


Evaluation and Comparison of Dermo-Cosmetic Activities of Three Oak Species by Targeting Antioxidant Metabolites and Skin Enzyme Inhibitors


Abstract
:
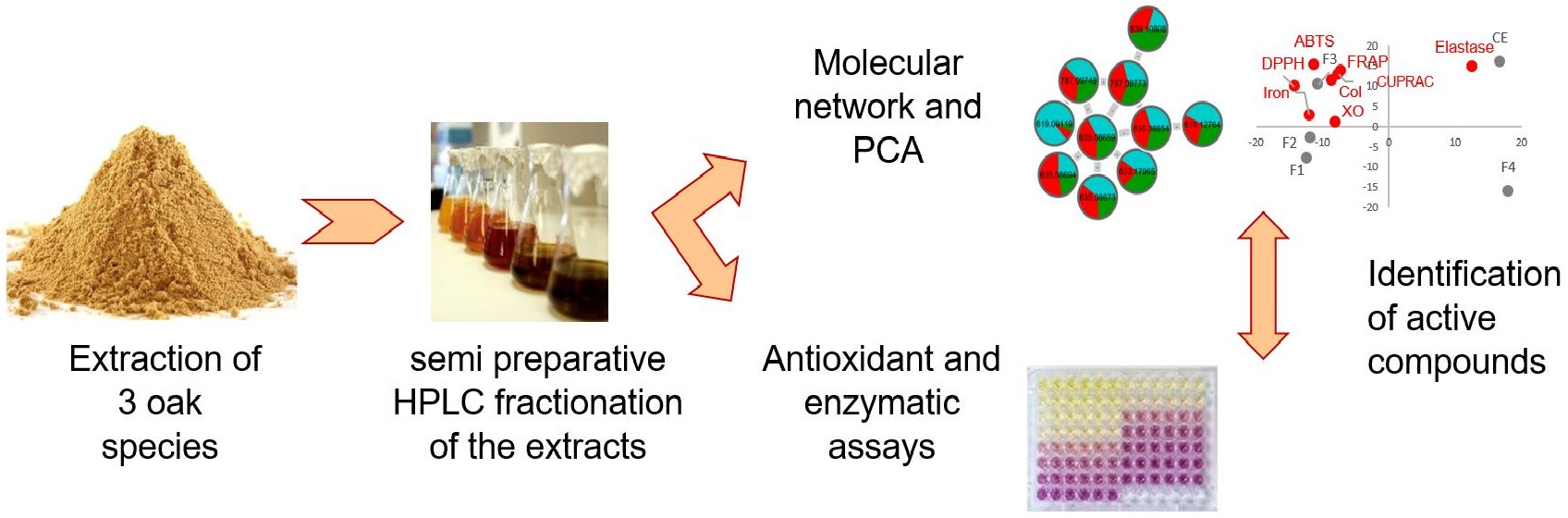{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Sample Preparation
2.3. UHPLC-DAD-ELSD Analysis
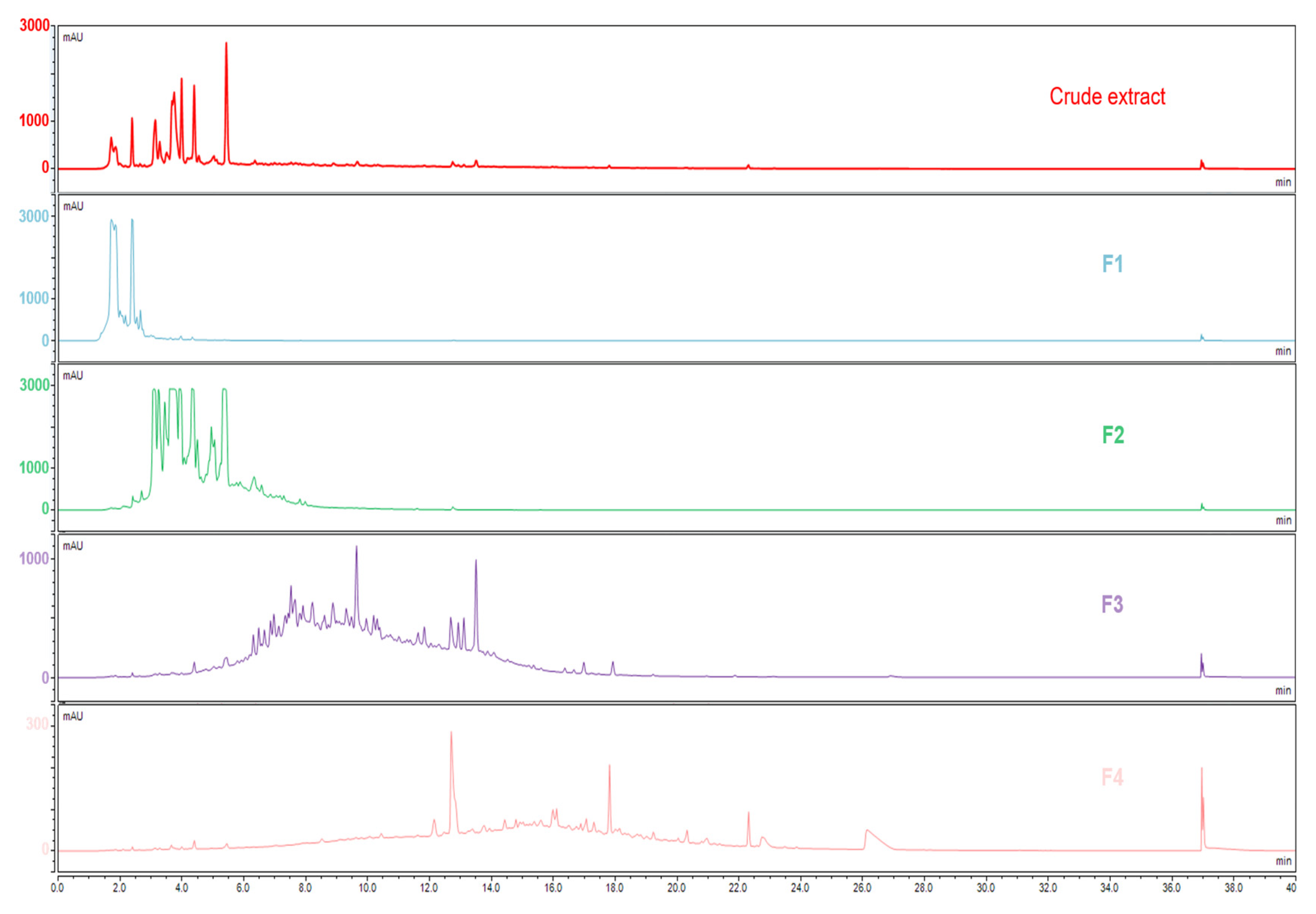
2.4. Fractionation by Semi-Preparative Liquid Chromatography
2.5. In Vitro Antioxidant Capability
2.5.1. DPPH Radical Scavenging Activity Assay
2.5.2. ABTS Radical Cation Decolorization Assay
2.5.3. FRAP (Ferric Ion-Reducing Antioxidant Power) Assay
2.5.4. CUPRAC (Cupric Ion-Reducing Antioxidant Activity) Assay
2.5.5. Iron (II) Chelating Assay
2.6. Enzymatic Tests
2.6.1. Elastase Inhibition Assay
2.6.2. Collagenase Inhibition Assay
2.6.3. Tyrosinase Inhibition Assay
2.6.4. Xanthine Oxidase Assay
2.7. UHPLC-UV-MS/HRMS
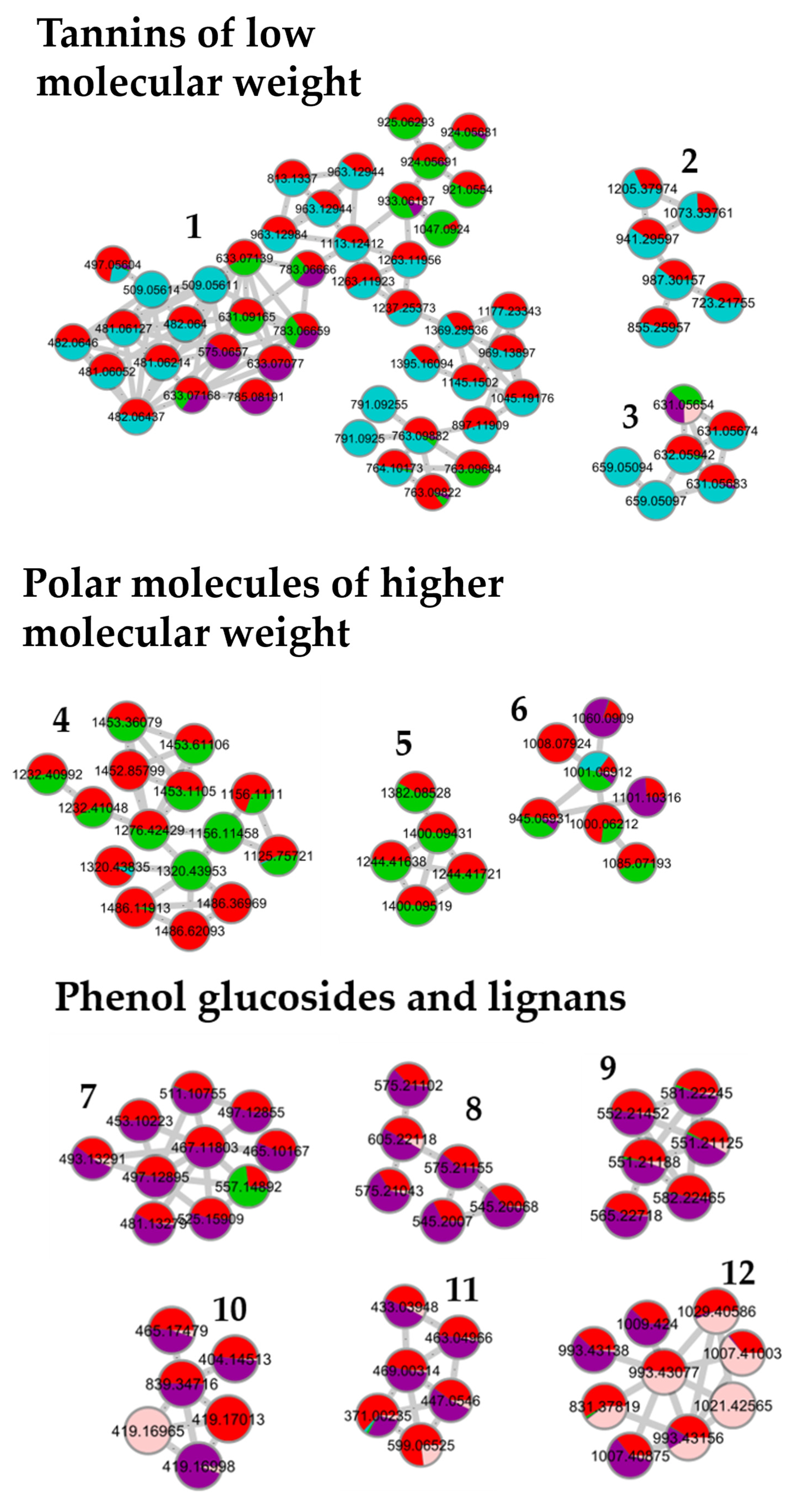
2.8. Molecular Network Design
2.9. Statistical Analysis
3. Results and Discussion
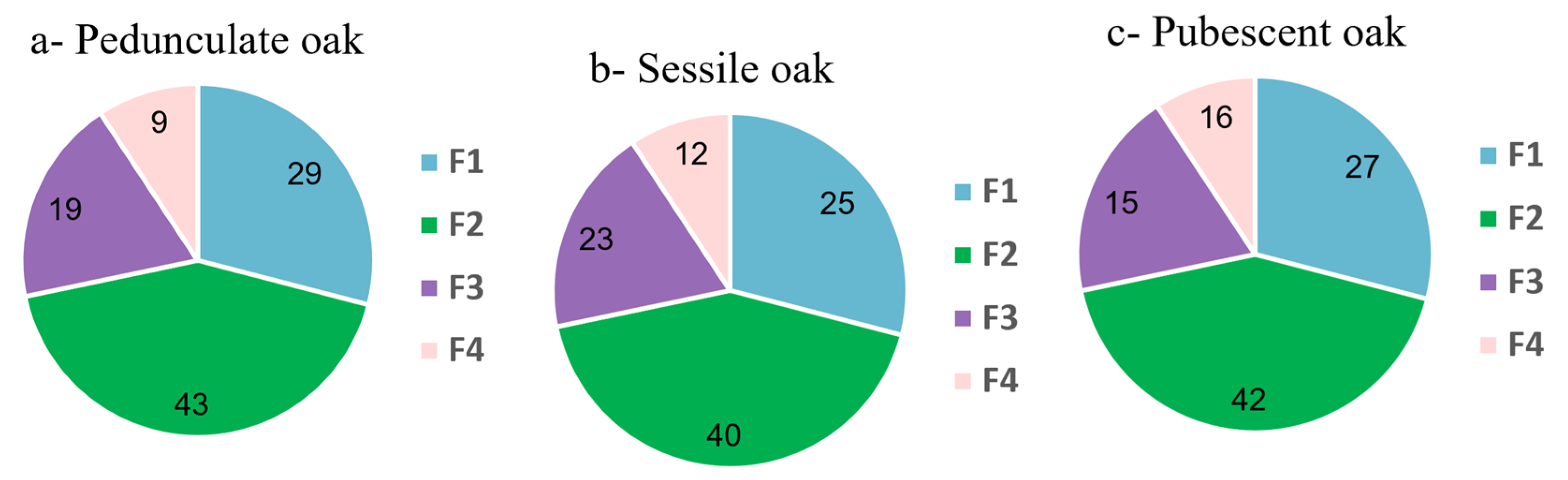
3.1. Extract Fractionation and Fraction Analysis
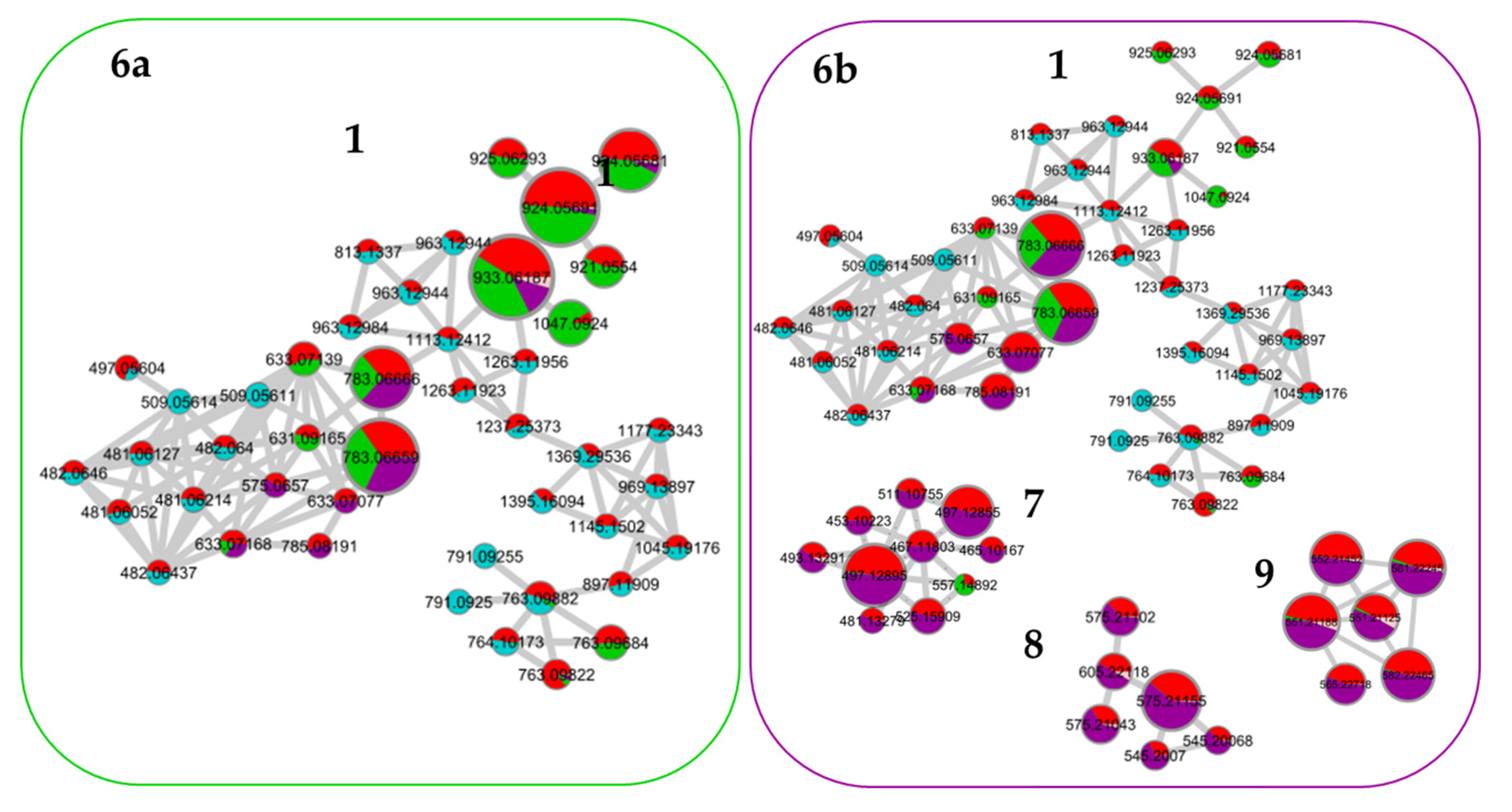
3.2. Comparison of the Chemical Composition of Fractions and Crude Extract
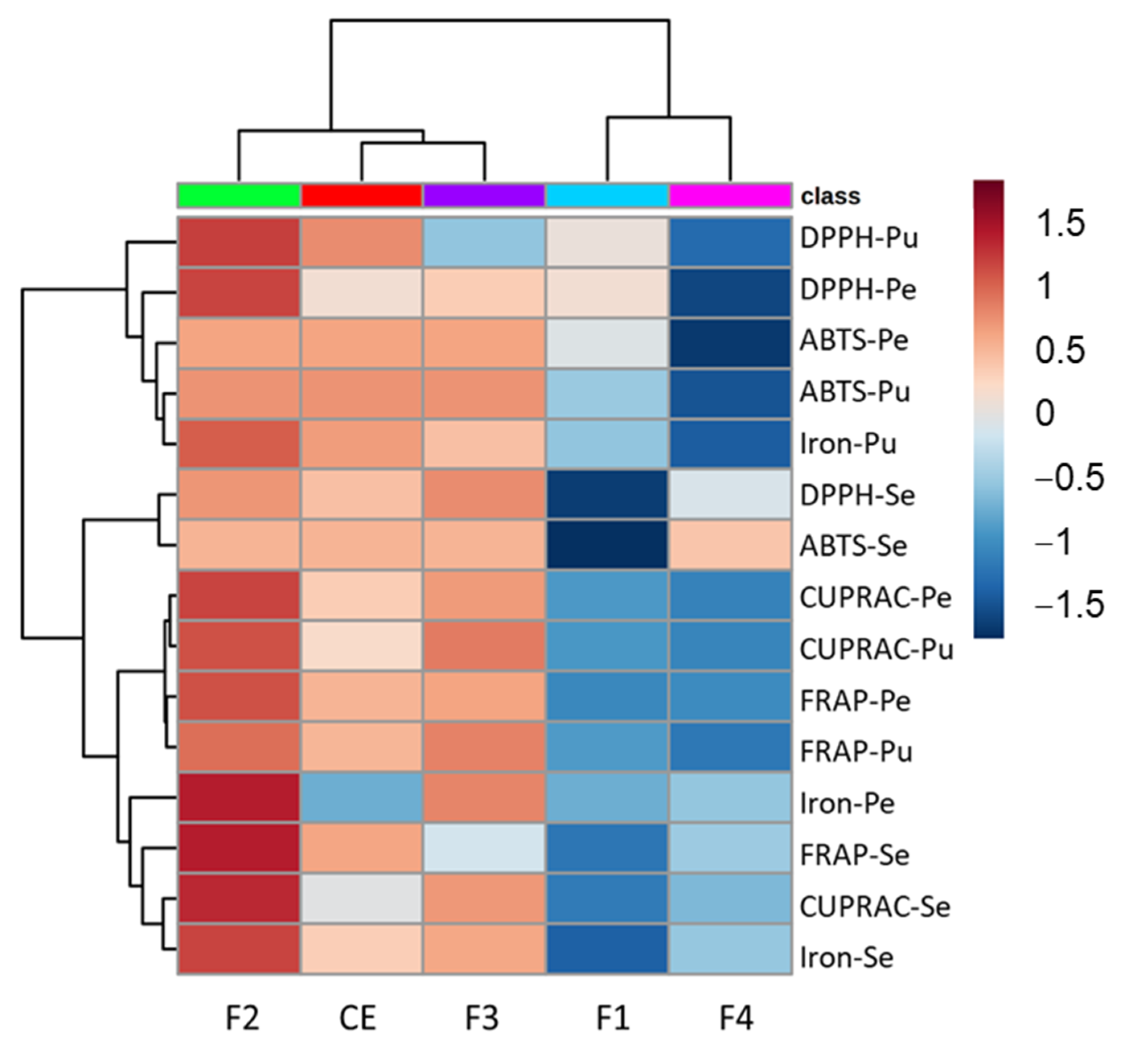
3.3. In Vitro Antioxidant Capability
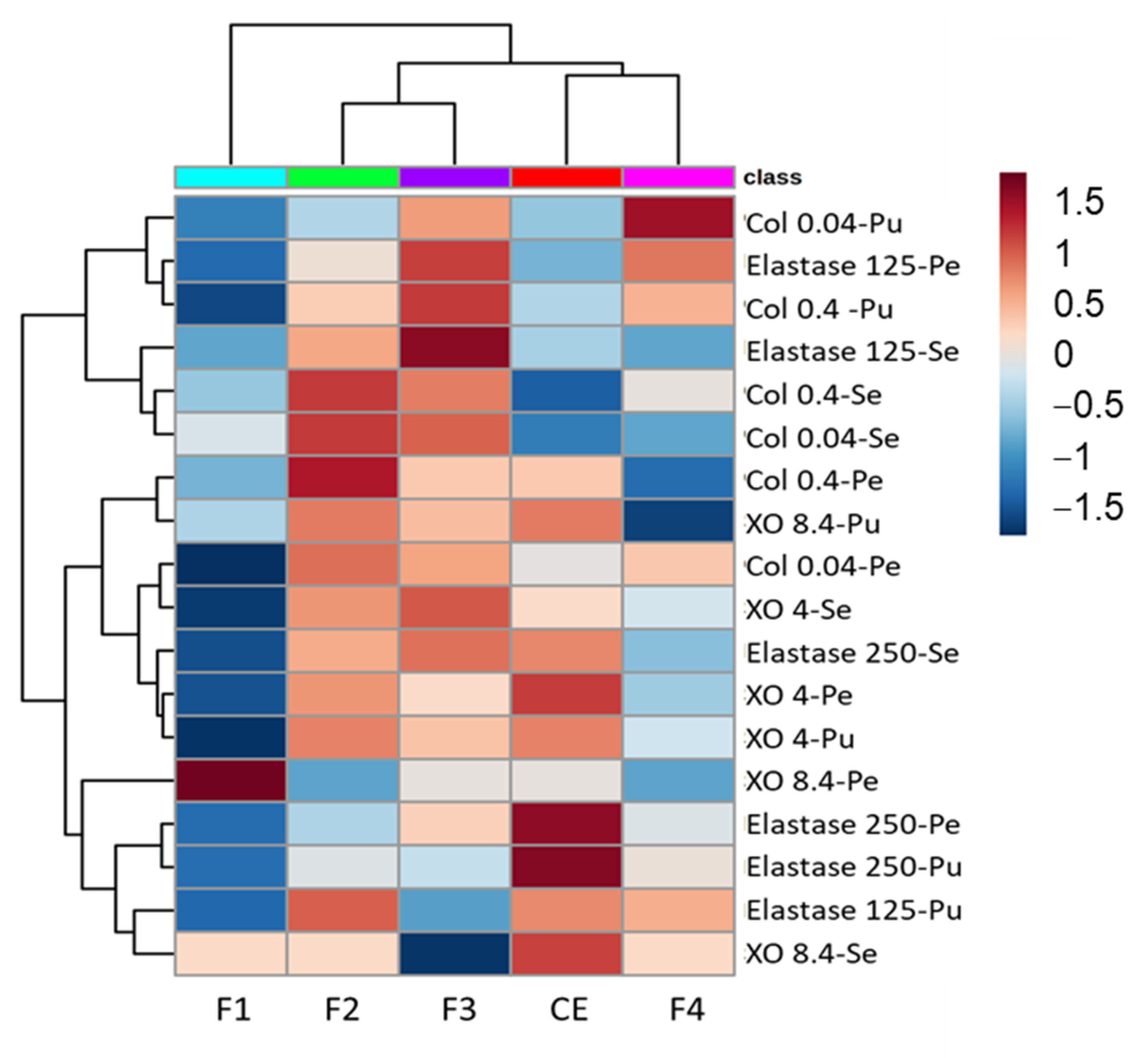
3.4. Enzymatic Tests
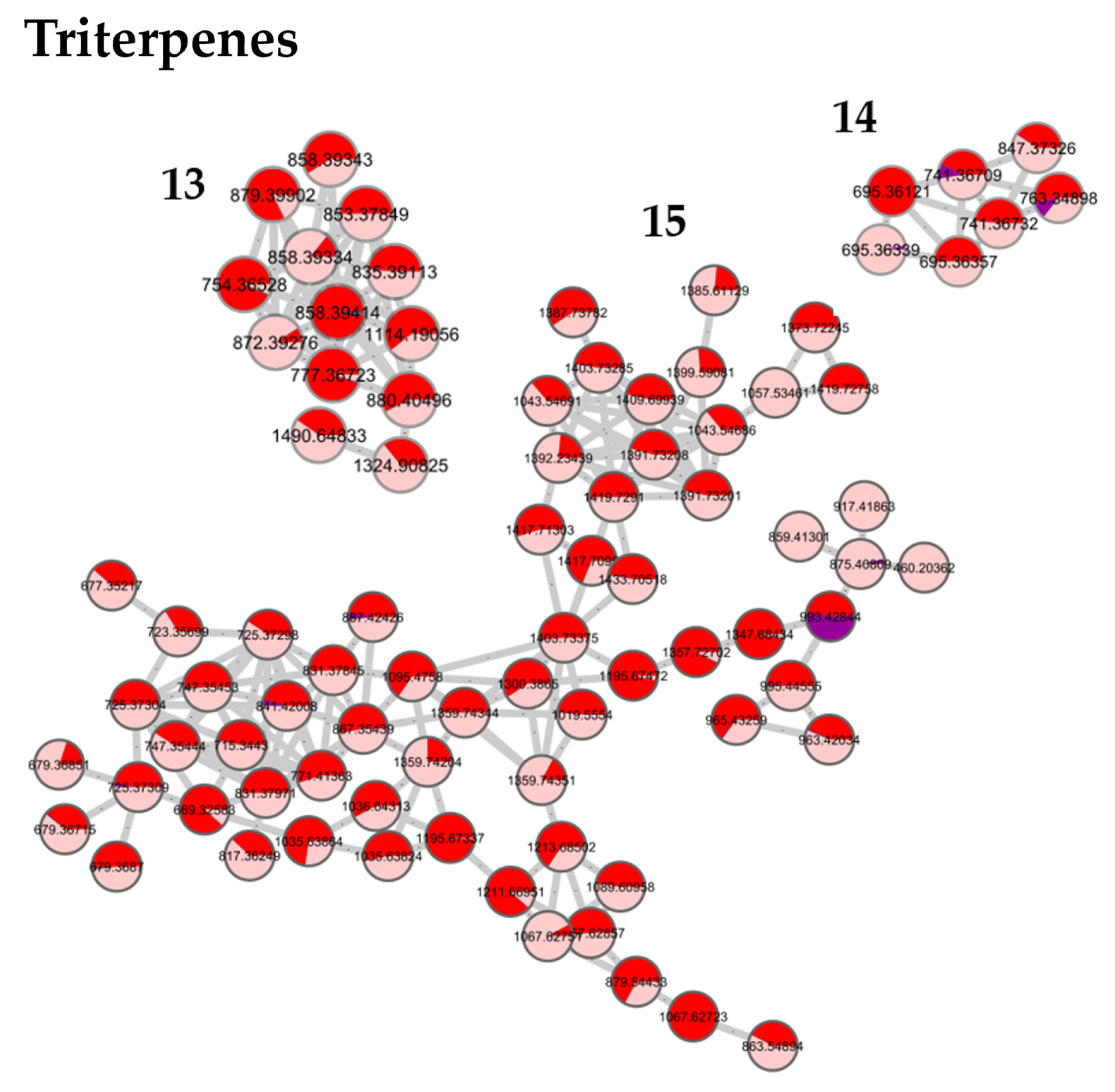
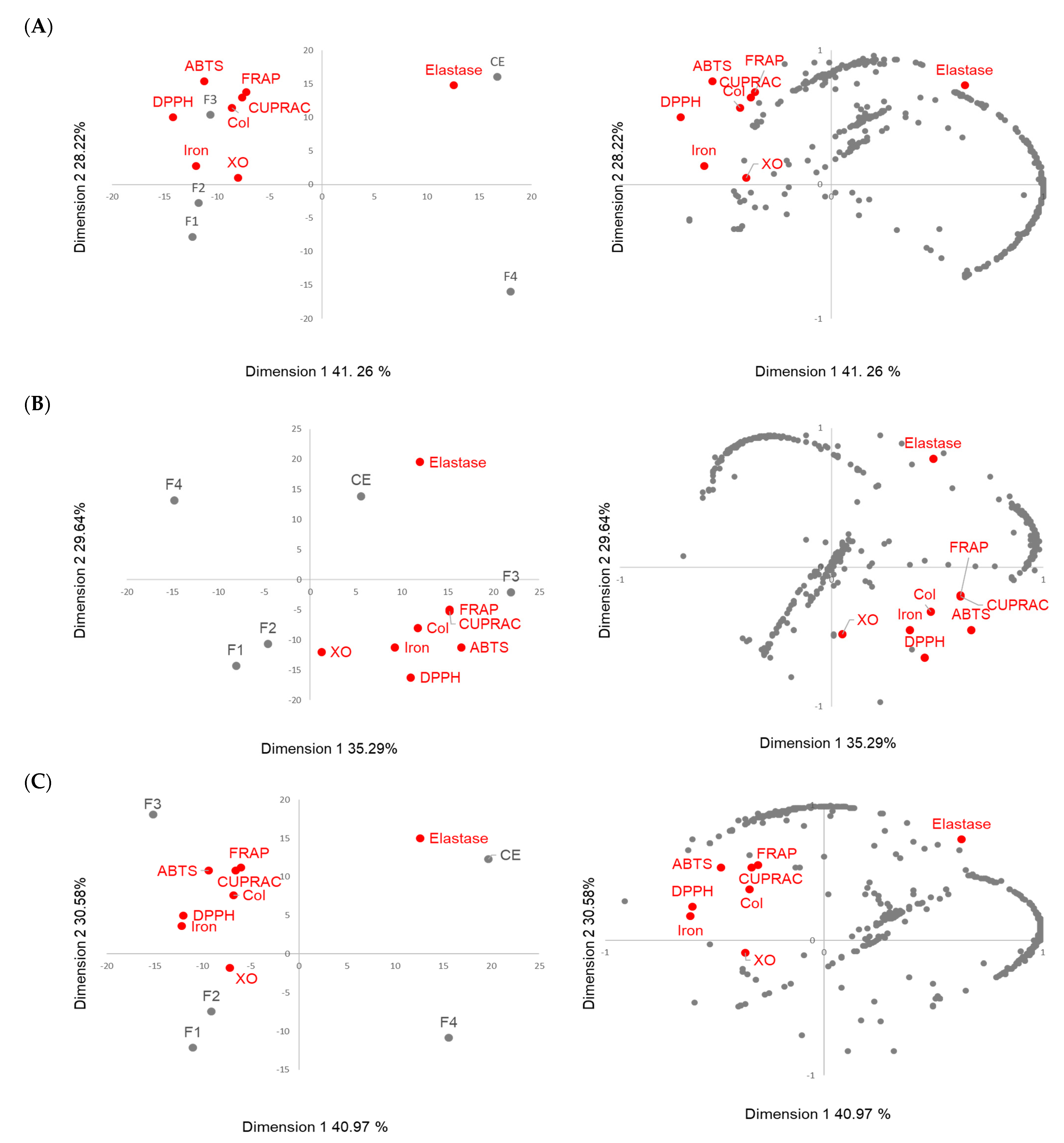
3.5. Correlation between Molecular Content and Activity
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Larrauri, J.A.; Sánchez-Moreno, C.; Rupérez, P.; Saura-Calixto, F. Free Radical Scavenging Capacity in the Aging of Selected Red Spanish Wines. J. Agric. Food Chem. 1999, 47, 1603–1606. [Google Scholar] [CrossRef] [PubMed]
- Canas, S.; Casanova, V.; Pedro Belchior, A. Antioxidant Activity and Phenolic Content of Portuguese Wine Aged Brandies. J. Food Compos. Anal. 2008, 21, 626–633. [Google Scholar] [CrossRef]
- Alonso, Á.M.; Castro, R.; Rodríguez, M.C.; Guillén, D.A.; Barroso, C.G. Study of the Antioxidant Power of Brandies and Vinegars Derived from Sherry Wines and Correlation with Their Content in Polyphenols. Food Res. Int. 2004, 37, 715–721. [Google Scholar] [CrossRef]
- Jenkins, G. Molecular Mechanisms of Skin Ageing. Mech. Ageing Dev. 2002, 123, 801–810. [Google Scholar] [CrossRef]
- Kohl, E.; Steinbauer, J.; Landthaler, M.; Szeimies, R.-M. Skin Ageing. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 873–884. [Google Scholar] [CrossRef]
- Dróżdż, P.; Pyrzynska, K. Assessment of Polyphenol Content and Antioxidant Activity of Oak Bark Extracts. Eur. J. Wood Prod. 2018, 76, 793–795. [Google Scholar] [CrossRef] [Green Version]
- Gallé, A.; Haldimann, P.; Feller, U. Photosynthetic Performance and Water Relations in Young Pubescent Oak (Quercus pubescens) Trees during Drought Stress and Recovery. New Phytol. 2007, 174, 799–810. [Google Scholar] [CrossRef]
- Haldimann, P.; Gallé, A.; Feller, U. Impact of an Exceptionally Hot Dry Summer on Photosynthetic Traits in Oak (Quercus pubescens) Leaves. Tree Physiol. 2008, 28, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plainfossé, H.; Burger, P.; Azoulay, S.; Landreau, A.; Verger-Dubois, G.; Fernandez, X. Development of a Natural Anti-Age Ingredient Based on Quercus pubescens Willd. Leaves Extract—A Case Study. Cosmetics 2018, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Nisca, A.; Ștefănescu, R.; Mocan, A.; Babotă, M.; Nicolescu, A.; Mare, A.D.; Ciurea, C.N.; Man, A.; Tanase, C. A Comparative Analysis of Polyphenol Content and Biological Potential of Quercus petraea Matt. and Q. pubescens Willd. Bark Extracts. Forests 2023, 14, 116. [Google Scholar] [CrossRef]
- Guichoux, E.; Lagache, L.; Wagner, S.; Chaumeil, P.; Léger, P.; Lepais, O.; Lepoittevin, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current Trends in Microsatellite Genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef]
- Lee, S.K.; Mbwambo, Z.H.; Chung, H.; Luyengi, L.; Gamez, E.J.; Mehta, R.G.; Kinghorn, A.D.; Pezzuto, J.M. Evaluation of the Antioxidant Potential of Natural Products. Comb. Chem. High Throughput Screen. 1998, 1, 35–46. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In Vitro Bio-Accessibility and Antioxidant Activity of Grape Polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel Total Antioxidant Capacity Index for Dietary Polyphenols and Vitamins C and E, Using Their Cupric Ion Reducing Capability in the Presence of Neocuproine: CUPRAC Method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef] [PubMed]
- Dinis, T.C.P.; Madeira, V.M.C.; Almeida, L.M. Action of Phenolic Derivatives (Acetaminophen, Salicylate, and 5-Aminosalicylate) as Inhibitors of Membrane Lipid Peroxidation and as Peroxyl Radical Scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Evaluation of Antioxidant, Antibacterial and Anti-Tyrosinase Activities of Four Macaranga Species—Science Direct. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0308814608011862 (accessed on 23 March 2023).
- Kweon, M.-H.; Hwang, H.-J.; Sung, H.-C. Identification and Antioxidant Activity of Novel Chlorogenic Acid Derivatives from Bamboo (Phyllostachys edulis). J. Agric. Food Chem. 2001, 49, 4646–4655. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with GNPS. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feature-Based Molecular Networking in the GNPS Analysis Environment|Nature Methods. Available online: https://www.nature.com/articles/s41592-020-0933-6 (accessed on 5 July 2021).
- Prida, A.; Boulet, J.; Ducousso, A.; Nepveu, G.; Puech, J. Effect of Species and Ecological Conditions on Ellagitannin Content in Oak Wood from an Even-Aged and Mixed Stand of Quercus robur L. and Quercus petraea Liebl. Ann. For. Sci. 2006, 63, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Buche, G.; Colas, C.; Fougère, L.; Destandau, E. Oak Species Quercus robur L. and Quercus petraea Liebl. Identification Based on UHPLC-HRMS/MS Molecular Networks. Metabolites 2021, 11, 684. [Google Scholar] [CrossRef]
- Buche, G.; Colas, C.; Fougère, L.; Giordanengo, T.; Destandau, E. Untargeted UHPLC-Q-TOF-HRMS Based Determination of Discrimating Compounds for Oak Species Quercus robur L. and Quercus petraea Liebl. Identification. Phytochem. Anal. 2020, 32, 660–671. [Google Scholar] [CrossRef]
- Marchal, A.; Prida, A.; Dubourdieu, D. New Approach for Differentiating Sessile and Pedunculate Oak: Development of a LC-HRMS Method To Quantitate Triterpenoids in Wood. J. Agric. Food Chem. 2016, 64, 618–626. [Google Scholar] [CrossRef]
- Marchal, A.; Dubourdieu, D. Method for Identifying the Oak Species of an Oak Wood Sample. U.S. Patent 2016/0011166 A1, 14 January 2016. [Google Scholar]
- Frank, A.M.; Bandeira, N.; Shen, Z.; Tanner, S.; Briggs, S.P.; Smith, R.D.; Pevzner, P.A. Clustering Millions of Tandem Mass Spectra. J. Proteome Res. 2007, 7, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef]
- Fraga-Corral, M.; Otero, P.; Echave, J.; Garcia-Oliveira, P.; Carpena, M.; Jarboui, A.; Nuñez-Estevez, B.; Simal-Gandara, J.; Prieto, M.A. By-Products of Agri-Food Industry as Tannin-Rich Sources: A Review of Tannins’ Biological Activities and Their Potential for Valorization. Foods 2021, 10, 137. [Google Scholar] [CrossRef]
- Centrifugal Partition Chromatography Applied to the Isolation of Oak Wood Aroma Precursors|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S0308814613005153?token=E993B2DAF29A3BBE1B6F2EB9836E50326FC688D5DA80E6036E1EB04F1BC6F141B831A98064361AB4E5DE019E76585EDB&originRegion=eu-west-1&originCreation=20210707140737 (accessed on 7 July 2021).
- Winstel, D.; Marchal, A. Lignans in Spirits: Chemical Diversity, Quantification, and Sensory Impact of (±)-Lyoniresinol. Molecules 2019, 24, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alañón, M.E.; Castro-Vázquez, L.; Díaz-Maroto, M.C.; Gordon, M.H.; Pérez-Coello, M.S. A Study of the Antioxidant Capacity of Oak Wood Used in Wine Ageing and the Correlation with Polyphenol Composition. Food Chem. 2011, 128, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- González Burgos, E.; Gómez-Serranillos, M. Terpene Compounds in Nature: A Review of Their Potential Antioxidant Activity. Curr. Med. Chem. 2012, 19, 5319–5341. [Google Scholar] [CrossRef]
- Vivas, N.; Glories, Y.; Bourgeois, G.; Vitry, C. The Heartwood Ellagitannins of Different Oaks (Quercus Sp.) and Chestnut Species (Castanea Sativa Mill.). Quantity Analysis of Red Wines Aging in Barrels. J. Des Sci. Tech. Tonnelerie 1996. [Google Scholar]
- Saucier, C.; Jourdes, M.; Glories, Y.; Quideau, S. Extraction, Detection, and Quantification of Flavano-Ellagitannins and Ethylvescalagin in a Bordeaux Red Wine Aged in Oak Barrels. J. Agric. Food Chem. 2006, 54, 7349–7354. [Google Scholar]
- Cammann, J.; Denzel, K.; Schilling, G.; Gross, G.G. Biosynthesis of Gallotannins: Beta-Glucogallin-Dependent Formation of 1,2,3,4,6-Pentagalloylglucose by Enzymatic Galloylation of 1,2,3,6-Tetragalloylglucose. Arch. Biochem. Biophys. 1989, 273, 58–63. [Google Scholar] [CrossRef]
- Dad, G.; Corbani, A.; Manitto, P.; Speranza, G.; Lunazzi, L. Lignan Glycosides from the Heartwood of European Oak Quercus Petraea. J. Nat. Prod. 2004, 52, 1327–1330. [Google Scholar] [CrossRef]
- Othón-Díaz, E.D.; Fimbres-García, J.O.; Flores-Sauceda, M.; Silva-Espinoza, B.A.; López-Martínez, L.X.; Bernal-Mercado, A.T.; Ayala-Zavala, J.F. Antioxidants in Oak (Quercus Sp.): Potential Application to Reduce Oxidative Rancidity in Foods. Antioxidants 2023, 12, 861. [Google Scholar] [CrossRef]
- Lobiuc, A.; Pavăl, N.-E.; Mangalagiu, I.I.; Gheorghiță, R.; Teliban, G.C.; Amăriucăi-Mantu, D.; Stoleru, V. Future Antimicrobials: Natural and Functionalized Phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buche, G.; Laffon, M.; Fougère, L.; Destandau, E. Evaluation and Comparison of Dermo-Cosmetic Activities of Three Oak Species by Targeting Antioxidant Metabolites and Skin Enzyme Inhibitors. Metabolites 2023, 13, 804. https://doi.org/10.3390/metabo13070804
Buche G, Laffon M, Fougère L, Destandau E. Evaluation and Comparison of Dermo-Cosmetic Activities of Three Oak Species by Targeting Antioxidant Metabolites and Skin Enzyme Inhibitors. Metabolites. 2023; 13(7):804. https://doi.org/10.3390/metabo13070804
Chicago/Turabian StyleBuche, Gaëlle, Malorie Laffon, Laëtitia Fougère, and Emilie Destandau. 2023. "Evaluation and Comparison of Dermo-Cosmetic Activities of Three Oak Species by Targeting Antioxidant Metabolites and Skin Enzyme Inhibitors" Metabolites 13, no. 7: 804. https://doi.org/10.3390/metabo13070804
APA StyleBuche, G., Laffon, M., Fougère, L., & Destandau, E. (2023). Evaluation and Comparison of Dermo-Cosmetic Activities of Three Oak Species by Targeting Antioxidant Metabolites and Skin Enzyme Inhibitors. Metabolites, 13(7), 804. https://doi.org/10.3390/metabo13070804