Untargeted Metabolomic Analysis of Rat Neuroblastoma Cells as a Model System to Study the Biochemical Effects of the Acute Administration of Methamphetamine



Abstract
:1. Introduction
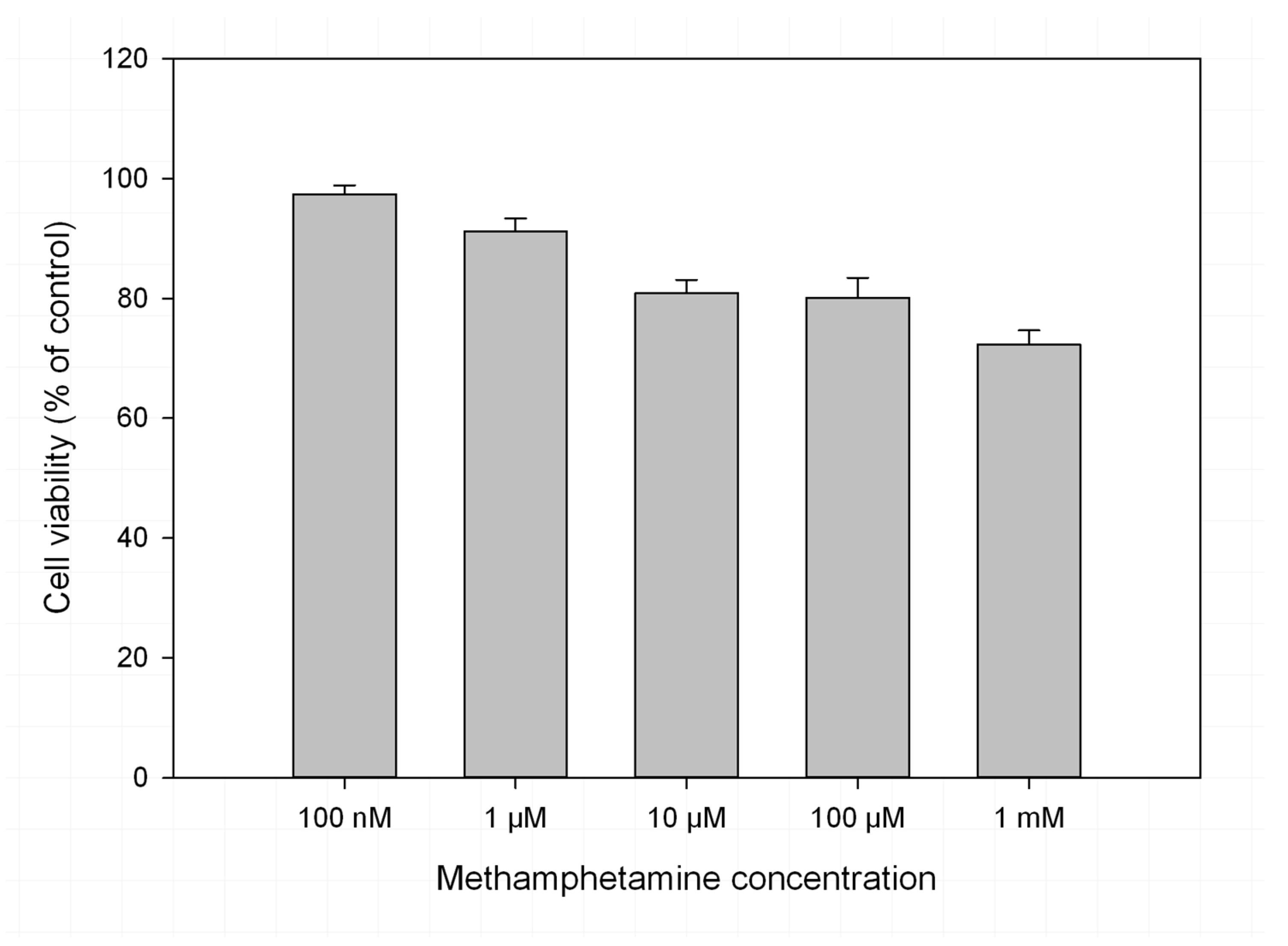
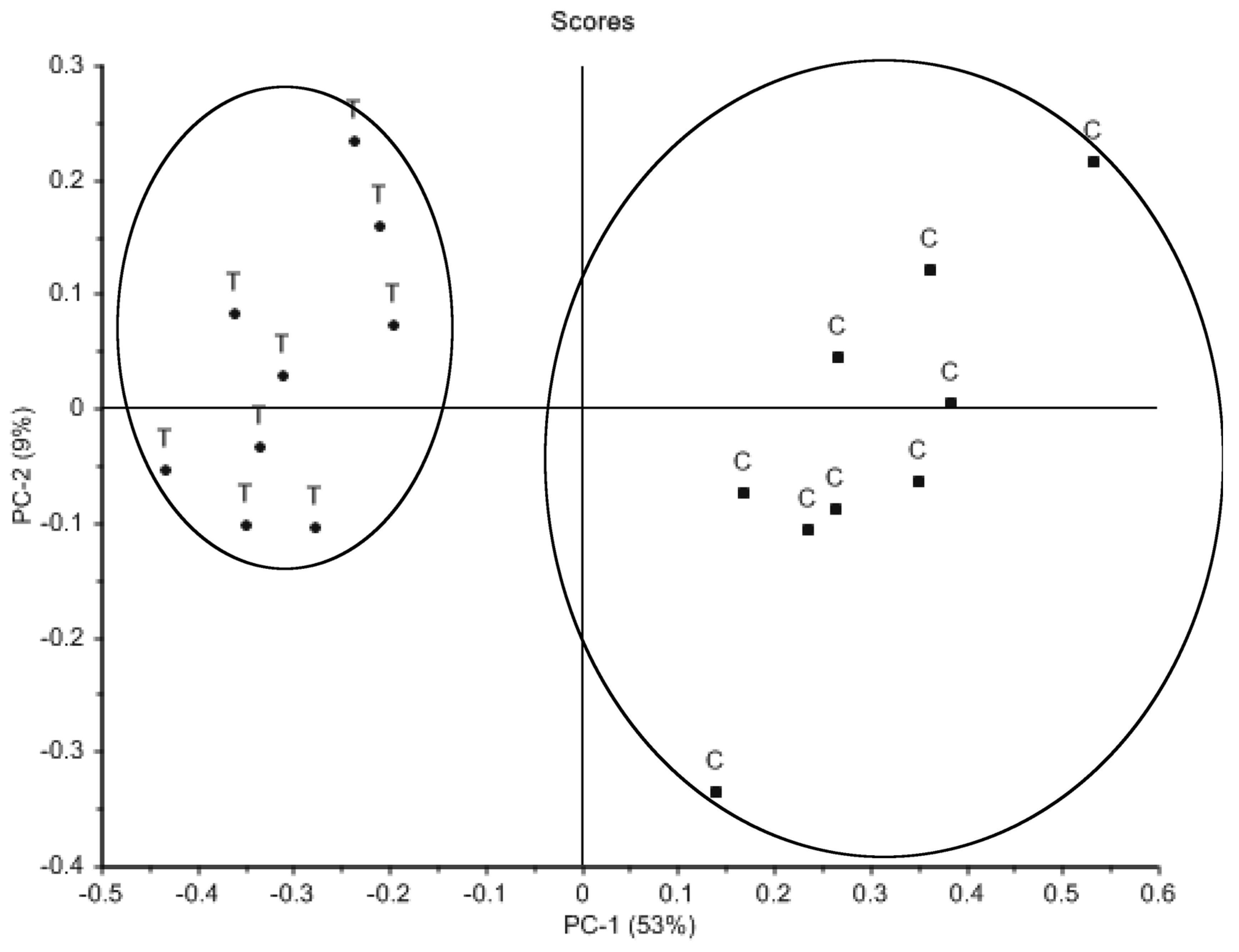
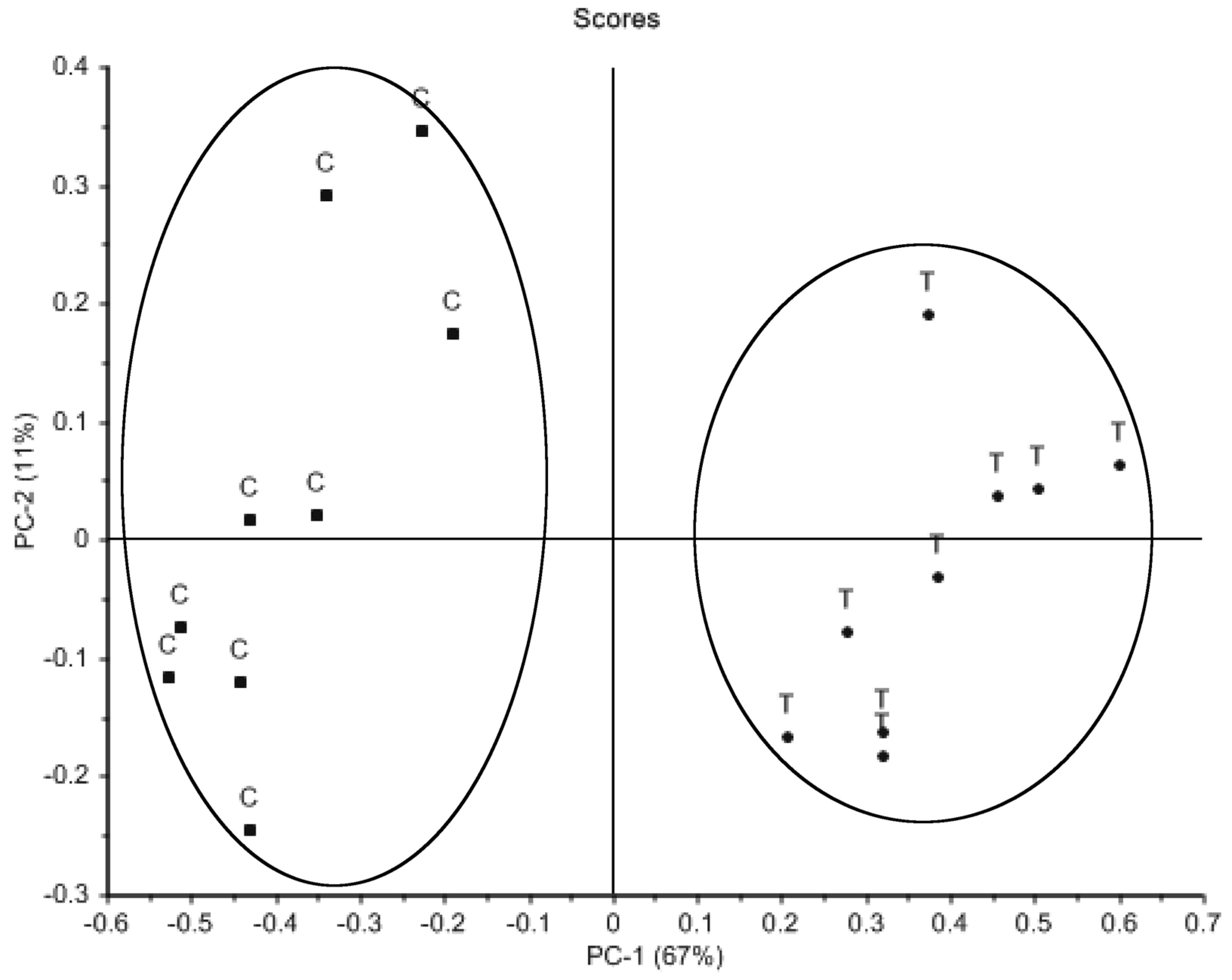
2. Results
2.1. Amino Acids
2.2. Other Metabolites
3. Discussion
3.1. Amino Acids
3.2. Energy Metabolism
3.3. Conclusions
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Sample Preparation
4.4. Analysis
4.5. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Australian Institute of Health and Welfare (AIHW). 2010 National Drug Strategy Household Survey Report; Drug Statistics Series; AIHW: Canberra, Australia, 2011; Volume 25. [Google Scholar]
- Zweben, J.E.; Cohen, J.B.; Christian, D.; Galloway, G.P.; Salinardi, M.; Parent, D.; Iguchi, M.; Methamphetamine Treatment Project. Psychiatric symptoms in methamphetamine users. Am. J. Addict. 2004, 13, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Darke, S.; Kaye, S.; McKetin, R.; Duflou, J. Major physical and psychological harms of methamphetamine use. Drug Alcohol Rev. 2008, 27, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Homer, B.D.; Solomon, T.M.; Moeller, R.W.; Mascia, A.; DeRaleau, L.; Halkitis, P.N. Methamphetamine abuse and impairment of social functioning: A review of the underlying neurophysiological causes and behavioral implications. Psychol. Bull. 2008, 134, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.C.; Woods, S.P.; Matt, G.E.; Meyer, R.A.; Heaton, R.K.; Atkinson, J.H.; Grant, I. Neurocognitive effects of methamphetamine: A critical review and meta-analysis. Neuropsychol. Rev. 2007, 17, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Barua, R.; Ambrose, J. Methamphetamine-associated acute left ventricular dysfunction: A variant of stress-induced cardiomyopathy. Cardiology 2008, 109, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Zolkowska, D.; Rothman, R.B.; Baumann, M.H. Amphetamine analogs increase plasma serotonin: Implications for cardiac and pulmonary disease. J. Pharmacol. Exp. Ther. 2006, 318, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Sonders, M.S.; Poulsen, N.W.; Galli, A. Mechanisms of neurotransmitter release by amphetamines: A review. Prog. Neurobiol. 2005, 75, 406–433. [Google Scholar] [CrossRef] [PubMed]
- Krasnova, I.N.; Cadet, J.L. Methamphetamine toxicity and messengers of death. Brain Res. Rev. 2009, 60, 379–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, J.F.; Cho, A.K. Chemical release of dopamine from striatal homogenates: Evidence for an exchange diffusion model. J. Pharmacol. Exp. Ther. 1979, 208, 203–209. [Google Scholar] [PubMed]
- Cadet, J.L.; Brannock, C. Free radicals and the pathobiology of brain dopamine systems. Neurochem. Int. 1998, 32, 117–131. [Google Scholar] [CrossRef]
- Harold, C.; Wallace, T.; Friedman, R.; Gudelsky, G.; Yamamoto, B. Methamphetamine selectively alters brain glutathione. Eur. J. Pharmacol. 2000, 400, 99–102. [Google Scholar] [CrossRef]
- Gluck, M.R.; Moy, L.Y.; Jayatilleke, E.; Hogan, K.A.; Manzino, L.; Sonsalla, P.K. Parallel increases in lipid and protein oxidative markers in several mouse brain regions after methamphetamine treatment. J. Neurochem. 2001, 79, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Lee, Y.C.; Huang, C.L.; Liu, H.K.; Liao, W.C.; Lai, W.L.; Lin, Y.R.; Huang, N.K. Methamphetamine downregulates peroxiredoxins in rat pheochromocytoma cells. Biochem. Biophys. Res. Commun. 2007, 354, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Iwazaki, T.; McGregor, I.S.; Matsumoto, I. Protein expression profile in the striatum of acute methamphetamine-treated rats. Brain Res. 2006, 1097, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, H.; Qiu, P.; Luo, H. Proteomic profiling of proteins associated with methamphetamine-induced neurotoxicity in different regions of rat brain. Neurochem. Int. 2008, 52, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, P.S.; Tong, J.; Yazdanpanah, M.; Liu, P.P.; Kalasinsky, K.S.; Kish, S.J. Levels of 4-hydroxynonenal and malondialdehyde are increased in brain of human chronic users of methamphetamine. J. Pharmacol. Exp. Ther. 2006, 319, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Jeng, W.; Ramkissoon, A.; Parman, T.; Wells, P.G. Prostaglandin H synthase-catalyzed bioactivation of amphetamines to free radical intermediates that cause CNS regional DNA oxidation and nerve terminal degeneration. FASEB J. 2006, 20, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Mark, K.A.; Soghomonian, J.J.; Yamamoto, B.K. High-dose methamphetamine acutely activates the striatonigral pathway to increase striatal glutamate and mediate long-term dopamine toxicity. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 11449–11456. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Rothman, S.M. The role of glutamate neurotoxicity in hypoxic-ischemic neuronal death. Annu. Rev. Neurosci. 1990, 13, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Gwag, B.J.; Koh, J.Y.; DeMaro, J.A.; Ying, H.S.; Jacquin, M.; Choi, D.W. Slowly triggered excitotoxicity occurs by necrosis in cortical cultures. Neuroscience 1997, 77, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Sonsalla, P.K.; Albers, D.S.; Zeevalk, G.D. Role of glutamate in neurodegeneration of dopamine neurons in several animal models of Parkinsonism. Amino Acids 1998, 14, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Chang, L.; Wang, G.J.; Fowler, J.S.; Franceschi, D.; Sedler, M.J.; Gatley, S.J.; Hitzemann, R.; Ding, Y.S.; Wong, C.; et al. Higher cortical and lower subcortical metabolism in detoxified methamphetamine abusers. Am. J. Psychiatry 2001, 158, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Berman, S.M.; Voytek, B.; Mandelkern, M.A.; Hassid, B.D.; Isaacson, A.; Monterosso, J.; Miotto, K.; Ling, W.; London, E.D. Changes in cerebral glucose metabolism during early abstinence from chronic methamphetamine abuse. Mol. Psychiatry 2008, 13, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Funada, M.; Zhou, X.; Satoh, M.; Wada, K. Profiling of methamphetamine-induced modifications of gene expression patterns in the mouse brain. Ann. N. Y. Acad. Sci. 2004, 1025, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.; Jayanthi, S.; McCoy, M.T.; Brannock, C.; Ladenheim, B.; Garrett, T.; Lehrmann, E.; Becker, K.; Cadet, J.L. Methamphetamine causes differential alterations in gene expression and patterns of histone acetylation/hypoacetylation in the rat nucleus accumbens. PLoS ONE 2012, 7, e34236. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.L.; Jayanthi, S.; McCoy, M.T.; Ladenheim, B.; Saint-Preux, F.; Lehrmann, E.; De, S.; Becker, K.; Brannock, C. Genome-wide profiling identifies a subset of methamphetamine (METH)-induced genes associated with METH-induced increased H4K5Ac binding in the rat striatum. BMC Genom. 2013, 14, 545–563. [Google Scholar] [CrossRef] [PubMed]
- Hutschenreuther, A.; Kiontke, A.; Birkenmeier, G.; Birkemeyer, C. Comparison of extraction conditions and normalization approaches for cellular metabolomics of adherent growing cells with GC-MS. Anal. Methods 2012, 4, 1953–1963. [Google Scholar] [CrossRef]
- Zaitsu, K.; Miyawaki, I.; Bando, K.; Horie, H.; Shima, N.; Katagi, M.; Tatsuno, M.; Bamba, T.; Sato, T.; Ishii, A.; et al. Metabolic profiling of urine and blood plasma in rat models of drug addiction on the basis of morphine, methamphetamine, and cocaine-induced conditioned place preference. Anal. Bioanal. Chem. 2014, 406, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Kim, S.P.; Hwang, S.; Hwang, J.; Yang, C.H.; Lee, S. Metabolic characterization in urine and hair from a rat model of methamphetamine self-administration using LC-QTOF-MS-based metabolomics. Metabolomics 2017, 13, 119. [Google Scholar] [CrossRef]
- Zheng, T.; Liu, L.; Shi, J.; Yu, X.; Xiao, W.; Sun, R.; Zhou, Y.; Aa, J.; Wang, G. The metabolic impact of methamphetamine on the systemic metabolism of rats and potential markers of methamphetamine abuse. Mol. Biosyst. 2014, 10, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Choucha Snouber, L.; Bunescu, A.; Naudot, M.; Legallais, C.; Brochot, C.; Dumas, M.E.; Elena-Herrmann, B.; Leclerc, E. Metabolomics-on-a-chip of hepatotoxicity induced by anticancer drug flutamide and its active metabolite hydroxyflutamide using HepG2/C3a microfluidic biochips. Toxicol. Sci. 2013, 132, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Van den Hof, W.F.; Ruiz-Aracama, A.; Van Summeren, A.; Jennen, D.G.; Gaj, S.; Coonen, M.L.; Brauers, K.; Wodzig, W.K.; van Delft, J.H.; Kleinjans, J.C. Integrating multiple omics to unravel mechanisms of cyclosporin a induced hepatotoxicity in vitro. Toxicol. In Vitro 2015, 29, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, E.; Morath, S.; Eskes, C.; Linge, J.; Rappsilber, J.; Honegger, P.; Hartung, T.; Coecke, S. A novel in vitro metabolomics approach for neurotoxicity testing, proof of principle for methyl mercury chloride and caffeine. Neurotoxicology 2008, 29, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hayton, S.; Maker, G.L.; Mullaney, I.; Trengove, R.D. Untargeted metabolomics of neuronal cell culture: A model system for the toxicity testing of insecticide chemical exposure. J. Appl. Toxicol. 2017, 37, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, T.; Daneshian, M.; Kamp, H.; Bois, F.; Clench, M.; Coen, M.; Donley, B.; Fischer, S.; Ekman, D.; Fabian, E.; et al. Metabolomics in toxicology and preclinical research. ALTEX 2013, 30, 209–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiamulera, C.; Padovani, L.; Corsi, M. Drug discovery for the treatment of substance use disorders: Novel targets, repurposing, and the need for new paradigms. Curr. Opin. Pharmacol. 2017, 35, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.H.; Jiang, Z.L.; Chen, Z.Q.; Li, X.; Peng, L.L. Neuroprotective effect of L-serine against temporary cerebral ischemia in rats. J. Neurosci. Res. 2010, 88, 2035–2045. [Google Scholar] [CrossRef] [PubMed]
- Naderi Maralani, M.; Movahedian, A.; Haghjooy Javanmard, S. Antioxidant and cytoprotective effects of L-serine on human endothelial cells. Res. Pharm. Sci. 2012, 7, 209–215. [Google Scholar]
- Ibegbu, A.; Fyfe, L.; McBean, D.; Mullaney, I. The effects of hypoxia and opioid receptor agonist treatment in cortical b50 neuronal cells in culture. J. Biol. Environ. Sci. 2012, 6, 219–231. [Google Scholar]
- Nagamatsu, S.; Sawa, H.; Kamada, K.; Nakamichi, Y.; Yoshimoto, K.; Hoshino, T. Neuron-specific glucose transporter (NSGT): CNS distribution of GLUT3 rat glucose transporter (RGT3) in rat central neurons. FEBS Lett. 1993, 334, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Abdul Muneer, P.M.; Alikunju, S.; Szlachetka, A.M.; Haorah, J. Methamphetamine inhibits the glucose uptake by human neurons and astrocytes: Stabilization by acetyl-l-carnitine. PLoS ONE 2011, 6, e19258. [Google Scholar] [CrossRef] [PubMed]
- LaVoie, M.J.; Hastings, T.G. Dopamine quinone formation and protein modification associated with the striatal neurotoxicity of methamphetamine: Evidence against a role for extracellular dopamine. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 1484–1491. [Google Scholar] [CrossRef]
- Njalsson, R. Glutathione synthetase deficiency. Cell. Mol. Life Sci. 2005, 62, 1938–1945. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.R.; Silva, C.G.; Ruschel, C.; Helegda, C.; Wyse, A.T.; Wannmacher, C.M.; Wajner, M.; Dutra-Filho, C.S. l-pyroglutamic acid inhibits energy production and lipid synthesis in cerebral cortex of young rats in vitro. Neurochem. Res. 2001, 26, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Sinchai, T.; Plasen, S.; Sanvarinda, Y.; Jaisin, Y.; Govitrapong, P.; Morales, N.P.; Ratanachamnong, P.; Plasen, D. Caffeine potentiates methamphetamine-induced toxicity both in vitro and in vivo. Neurosci. Lett. 2011, 502, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Hu, Z.; Hu, C.; Bu, Q.; Yan, G.; Deng, P.; Lv, L.; Wu, D.; Deng, Y.; Zhao, J.; et al. Taurine protects methamphetamine-induced developmental angiogenesis defect through antioxidant mechanism. Toxicol. Appl. Pharmacol. 2012, 260, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.S.; Nogueira, T.B.; Costa, V.M.; Branco, P.S.; Ferreira, L.M.; Fernandes, E.; Bastos, M.L.; Meisel, A.; Carvalho, F.; Capela, J.P. Neurotoxicity of “ecstasy” and its metabolites in human dopaminergic differentiated SH-SY5Y cells. Toxicol. Lett. 2013, 216, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Xie, W.B.; Qiao, D.; Qiu, P.; Huang, E.; Li, B.; Chen, C.; Liu, C.; Wang, Q.; Lin, Z.; et al. Caspase-11 plays an essential role in methamphetamine-induced dopaminergic neuron apoptosis. Toxicol. Sci. 2015, 145, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Kronthaler, J.; Gstraunthaler, G.; Heel, C. Optimizing high-throughput metabolomic biomarker screening: A study of quenching solutions to freeze intracellular metabolism in CHO cells. OMICS A J. Integr. Biol. 2012, 16, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Abbiss, H.; Maker, G.L.; Gummer, J.; Sharman, M.J.; Phillips, J.K.; Boyce, M.; Trengove, R.D. Development of a non-targeted metabolomics method to investigate urine in a rat model of polycystic kidney disease. Nephrology 2012, 17, 104–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gummer, J.P.; Krill, C.; Du Fall, L.; Waters, O.D.; Trengove, R.D.; Oliver, R.P.; Solomon, P.S. Metabolomics protocols for filamentous fungi. Methods Mol. Biol. 2012, 835, 237–254. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Metabolite | Cells (Fold Change) | Medium (Fold Change) |
|---|---|---|
| Amino Acids | ||
| N-Acetylglutamate | 4.7 ** | 1.4 * |
| l-Alanine | 0.8 | 1.4 ** |
| l-Arginine | 1.1 | 1.3 * |
| l-Aspartate | 8.0 ** | 2.6 ** |
| l-Glutamate | 3.3 ** | 2.1 ** |
| Glycine | - | 1.1 |
| l-Isoleucine | 0.9 | 1.7 ** |
| l-Leucine | 1.4 | 1.1 |
| l-Methionine | 3.5 ** | 2.5 ** |
| l-Phenylalanine | 0.5 ** | 1.9 ** |
| l-Proline | 0.7 ** | 1.1 |
| Pyroglutamate | 1.1 | 1.7 ** |
| l-Serine | 0.3 ** | 3.0 ** |
| l-Threonine | 0.9 | 1.3 * |
| l-Tryptophan | - | 1.7 ** |
| l-Tyrosine | - | 1.7 ** |
| l-Valine | - | 1.4 ** |
| Carbohydrates | ||
| Arabitol | - | - |
| Fructose | 1.2 | - |
| Galactose | 1.1 ** | 1.2 ** |
| Glucose | 2.4 | - |
| Mannose | 0.6 | - |
| Unidentified carbohydrate 1 | 1.6 ** | 1.2 |
| Unidentified carbohydrate 2 | 1.3 | 0.9 |
| Unidentified carbohydrate 3 | 1.3 | 1.1 |
| Other metabolites | ||
| Cholesterol | 0.6 ** | not detected |
| Erythronate | 0.9 | 1.1 |
| Fumarate | 2.1 ** | 1.1 |
| GABA | 0.2 ** | not detected |
| Glycerol | 0.2 ** | - |
| Hexadecanoate | - | 1.1 |
| myo-Inositol | 1.3 ** | - |
| Octadecanoate | 1.3 * | 0.8 |
| Succinate | 1.6 * | - |
| Tryptamine | 2.1 ** | not detected |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maker, G.L.; Green, T.; Mullaney, I.; Trengove, R.D. Untargeted Metabolomic Analysis of Rat Neuroblastoma Cells as a Model System to Study the Biochemical Effects of the Acute Administration of Methamphetamine. Metabolites 2018, 8, 38. https://doi.org/10.3390/metabo8020038
Maker GL, Green T, Mullaney I, Trengove RD. Untargeted Metabolomic Analysis of Rat Neuroblastoma Cells as a Model System to Study the Biochemical Effects of the Acute Administration of Methamphetamine. Metabolites. 2018; 8(2):38. https://doi.org/10.3390/metabo8020038
Chicago/Turabian StyleMaker, Garth L., Tobias Green, Ian Mullaney, and Robert D. Trengove. 2018. "Untargeted Metabolomic Analysis of Rat Neuroblastoma Cells as a Model System to Study the Biochemical Effects of the Acute Administration of Methamphetamine" Metabolites 8, no. 2: 38. https://doi.org/10.3390/metabo8020038
APA StyleMaker, G. L., Green, T., Mullaney, I., & Trengove, R. D. (2018). Untargeted Metabolomic Analysis of Rat Neuroblastoma Cells as a Model System to Study the Biochemical Effects of the Acute Administration of Methamphetamine. Metabolites, 8(2), 38. https://doi.org/10.3390/metabo8020038