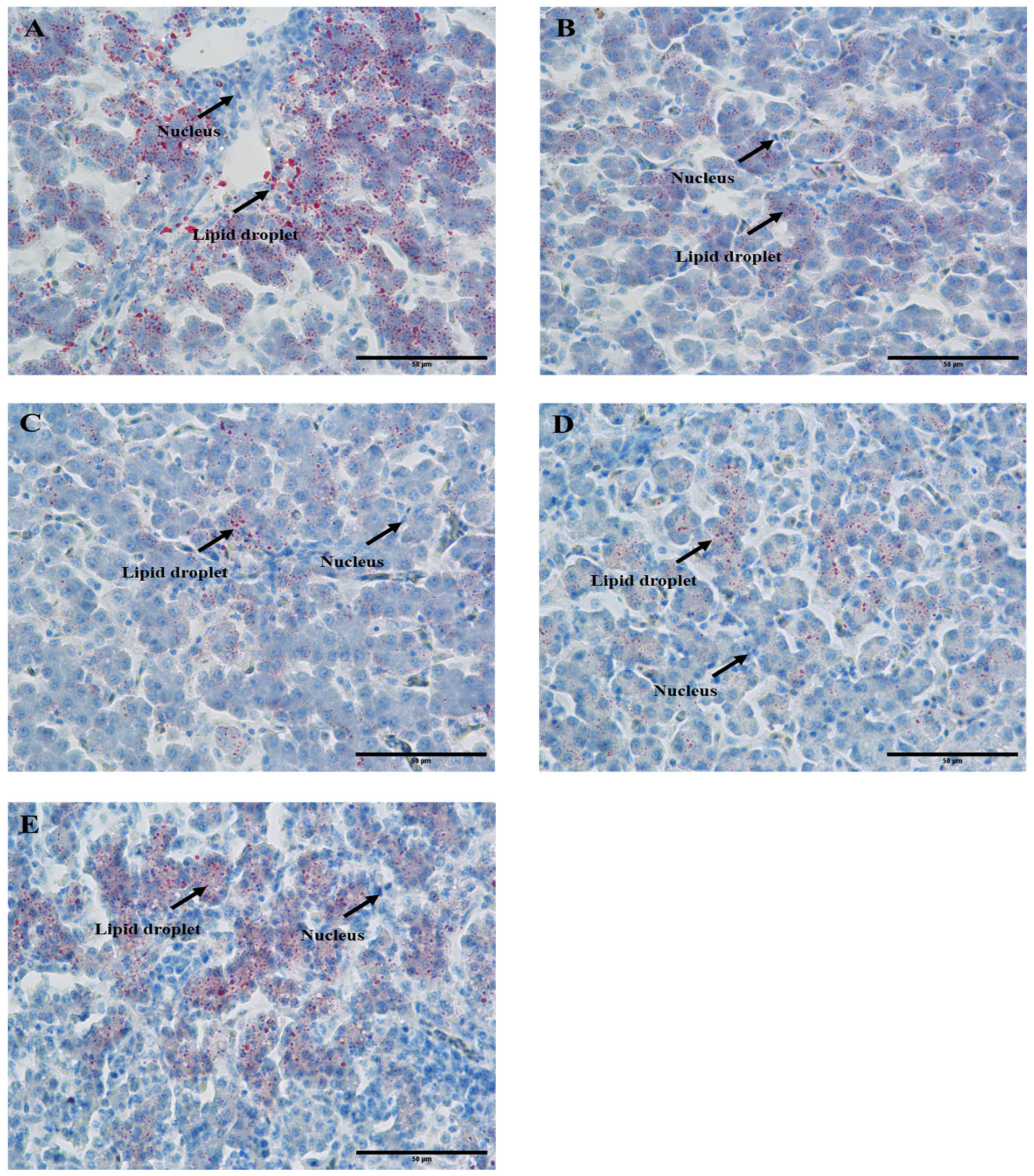
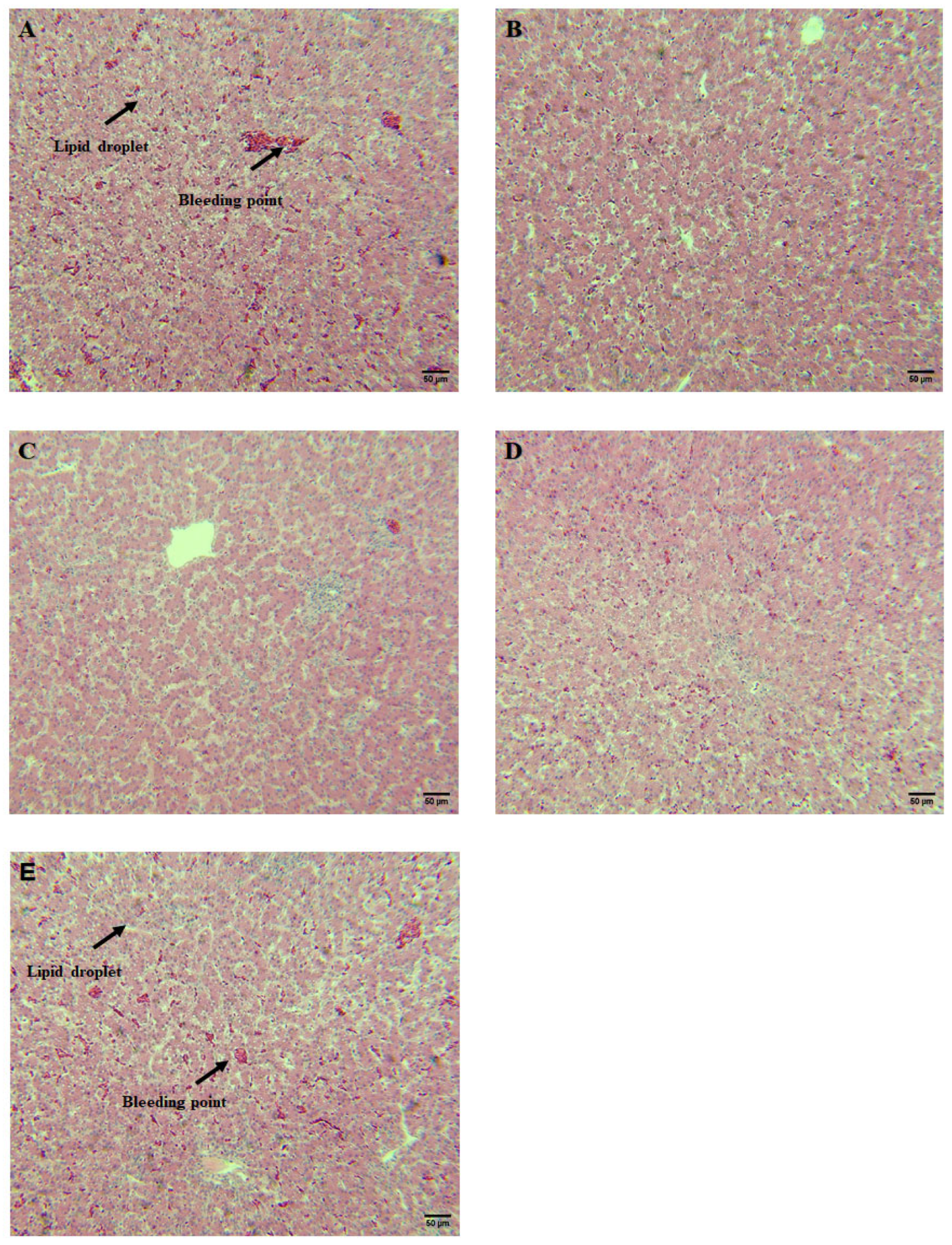
3.2. CSB Decreased Liver Lipid Droplets
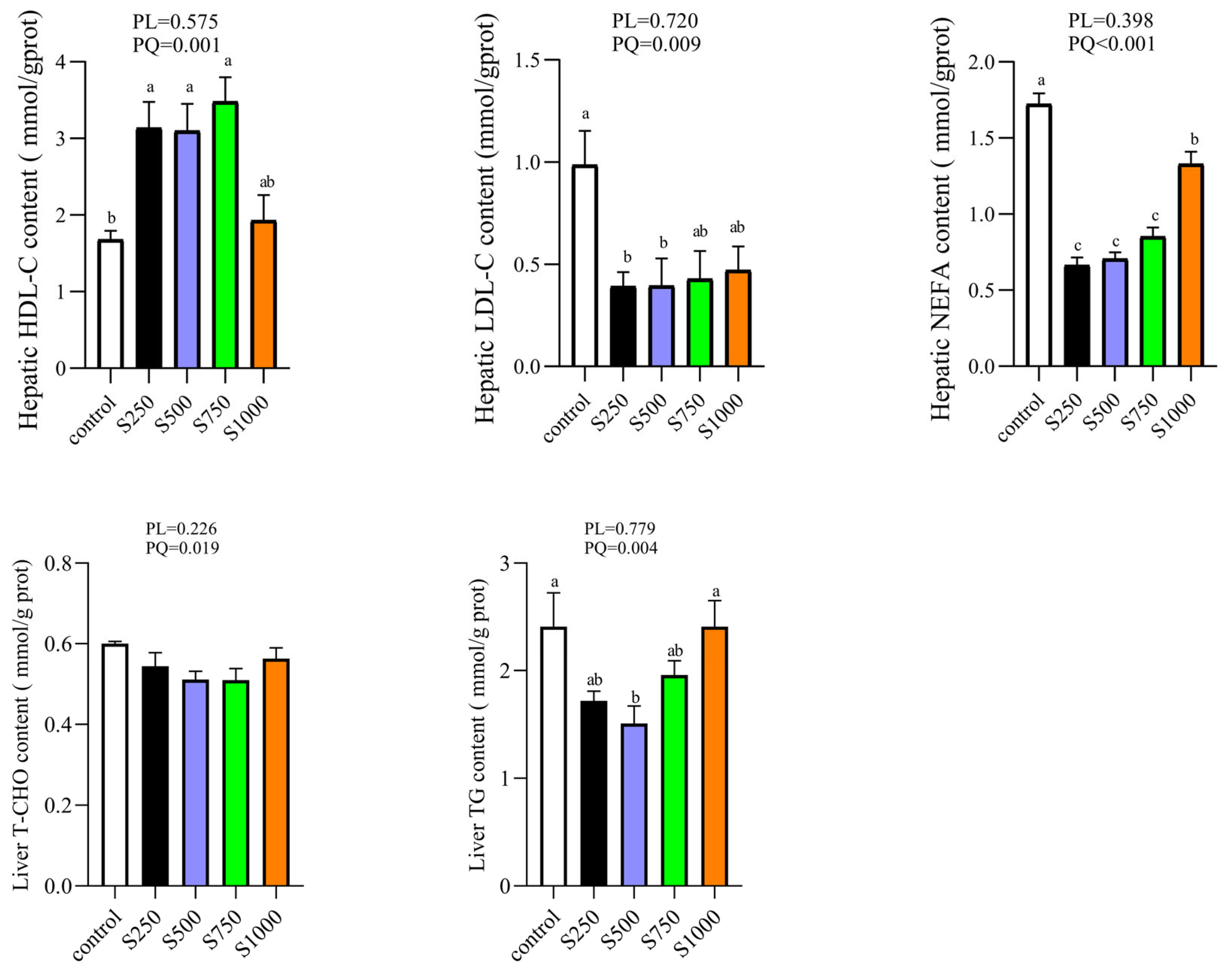
3.3. CSB Regulated Hepatic Lipid Profile
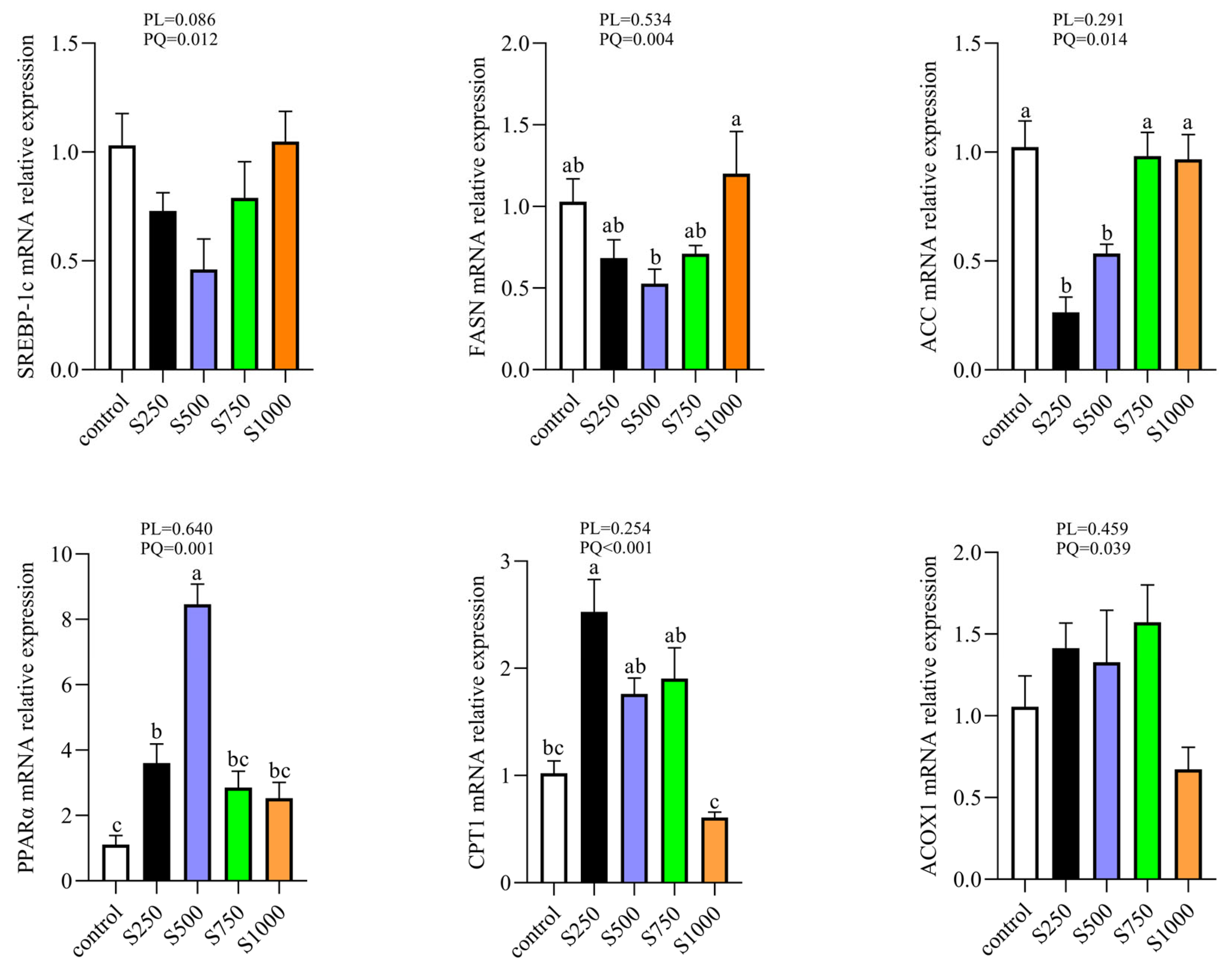
3.4. CSB Alleviated Hepatic Lipid Metabolism
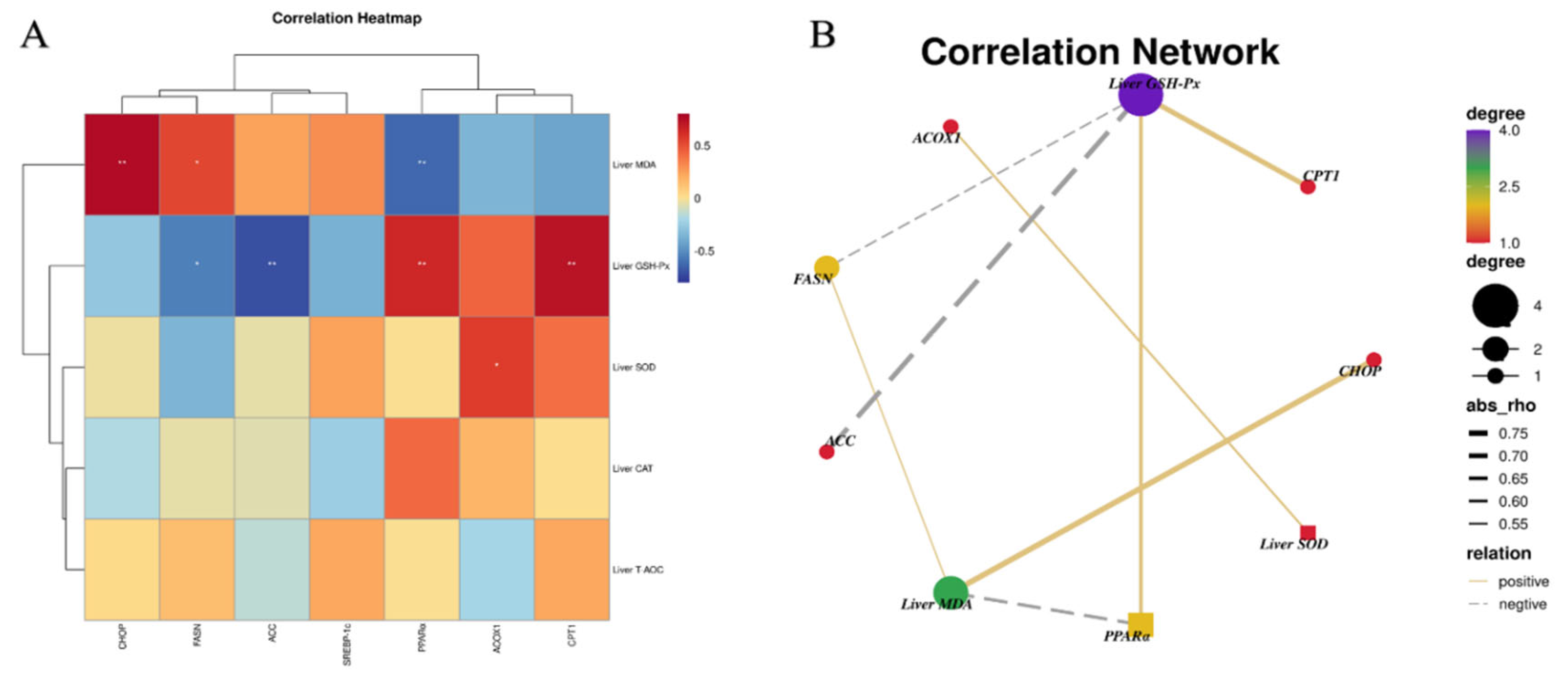
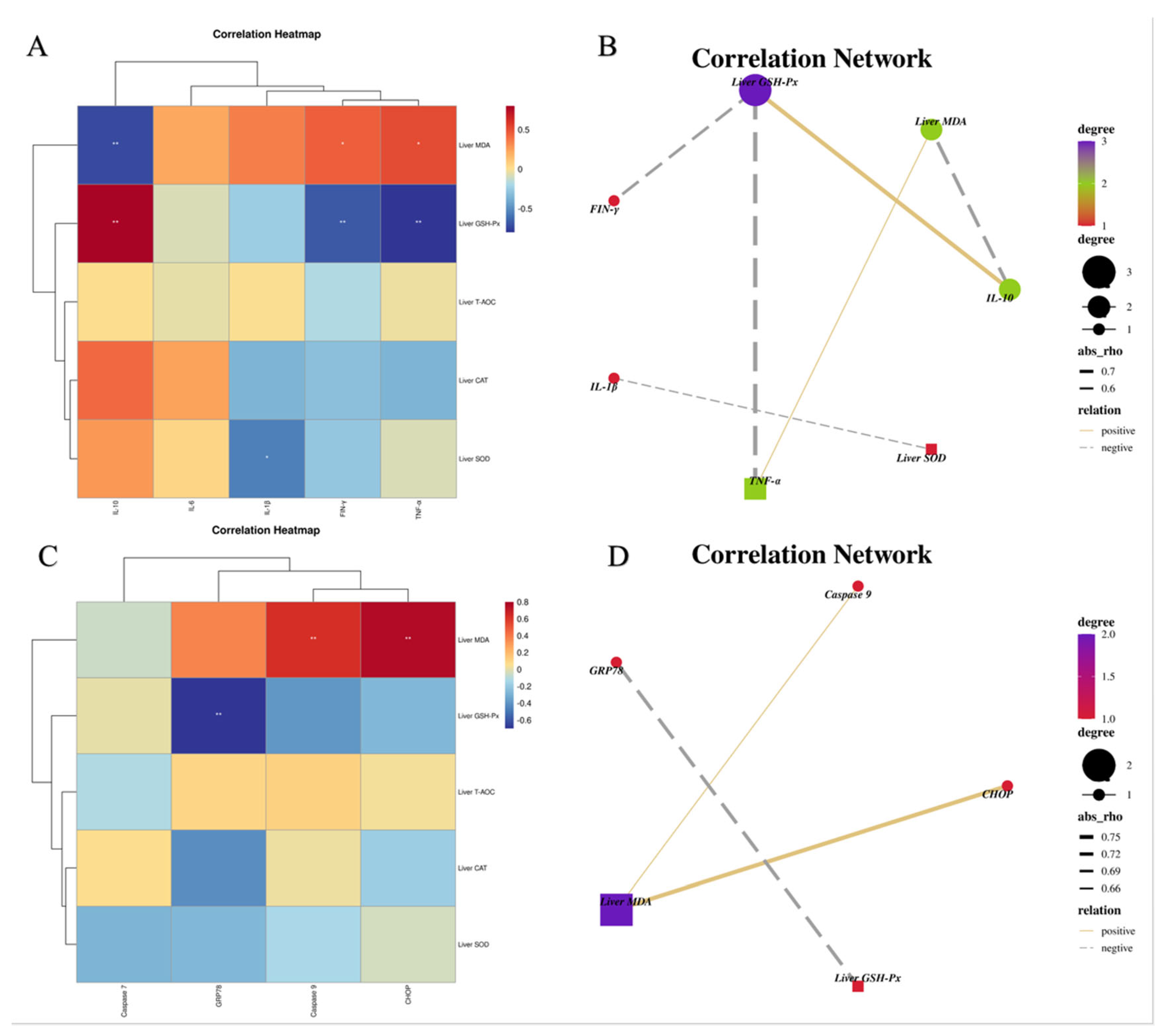
3.5. Relationship between Lipid Metabolism and Liver Antioxidant Indexes
3.6. CSB Attenuated Liver Steatosis and Inflammatory Reaction
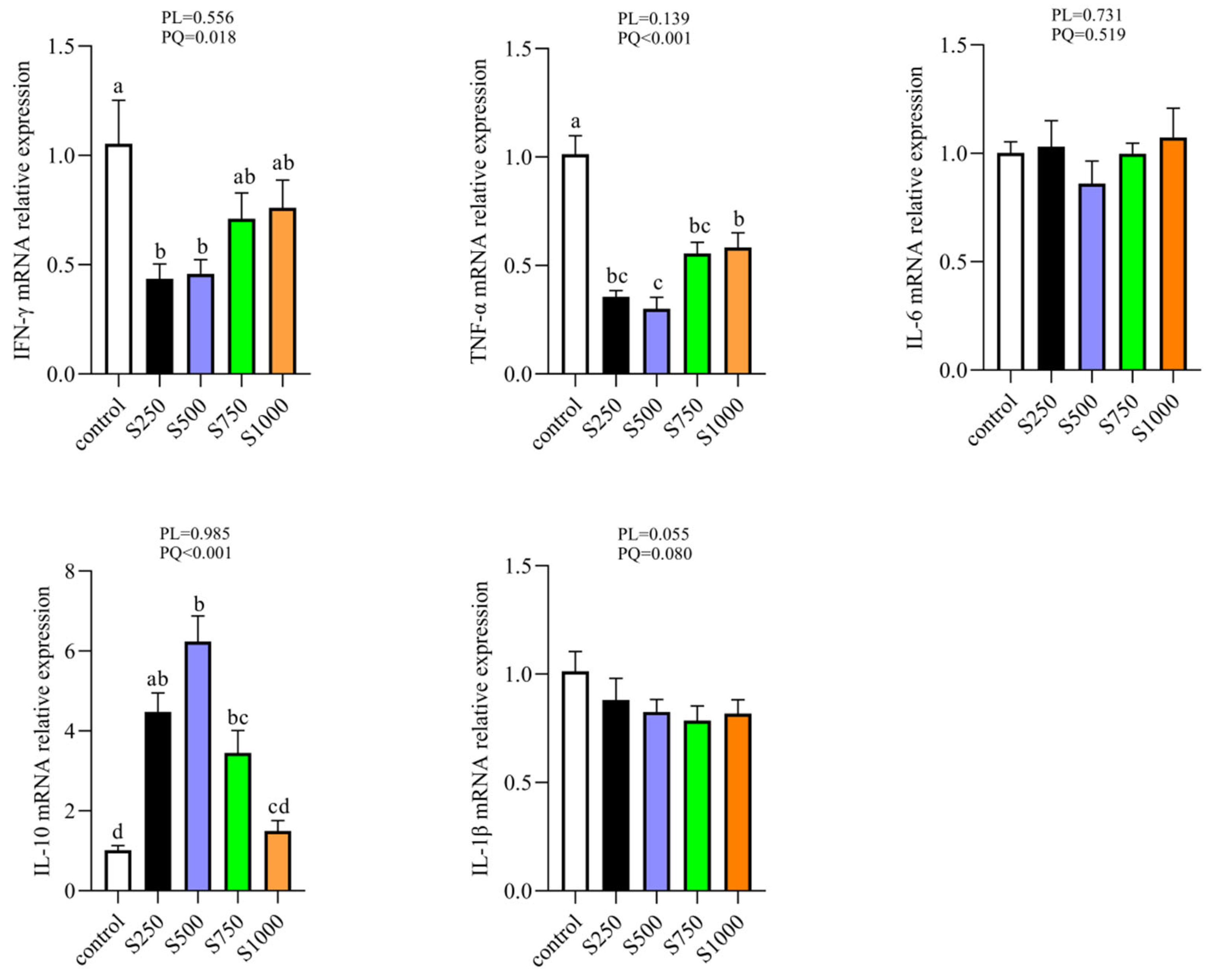
3.7. CSB Enhanced Hepatic Anti-Inflammation Status
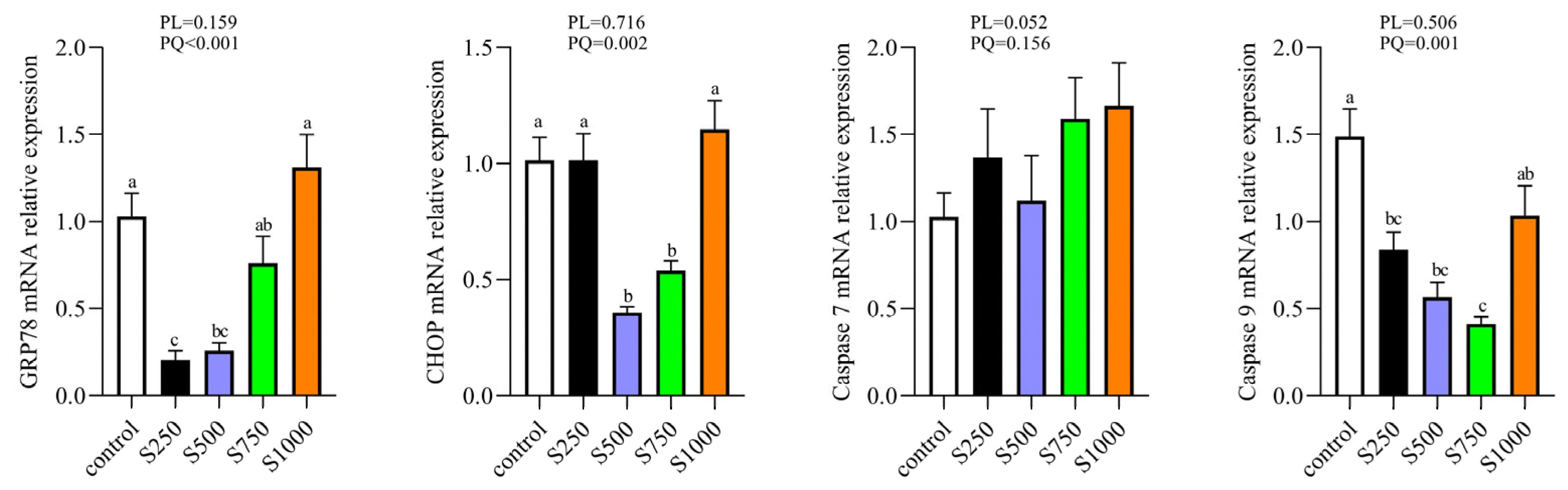
3.8. CSB Regulated ERS of the Liver
3.9. Relationship among Hepatic Inflammatory, ERS Indexes and Antioxidant Parameters
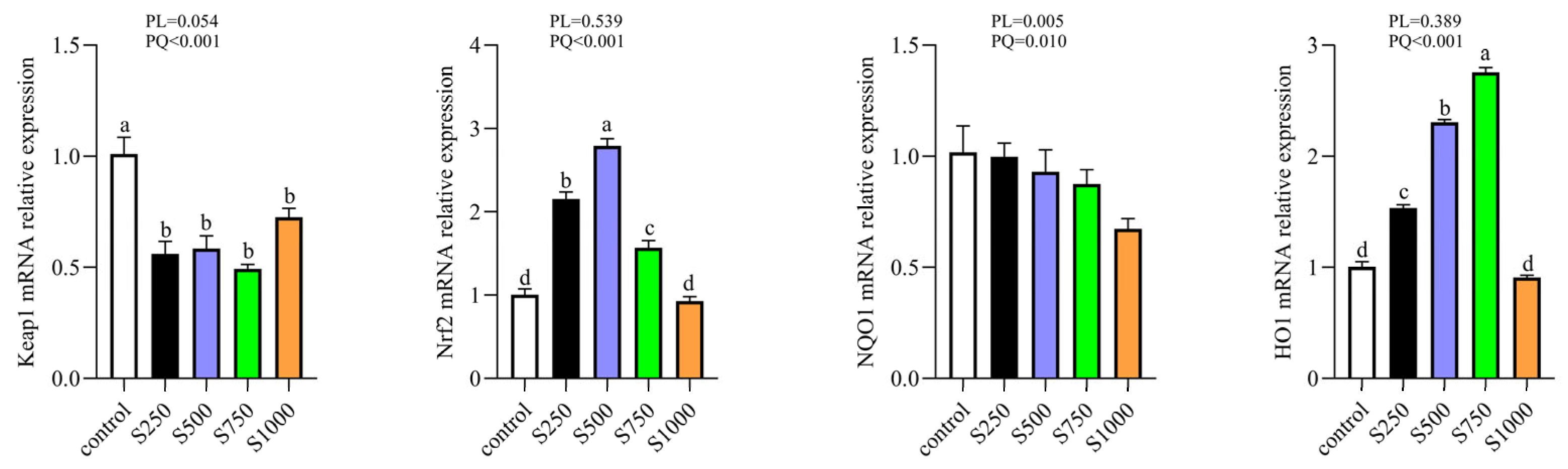
3.10. CSB Regulated the Nrf2 Antioxidant Signaling Pathway
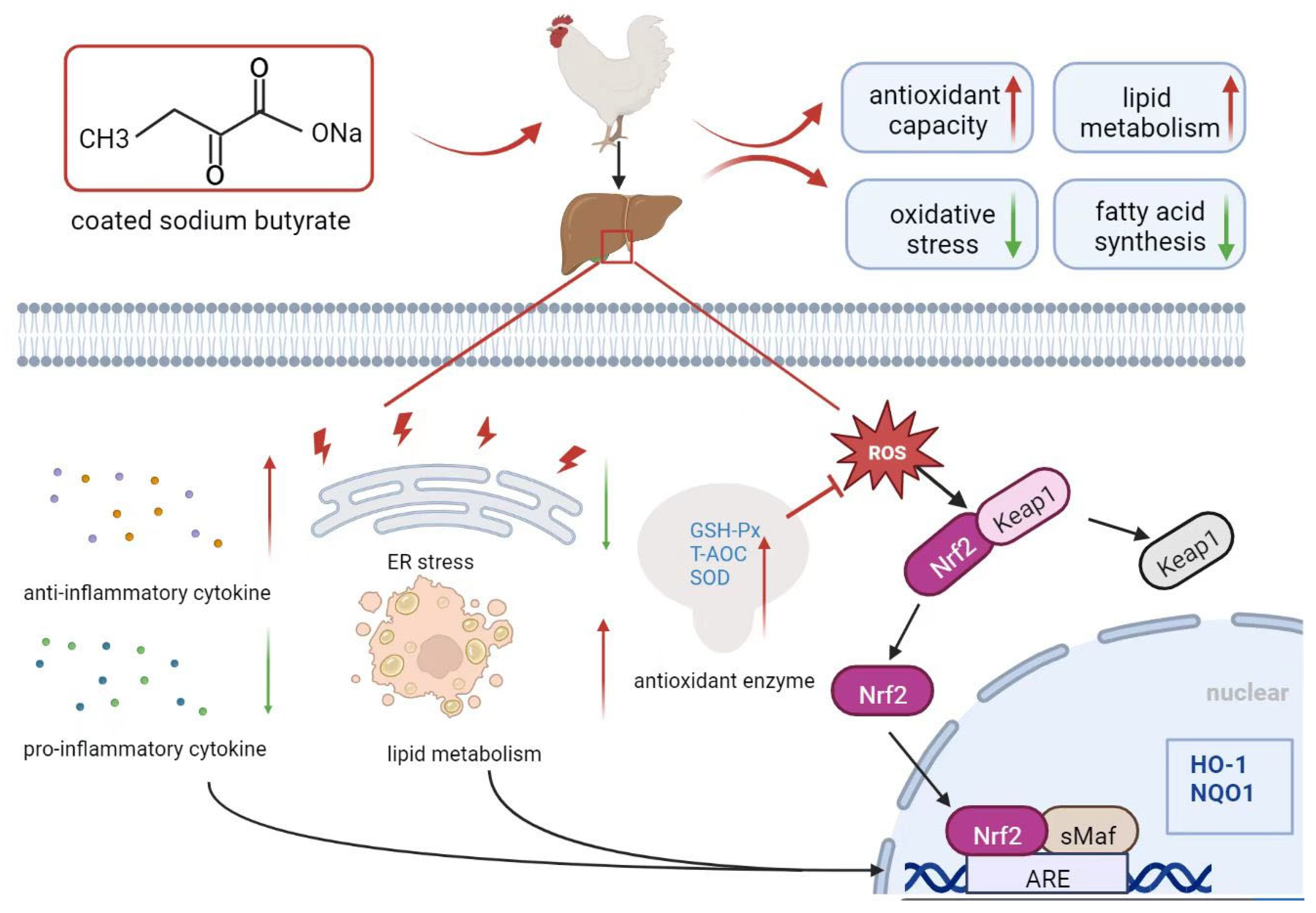
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, Y.; Zhang, X.L.; Du, P.F.; Wang, Z.Y.; Luo, P.N.; Huang, Y.Q.; Liu, Z.H.; Zhang, H.Y.; Chen, W. Dietary herbaceous mixture supplementation reduced hepatic lipid deposition and improved hepatic health status in post-peak laying hens. Poult. Sci. 2022, 101, 6. [Google Scholar] [CrossRef] [PubMed]
- Shini, A.; Shini, S.; Bryden, W.L. Fatty liver haemorrhagic syndrome occurrence in laying hens: Impact of production system. Avian. Pathol. 2019, 48, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Xing, C.; Cao, H.; Zhang, C.; Luo, J.; Guo, X.; Hu, G. Insulin resistance and metabonomics analysis of fatty liver haemorrhagic syndrome in laying hens induced by a high-energy low-protein diet. Sci. Rep. 2019, 9, 10141. [Google Scholar] [CrossRef]
- Gao, X.; Liu, P.; Wu, C.; Wang, T.; Liu, G.; Cao, H.; Zhang, C.; Hu, G.; Guo, X. Effects of fatty liver hemorrhagic syndrome on the AMP-activated protein kinase signaling pathway in laying hens. Poult. Sci. 2019, 98, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Wu, J.; Jiao, H.; Wang, X.; Zhao, J.; Lin, H. The development of antioxidant system in the intestinal tract of broiler chickens. Poult. Sci. 2019, 98, 664–678. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.F.; Chen, Y.P.; Jin, R.; Wang, C.; Wen, C.; Zhou, Y.M. Age-related changes in liver metabolism and antioxidant capacity of laying hens. Poult. Sci. 2021, 100, 101478. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free. Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Trott, K.A.; Giannitti, F.; Rimoldi, G.; Hill, A.; Woods, L.; Barr, B.; Anderson, M.; Mete, A. Fatty liver hemorrhagic syndrome in the backyard chicken: A retrospective histopathologic case series. Vet. Pathol. 2014, 51, 787–795. [Google Scholar] [CrossRef]
- Elnesr, S.S.; Ropy, A.; Abdel-Razik, A.H. Effect of dietary sodium butyrate supplementation on growth, blood biochemistry, hematology andzz histomorphometry of intestine and immune organs of Japanese quail. Animals 2019, 13, 1234–1244. [Google Scholar]
- Lan, R.X.; Li, S.Q.; Zhao, Z.; An, L.L. Sodium butyrate as an effective feed additive to improve growth performance and gastrointestinal development in broilers. Vet. Med. Sci. 2020, 6, 491–499. [Google Scholar] [CrossRef]
- Lin, F.; Li, X.; Wen, J.; Wang, C.; Peng, Y.; Feng, J.; Hu, C. Effects of coated sodium butyrate on performance, diarrhea, intestinal microflora and barrier function of pigs during the first 2-week post-weaning. Anim. Feed. Sci. Technol. 2020, 263, 114464. [Google Scholar] [CrossRef]
- Zhang, W.H.; Gao, F.; Zhu, Q.F.; Li, C.; Jiang, Y.; Dai, S.F.; Zhou, G.H. Dietary sodium butyrate alleviates the oxidative stress induced by corticosterone exposure and improves meat quality in broiler chickens. Poult. Sci. 2011, 90, 11. [Google Scholar] [CrossRef] [PubMed]
- Mattace Raso, G.; Simeoli, R.; Russo, R.; Iacono, A.; Santoro, A.; Paciello, O.; Ferrante, M.C.; Canani, R.B.; Calignano, A.; Meli, R. Effects of sodium butyrate and its synthetic amide derivative on liver inflammation and glucose tolerance in an animal model of steatosis induced by high fat diet. PLoS ONE 2013, 8, e68626. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, C.; Pedroso, A.A.; Mallo, J.J.; Puyalto, M.; Kim, W.K.; Applegate, T.J. Sodium butyrate improved performance while modulating the cecal microbiota and regulating the expression of intestinal immune-related genes of broiler chickens. Poult. Sci. 2017, 96, 11. [Google Scholar] [CrossRef]
- Zhang, W.H.; Jiang, Y.; Zhu, Q.F.; Gao, F.; Dai, S.F.; Chen, J.; Zhou, G.H. Sodium butyrate maintains growth performance by regulating the immune response in broiler chickens. Br. Poult. Sci. 2011, 52, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, S.; Cheng, Y.C.; Yakout, H.M.; Kim, S.W. 367 Combinational use of sodium butyrate and phytogenics on intestinal health of nursery pigs. J. Anim. Sci. 2019, 97, 132–133. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, K.Y.; Wang, J.P.; Bai, S.P.; Zeng, Q.F.; Peng, H.W.; Zhang, B.; Xuan, Y.; Ding, X.M. Effects of coated sodium butyrate on performance, egg quality, nutrient digestibility, and intestinal health of laying hens. Poult. Sci. 2022, 101, 102020. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Y.L.; Liu, K.P.; Fan, R.B.; Li, Q.; Zhou, Z.L. Dietary sodium butyrate and/or vitamin D3 supplementation alters growth performance, meat quality, chemical composition, and oxidative stability in broilers. Food Chem. 2022, 390, 133138. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, Y.M.; Mo, Q.; Kulyar, M.F.; He, Y.Y.; Yao, W.Y.; Quan, C.X.; Gong, S.S.; Li, F.R.; Fu, Y.H.; et al. Sodium butyrate ameliorates thiram-induced tibial dyschondroplasia and gut microbial dysbiosis in broiler chickens. Ecotoxicol. Environ. Saf. 2022, 245, 114134. [Google Scholar] [CrossRef]
- Lee, B.K.; Kim, J.S.; Ahn, H.J.; Hwang, J.H.; Kim, J.M.; Lee, H.T.; An, B.K.; Kang, C.W. Changes in hepatic lipid parameters and hepatic messenger ribonucleic acid expression following estradiol administration in laying hens (Gallus domesticus). Poult. Sci. 2010, 89, 2660–2667. [Google Scholar] [CrossRef]
- Liu, J.; Fu, Y.; Zhou, S.; Zhao, P.; Zhao, J.; Yang, Q.; Wu, H.; Ding, M.; Li, Y. Comparison of the effect of quercetin and daidzein on production performance, anti-oxidation, hormones, and cecal microflora in laying hens during the late laying period. Poult. Sci. 2023, 102, 102674. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.P.; Gong, Y.J.; Li, H.Y.; Xie, C.; Xu, Q.Q.; Dong, X.Y.; Elwan, H.A.M.; Zou, X.T. Alterations in cecal microbiota and intestinal barrier function of laying hens fed on fluoride supplemented diets. Ecotox. Environ. Saf. 2020, 193, 110372. [Google Scholar] [CrossRef] [PubMed]
- Miao, S.S.; Zhou, W.T.; Li, H.Y.; Zhu, M.K.; Dong, X.Y.; Zou, X.T. Effects of coated sodium butyrate on production performance, egg quality, serum biochemistry, digestive enzyme activity, and intestinal health of laying hens. Ital. J. Anim. Sci. 2021, 20, 1452–1461. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Xiao, Y.; Peng, Y.; He, J.; Chen, C.; Xiao, D.; Yin, Y.; Li, F. Mulberry leaf powder regulates antioxidative capacity and lipid metabolism in finishing pigs. Anim. Nutr. 2021, 7, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lu, M.; Wang, J.; Zhang, H.; Qiu, K.; Qi, G.; Wu, S. Dietary oregano essential oil supplementation improves intestinal functions and alters gut microbiota in late-phase laying hens. J. Anim. Sci. Biotechnol. 2021, 12, 72. [Google Scholar] [CrossRef]
- Koizumi, T.; Shirakura, H.; Kumagai, H.; Tatsumoto, H.; Suzuki, K.T. Mechanism of cadmium-induced cytotoxicity in rat hepatocytes: Cadmium-induced active oxygen-related permeability changes of the plasma membrane. Toxicology 1996, 114, 125–134. [Google Scholar] [CrossRef]
- Ma, N.; Abaker, J.A.; Bilal, M.S.; Dai, H.Y.; Shen, X.Z. Sodium butyrate improves antioxidant stability in sub-acute ruminal acidosis in dairy goats. BMC Vet. Res. 2018, 14, 275. [Google Scholar] [CrossRef]
- Li, H.; Gu, Y.; Jin, R.; He, Q.; Zhou, Y. Effects of Dietary Rutin Supplementation on the Intestinal Morphology, Antioxidant Capacity, Immunity, and Microbiota of Aged Laying Hens. Antioxidants 2022, 11, 1843. [Google Scholar] [CrossRef]
- Adeyanju, O.A.; Badejogbin, O.C.; Areola, D.E.; Olaniyi, K.S.; Dibia, C.; Soetan, O.A.; Oniyide, A.A.; Michael, O.S.; Olatunji, L.A.; Soladoye, A.O. Sodium butyrate arrests pancreato-hepatic synchronous uric acid and lipid dysmetabolism in high fat diet fed Wistar rats. Biomed. Pharmacother. 2021, 133, 110994. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Y.; Zhou, Y.; Zhang, Z.; Xie, Z.; Zhang, J.; Wan, X. Green tea polyphenols alleviate obesity in broiler chickens through the regulation of lipid-metabolism-related genes and transcription factor expression. J. Agric. Food. Chem. 2013, 61, 8565–8572. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Huang, S.; Zhao, G.; Ma, Q. Dietary supplementation of porcine bile acids improves laying performance, serum lipid metabolism and cecal microbiota in late-phase laying hens. Anim. Nutr. 2022, 11, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Gao, S.; Chen, J.; Zhao, R.; Yang, X. Maternal sodium butyrate supplement elevates the lipolysis in adipose tissue and leads to lipid accumulation in offspring liver of weaning-age rats. Lipids Health Dis. 2016, 15, 119. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, X.B.; Peng, Z.C.; Fu, S.P.; Zhao, C.X.; Du, X.L.; Fang, Z.Y.; Wang, Z.; Liu, G.W.; Li, X.W. Berberine Protects against NEFA-Induced Impairment of Mitochondrial Respiratory Chain Function and Insulin Signaling in Bovine Hepatocytes. Int. J. Mol. Sci. 2018, 19, 1691. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, R.; Bril, F.; Portillo-Sanchez, P.; Ortiz-Lopez, C.; Orsak, B.; Biernacki, D.; Lo, M.; Suman, A.; Weber, M.H.; Cusi, K. Metabolic Impact of Nonalcoholic Steatohepatitis in Obese Patients with Type 2 Diabetes. Diabetes Care 2016, 39, 632–638. [Google Scholar] [CrossRef]
- Li, C.; Yang, W.; Zhang, J.; Zheng, X.; Yao, Y.; Tu, K.; Liu, Q. SREBP-1 has a prognostic role and contributes to invasion and metastasis in human hepatocellular carcinoma. Int. J. Mol. Sci. 2014, 15, 7124–7138. [Google Scholar] [CrossRef]
- Huang, Y.; Gao, S.; Chen, J.; Albrecht, E.; Zhao, R.; Yang, X. Maternal butyrate supplementation induces insulin resistance associated with enhanced intramuscular fat deposition in the offspring. Oncotarget 2017, 8, 13073–13084. [Google Scholar] [CrossRef]
- Hunkeler, M.; Hagmann, A.; Stuttfeld, E.; Chami, M.; Guri, Y.; Stahlberg, H.; Maier, T. Structural basis for regulation of human acetyl-CoA carboxylase. Nature 2018, 558, 470–474. [Google Scholar] [CrossRef]
- Schadinger, S.E.; Bucher, N.L.; Schreiber, B.M.; Farmer, S.R. PPARgamma2 regulates lipogenesis and lipid accumulation in steatotic hepatocytes. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E1195–E1205. [Google Scholar] [CrossRef]
- Schoonjans, K.; Staels, B.; Auwerx, J. Role of the peroxisome proliferator-activated receptor (PPAR) in mediating the effects of fibrates and fatty acids on gene expression. J. Lipid Res. 1996, 37, 907–925. [Google Scholar] [CrossRef]
- Louet, J.F.; Chatelain, F.; Decaux, J.F.; Park, E.A.; Kohl, C.; Pineau, T.; Girard, J.; Pegorier, J.P. Long-chain fatty acids regulate liver carnitine palmitoyltransferase I gene (L-CPT I) expression through a peroxisome-proliferator-activated receptor alpha (PPARalpha)-independent pathway. Biochem. J. 2001, 354, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S. Effects of fatty acids on gene expression: Role of peroxisome proliferator-activated receptor alpha, liver X receptor alpha and sterol regulatory element-binding protein-1c. Proc. Nutr. Soc. 2002, 61, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Dietary factors and low grade inflammation in relation to overweight and obesity revisted. Br. J. Nutr. 2022, 127, 1455–1457. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.W.; Dixit, V.D. Adipose tissue as an immunological organ. Obesity 2015, 23, 512–518. [Google Scholar] [CrossRef]
- Zhao, H.W.; Yue, Y.H.; Han, H.; Chen, X.L.; Lu, Y.G.; Zheng, J.M.; Hou, H.T.; Lang, X.M.; He, L.L.; Hu, Q.L.; et al. Effect of toll-like receptor 3 agonist poly I:C on intestinal mucosa and epithelial barrier function in mouse models of acute colitis. World J. Gastroenterol. 2017, 23, 999–1009. [Google Scholar] [CrossRef]
- Yang, C.; Feng, Q.; Liao, H.; Yu, X.; Liu, Y.; Wang, D. Anti-Diabetic Nephropathy Activities of Polysaccharides Obtained from Termitornyces albuminosus via Regulation of NF-κB Signaling in db/db Mice. Int. J. Mol. Sci. 2019, 20, 5205. [Google Scholar] [CrossRef]
- Uscátegui, Y.L.; Díaz, L.E.; Gómez-Tejedor, J.A.; Vallés-Lluch, A.; Vilariño-Feltrer, G.; Serrano, M.A.; Valero, M.F. Candidate Polyurethanes Based on Castor Oil (Ricinus communis), with Polycaprolactone Diol and Chitosan Additions, for Use in Biomedical Applications. Molecules 2019, 24, 237. [Google Scholar] [CrossRef]
- Dong, Y.; Han, Y.; Wang, Z.; Qin, Z.; Yang, C.; Cao, J.; Chen, Y. Role of serotonin on the intestinal mucosal immune response to stress-induced diarrhea in weaning mice. BMC Gastroenterol. 2017, 17, 82. [Google Scholar] [CrossRef]
- Wang, H.; Vilches-Moure, J.G.; Cherkaoui, S.; Tardy, I.; Alleaume, C.; Bettinger, T.; Lutz, A.; Paulmurugan, R. Chronic Model of Inflammatory Bowel Disease in IL-10−/− Transgenic Mice: Evaluation with Ultrasound Molecular Imaging. Theranostics 2019, 9, 6031–6046. [Google Scholar] [CrossRef]
- Gharib-Naseri, K.; Kheravii, S.K.; Li, L.; Wu, S.B. Buffered formic acid and a monoglyceride blend coordinately alleviate subclinical necrotic enteritis impact in broiler chickens. Poult. Sci. 2021, 100, 101214. [Google Scholar] [CrossRef]
- Nishitsuji, K.; Xiao, J.; Nagatomo, R.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; Tsuneyama, K. Analysis of the gut microbiome and plasma short-chain fatty acid profiles in a spontaneous mouse model of metabolic syndrome. Sci. Rep. 2017, 7, 15876. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Lv, Y.; Li, N.; Huang, X.; Liu, X.; Li, H.; Wang, C.; Jia, Y.F. Biological investigations on therapeutic effect of chitosan encapsulated nano resveratrol against gestational diabetes mellitus rats induced by streptozotocin. Drug Deliv. 2020, 27, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, X.; Li, F.; Lv, X.; Fu, X.; Gu, H.; Liu, H.; Liu, J.; Dai, M.; Zhang, B. Efficacy of celastrol combined with cisplatin in enhancing the apoptosis of U-2OS osteosarcoma cells via the mitochondrial and endoplasmic reticulum pathways of apoptosis. Oncol. Lett. 2019, 17, 3305–3313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, B.; Wang, M.; Tong, J.; Pan, J.; Wang, N.; Gong, P.; Long, M. Selenium Protects against Zearalenone-Induced Oxidative Stress and Apoptosis in the Mouse Kidney by Inhibiting Endoplasmic Reticulum Stress. Oxid. Med. Cell. Longev. 2020, 2020, 6059058. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, J.; Yuan, Y.; Chen, J.; Cheng, S.; Wang, H.; Xu, Y. Sodium butyrate mitigates type 2 diabetes by inhibiting PERK-CHOP pathway of endoplasmic reticulum stress. Environ. Toxicol. Pharmacol. 2018, 64, 112–121. [Google Scholar] [CrossRef]
- Xing, X.; Jiang, Z.; Tang, X.; Wang, P.; Li, Y.; Sun, Y.; Le, G.; Zou, S. Sodium butyrate protects against oxidative stress in HepG2 cells through modulating Nrf2 pathway and mitochondrial function. J. Physiol. Biochem. 2016, 73, 405–414. [Google Scholar] [CrossRef]
- Hu, H.; Dai, S.; Li, J.; Wen, A.; Bai, X. Glutamine improves heat stress-induced oxidative damage in the broiler thigh muscle by activating the nuclear factor erythroid 2-related 2/Kelch-like ECH-associated protein 1 signaling pathway. Poult. Sci. 2020, 99, 1454–1461. [Google Scholar] [CrossRef]
- Zhao, L.; Xu, L.; Tao, X.; Han, X.; Yin, L.; Qi, Y.; Peng, J. Protective Effect of the Total Flavonoids from Rosa laevigata Michx Fruit on Renal Ischemia-Reperfusion Injury through Suppression of Oxidative Stress and Inflammation. Molecules 2016, 21, 952. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; Yu, J.; Yan, H.; Zheng, P.; Luo, Y. Dietary dihydromyricetin supplementation enhances antioxidant capacity and improves lipid metabolism in finishing pigs. Food Funct. 2021, 12, 6925–6935. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Composition |
|---|---|
| Ingredients | Content (%) |
| Corn | 62 |
| Soybean meal | 24.5 |
| Soybean oil | 0.5 |
| Limestone Premix 1,2 | 8 5 |
| Total | 100 |
| Nutrient 3 | |
| Metabolism energy, MJ/kg | 10.99 |
| Crude protein, % | 15.67 |
| Lysine, % | 0.80 |
| Methionine, % | 0.34 |
| Calcium, % Total phosphorus, % | 3.69 0.54 |
| Target Gene | Primer | Primer Sequence (5′-3′) | Accession No. |
|---|---|---|---|
| β-Actin | Forward | TCCCTGGAGAAGAGCTATGAA | NM_205518.1 |
| Reverse | CAGGACTCCATACCCAAGAAAG | ||
| SREBP-1c | Forward | GCCATCGAGTACATCCGCTT | NM_204126.2 |
| Reverse | GGTCCTTGAGGGACTTGCTC | ||
| FASN | Forward | GAATCCAGAAGGGCCAACGA | NM_205155.4 |
| Reverse | TCCAAGGGAGCAGCTTTTGT | ||
| ACC | Forward | TACAGAGGTACCGGAGTGGT | NM_205505.1 |
| Reverse | TCTTCCCGAAGGGCAAAGAC | ||
| PPARα | Forward | AGGCCAAGTTGAAAGCAGAA | NM_001001464.1 |
| Reverse | TTTCCCTGCAAGGATGACTC | ||
| CPT1 | Forward | GGCTCTGGCAGGAGCTACA | XM_040700878.2 |
| Reverse | CACTGCAGCTGGGATCTTGA | ||
| ACOX1 | Forward | ACTGAGCTGTGTCTCTTGTATG | XM_015295164.2 |
| Reverse | GCTTCAGGTGTTTGTGGAAAG | ||
| IFN-γ | Forward | AGCTGACGGTGGACCTATTATT | NM_205149.1 |
| Reverse | GGCTTTGCGCTGGATTC | ||
| TNF-α | Forward | GACAGCCTATGCCAACAAGTA | AY765397.1 |
| Reverse | TCCACATCTTTCAGAGCATCAA | ||
| IL6 | Forward | CTCGTCCGGAACAACCTCAA | NM_204628.2 |
| Reverse | TCAGGCATTTCTCCTCGTCG | ||
| IL10 | Forward | CCAGGGACGATGAACTTAACA | NM_001004414.2 |
| Reverse | GATGGCTTTGCTCCTCTTCT | ||
| IL1β | Forward | ACTGGGCATCAAGGGCTA | XM_015297469.2 |
| Reverse | GGTAGAAGATGAAGCGGGTC | ||
| GRP78 | Forward | GTTACTGTGCCAGCCTACTT | NM_205491.1 |
| Reverse | CCGCTTCGCTTTCTCTACTT | ||
| Caspase 9 | Forward | GACCTGCTAACCATGCTACTT | XM_424580.6 |
| Reverse | TTCCACTGAATCCTCCAATCC | ||
| Caspase 7 | Forward | TGCAAAGCCAGACAGAAGTAG | XM_025151846.1 |
| Reverse | GGTCCATCGGTGCCATAAAT | ||
| CHOP | Forward | GCACAGCCCATTTCTGTTTC | XM_015273173.2 |
| Reverse | TGCCATCCCATTCTGCTAAG | ||
| Keap1 | Forward | TGCCCCTGTGGTCAAAGTG | XM_015274015.1 |
| Reverse | GGTTCGGTTACCGTCCTGC | ||
| Nrf2 | Forward | TGTGTGTGATTCAACCCGACT | NM_205117.1 |
| Reverse | TTAATGGAAGCCGCACCACT | ||
| HO1 | Forward | TTGGCAAGAAGCATCCAGA | NM_205344.1 |
| Reverse | TCCATCTCAAGGGCATTCA | ||
| NQO1 | Forward | AAAGCCATGCTGTCACTCACC | NM_001277619.1 |
| Reverse | GTAGCGCACGCTGTAGCAAAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, S.; Li, Y.; Mu, T.; Wang, X.; Zhao, W.; Li, R.; Dong, X.; Zou, X. Dietary Coated Sodium Butyrate Ameliorates Hepatic Lipid Accumulation and Inflammation via Enhancing Antioxidative Function in Post-Peaking Laying Hens. Metabolites 2023, 13, 650. https://doi.org/10.3390/metabo13050650
Miao S, Li Y, Mu T, Wang X, Zhao W, Li R, Dong X, Zou X. Dietary Coated Sodium Butyrate Ameliorates Hepatic Lipid Accumulation and Inflammation via Enhancing Antioxidative Function in Post-Peaking Laying Hens. Metabolites. 2023; 13(5):650. https://doi.org/10.3390/metabo13050650
Chicago/Turabian StyleMiao, Sasa, Yan Li, Tianming Mu, Xiaoming Wang, Wenyan Zhao, Ru Li, Xinyang Dong, and Xiaoting Zou. 2023. "Dietary Coated Sodium Butyrate Ameliorates Hepatic Lipid Accumulation and Inflammation via Enhancing Antioxidative Function in Post-Peaking Laying Hens" Metabolites 13, no. 5: 650. https://doi.org/10.3390/metabo13050650
APA StyleMiao, S., Li, Y., Mu, T., Wang, X., Zhao, W., Li, R., Dong, X., & Zou, X. (2023). Dietary Coated Sodium Butyrate Ameliorates Hepatic Lipid Accumulation and Inflammation via Enhancing Antioxidative Function in Post-Peaking Laying Hens. Metabolites, 13(5), 650. https://doi.org/10.3390/metabo13050650