Metabolomics Discovers Early-Response Metabolic Biomarkers that Can Predict Chronic Reproductive Fitness in Individual Daphnia magna


Abstract
:1. Introduction
2. Results
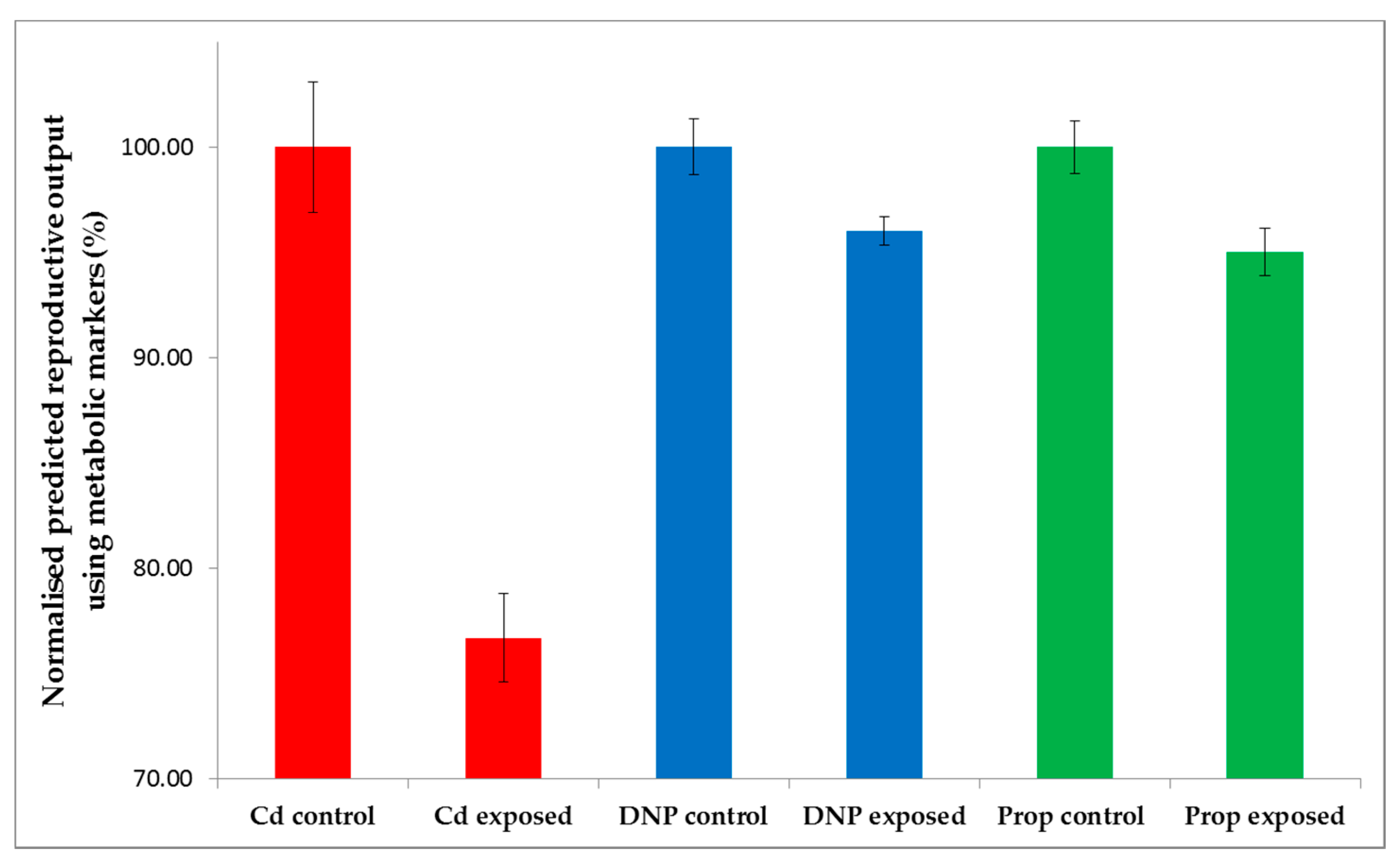
2.1. Chronic Chemical Exposures Reduced Daphnia Reproductive Fitness
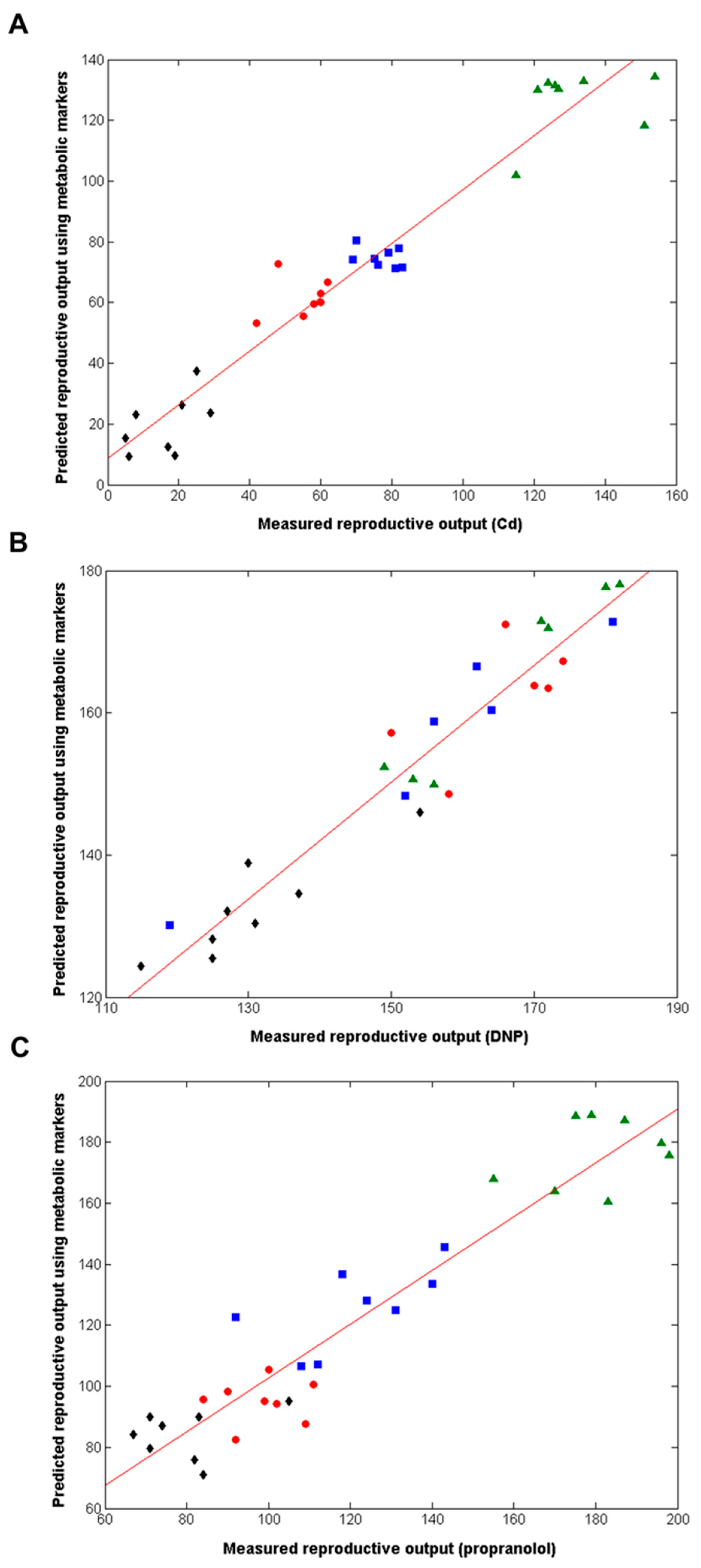
2.2. Metabolomics Discovers Biomarker Signatures that Predict Daphnia Reproductive Fitness Following Chronic Exposures to Individual Chemicals
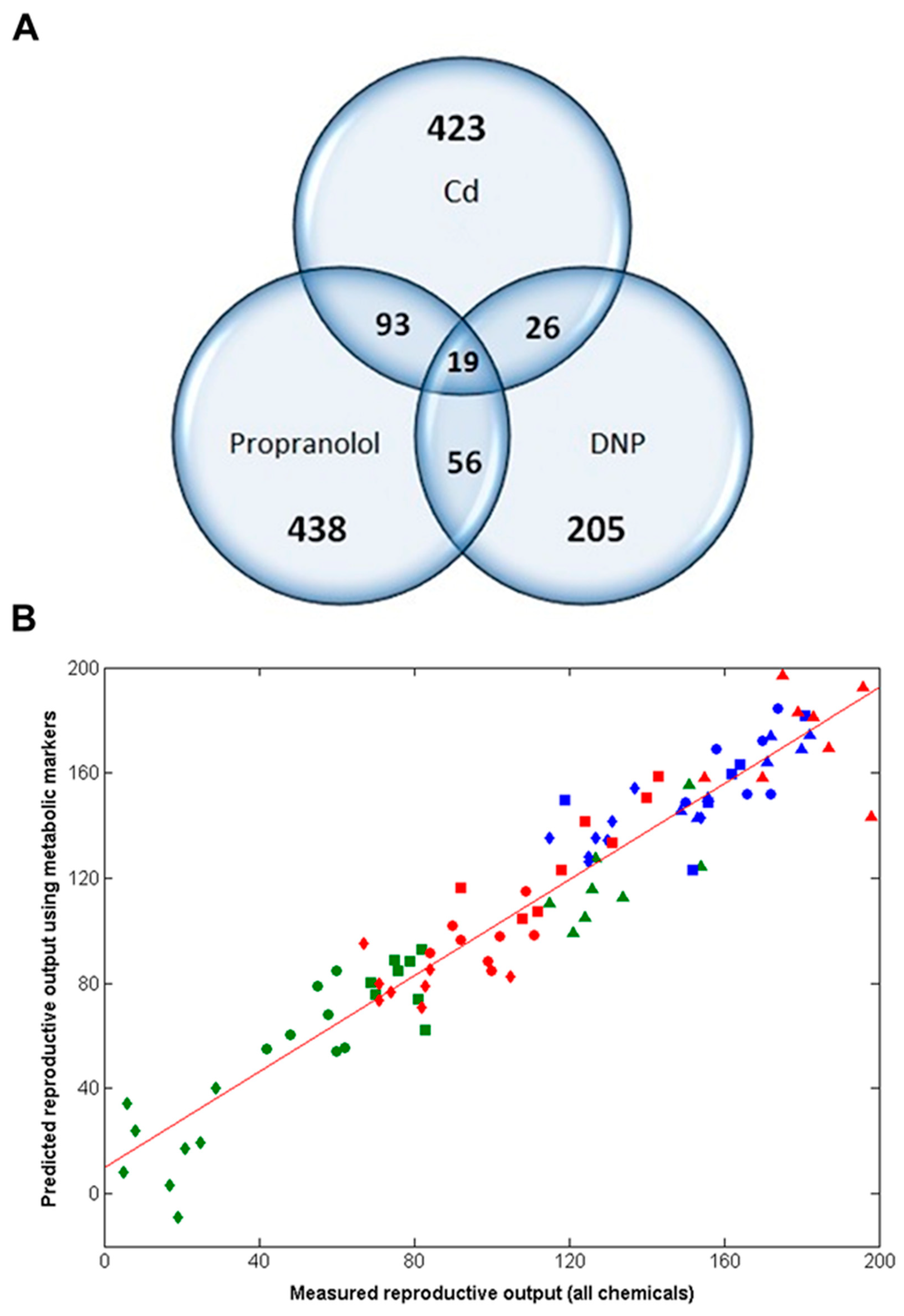
2.3. Metabolomics Discovers a 49-Biomarker Signature that Predicts Reproductive Fitness Following Chronic Exposure to All Three Chemicals
2.4. Chronic Reproductive Fitness Can Be Predicted from Early-Response Metabolic Measurements
3. Discussion
4. Materials and Methods
4.1. Daphnia Magna Culturing and Chemical Exposures
4.2. Metabolite Extraction and Mass Spectrometry Metabolomics
4.3. Statistical Analysis of Reproductive Output
4.4. Statistical Analysis of Metabolomics Datasets from 21-Day Exposures
4.5. Statistical Analysis of Metabolomics Datasets from 24-h Exposures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grandjean, P.; Ballanger, M. Calculation of the disease burden associated with environmental chemical exposures: Application of toxicological information in health economic estimation. Environ. Health 2017, 16, 123. [Google Scholar] [CrossRef] [PubMed]
- Darnerud, P.O. Toxic effects of brominated flame retardants in man and wildlife. Environ. Int. 2003, 29, 841–853. [Google Scholar] [CrossRef]
- Fry, M.; Toone, C.K. DDT-Induced feminization of gull embryos. Science 1981, 213, 922–924. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.M.; Ross-Innes, C.S.; Fitzgerald, R.C. The ‘-omics’ revolution and oesophageal adenocarcinoma. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Erickson, A.R.; Cantarel, B.L.; Lamendella, R.; Darzi, Y.; Mongodin, E.F.; Pan, C.; Shah, M.; Halfvarson, J.; Tysk, C.; Henrissat, B.; et al. Integrated metagenomics/metaproteomics reveals human host-microbiota signatures of Crohn’s disease. PLoS ONE 2012, 7, e49138. [Google Scholar] [CrossRef] [PubMed]
- Organization for Economic Cooperation and Development (OECD). OECD Guidelines for Testing of Chemicals, Section 4, Test No. 408: Repeated Dose 90-Day Oral Toxicity Study in Rodents; OECD Publishing: Paris, France, 1998. [Google Scholar]
- Organization for Economic Cooperation and Development (OECD). OECD Guidelines for Testing of Chemicals, Section 2, Test No. 202: Daphnia sp. Acute Immobilisation Test; OECD Publishing: Paris, France, 2004. [Google Scholar]
- Organization for Economic Cooperation and Development (OECD). OECD Guidelines for Testing of Chemicals, Section 2, Test No. 211: Daphnia Magna Reproduction Test; OECD Publishing: Paris, France, 2012. [Google Scholar]
- Pennak, R.W. Fresh-Water Invertebrates of the United States, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 1978; ISBN 978-0471042495. [Google Scholar]
- Rockwood, L.L.; Witt, J.W. Introduction to Population Ecology, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; ISBN 978-1-118-94757-9. [Google Scholar]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Organization for Economic Cooperation and Development (OECD). OECD Guidelines for Testing of Chemicals, Section 2, Test No. 210: Fish, Early-life Stage Toxicity Test; OECD Publishing: Paris, France, 2013. [Google Scholar]
- Ankley, G.T. Genomics in Regulatory Ecotoxicology: Applications and Challenges; In Collaboration with the Society of Environmental Toxicology and Chemistry; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Van Ravenzwaay, B.; Herold, M.; Kamp, H.; Kapp, M.D.; Fabian, E.; Looser, R.; Krennrich, G.; Mellert, W.; Prokoudine, A.; Strauss, V.; et al. Metabolomics: A tool for early detection of toxicological effects and an opportunity for biology based grouping of chemicals—From QSAR to QBAR. Mutat. Res. 2012, 746, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Ankley, G.T.; Jensen, K.M.; Makynen, E.A.; Kahl, M.D.; Korte, J.J.; Hornung, M.W.; Henry, T.R.; Denny, J.S.; Leino, R.L.; Wilson, V.S.; et al. Effects of the androgenic growth promoter 17-β-trenbolone on fecundity and reproductive endocrinology of the fathead minnow. Environ. Toxicol. Chem. 2003, 22, 1350–1360. [Google Scholar] [CrossRef] [PubMed]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Burden, N.; Mahoney, C.; Müller, B.P.; Terry, C.; Westmoreland, C.; Kimber, I. Aligning the 3Rs with new paradigms in the safety assessment of chemicals. Toxicology 2015, 330, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Tollefsen, K.E.; Scholz, S.; Cronin, M.T.; Edwards, S.W.; de Knecht, J.; Crofton, K.; Garcia-Reyero, N.; Hartung, T.; Worth, A.; Patlewicz, G. Applying Adverse Outcome Pathways (AOPs) to support Integrated Approaches to Testing and Assessment (IATA). Regul. Toxicol. Pharm. 2014, 70, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Forbes, V.E.; Palmqvist, A.; Bach, L. The use and misuse of biomarkers in ecotoxicology. Environ. Toxicol. Chem. 2006, 25, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; White, T.A.; Viant, M.R. Defining the Baseline and Oxidant Perturbed Lipidomic Profiles of Daphnia magna. Metabolites 2017, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, T.; Daneshian, M.; Kamp, H.; Bois, F.Y.; Clench, M.R.; Coen, M.; Donley, B.; Fischer, S.M.; Ekman, D.R.; Fabian, E.; et al. Metabolomics in toxicology and preclinical research. Altex-Altern. Anim. Exp. 2013, 30, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Robertson, D.G.; Watkins, P.B.; Reily, M.D. Metabolomics in toxicology: Preclinical and clinical applications. Toxicol. Sci. 2011, 120, S146–S170. [Google Scholar] [CrossRef] [PubMed]
- Viant, M.R.; Sommer, U. Mass spectrometry based environmental metabolomics: A primer and review. Metabolomics 2013, 9, S144–S158. [Google Scholar] [CrossRef]
- Van Ravenzwaay, B.; Sperber, S.; Lemke, O.; Faulhammer, F.; Kamp, H.; Mellert, W.; Strauss, V.; Strigun, A.; Peter, E.; Spitzer, M.; et al. Metabolomics as read-across tool: A case study with phenoxy herbicides. Regul. Toxicol. Pharmacol. 2016, 81, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.S.; Clewell, H.J.; Allen, B.C.; Wesselkamper, S.C.; Wang, N.C.Y.; Lambert, J.C.; Hess-Wislon, J.K.; Zhao, Q.J.; Anderson, M.E. Application of transcriptional benchmark dose values in quantitative cancer and noncancer risk assessment. Toxicol. Sci. 2011, 120, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Farmahin, R.; Williams, A.; Kuo, B.; Chepelev, N.L.; Thomas, R.S.; Barton-Maclaren, T.S.; Curran, I.H.; Nong, A.; Wade, M.G.; Yauk, C.L. Recommended approaches in the application of toxicogenomics to derive points of departure for chemical risk assessment. Arch. Toxicol. 2017, 91, 2045–2065. [Google Scholar] [CrossRef] [PubMed]
- Martyniuk, C.J.; Griffitth, R.J.; Denslow, N.D. Omics and environmental science. Environ. Toxicol. Chem. 2011, 30, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Weber, R.J.M.; White, T.A.; Viant, M.R. Discriminating between different acute chemical toxicities via changes in the daphnid metabolome. Toxicol. Sci. 2010, 118, 307–317. [Google Scholar] [CrossRef] [PubMed]
- De Coen, W.M.; Janssen, C.R. A multivariate biomarker-based model predicting population-level responses of Daphnia magna. Environ. Toxicol. Chem. 2003, 22, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.; Staff, F.J.; Widdows, J.; Compton, R.M.; Falciani, F.; Viant, M.R. Discovery of metabolic signatures for predicting whole organism toxicology. Toxicol. Sci. 2010, 115, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Kirwan, J.A.; Johnson, C.; Yan, N.D.; Viant, M.R.; Gunn, J.M.; McGeer, J.C. Predicting chronic copper and nickel reproductive toxicity to Daphnia pulex-pulicaria from whole animal metabolic profiles. Environ. Pollut. 2016, 212, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.R.; Pfrender, M.E.; Eads, B.D.; Klaper, R.; Callaghan, A.; Sibly, R.M.; Colson, I.; Jansen, B.; Gilbert, D.; Colbourne, J.K. Daphnia as an emerging model for toxicological genomics. In Comparative Toxicogenomics; Hogstrand, C., Kille, P., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2008; pp. 165–219. [Google Scholar]
- Lankadurai, B.P.; Nagato, E.G.; Sinmpson, M.J. Environmental metabolomics: An emerging approach to study organism responses to environmental stressors. Environ. Rev. 2013, 21, 180–205. [Google Scholar] [CrossRef]
- Kovacevic, V.; Simpson, A.J.; Simpson, M.J. 1H NMR-based metabolomics of Daphnia magna responses after sub-lethal exposure to triclosan, carbamazepine and ibuprofen. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 19, 199–210. [Google Scholar]
- Scanlan, L.D.; Loguinov, A.V.; Teng, Q.; Antczak, P.; Dailey, K.P.; Nowinski, D.T.; Kornbluh, J.; Lin, X.X.; Lachenauer, E.; Arai, A.; et al. Gene transcription, metabolite and lipid profiling in eco-indicator Daphnia magna indicate diverse mechanisms of toxicity by legacy and emerging flame retardants. Environ. Sci. Technol. 2015, 49, 7400–7410. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.K.; Perkins, E.J.; Habib, T.; Sims, J.G.; Chappell, P.; Escalon, B.L.; Wilbanks, M.; Garcia-Reyero, N. The Good, The Bad, and The Toxic: Approaching hormesis in Daphnia magna exposed to an energetic compound. Environ. Sci. Technol. 2013, 47, 9424–9433. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Weber, R.J.M.; Southam, A.D.; Payne, T.G.; Hrydziuszko, O.; Arvanitis, T.N.; Viant, M.R. A new approach to toxicity testing in Daphnia magna: Application of high throughput FT-ICR mass spectrometry metabolomics. Metabolomics 2009, 5, 44–58. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillard, J.; Frenkel, J.; Devos, V.; Sabbe, K.; Paul, C.; Rempt, M.; Inzé, D.; Pohnert, G.; Vuylsteke, M.; Vyverman, W. Metabolomics Enables the Structure Elucidation of a Diatom Sex Pheromone. Angew. Chem. Int. Ed. Engl. 2013, 52, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Denslow, N.; Colbourne, J.K.; Dix, D.; Freedman, J.H.; Helbing, C.C.; Kennedy, S.; Williams, P.L. Selection of surrogate animal species for comparative toxicogenomics. In Genomic Approaches for Cross-Species Extrapolation in Toxicology; Di Giulio, R., Benson, W.H., Eds.; Taylor and Francis: Washington, DC, USA, 2007. [Google Scholar]
- Lampert, W. Daphnia: Development of a Model Organism in Ecology and Evolution; Excellence in Ecology; International Ecology Institute: Luhe, Germany, 2011; Volume 21. [Google Scholar]
- U.S. Department of Health and Human Services: National Institutes of Health. Available online: https://www.nigms.nih.gov/Research/models/Pages/default.aspx (accessed on 20 June 2018).
- Garreta-Laura, E.; Campos, B.; Barata, C.; Lacorte, S.; Tauler, R. Metabolic profiling of Daphnia magna exposed to environmental stressors by GC-MS and chemometric tools. Metabolomics 2016, 12, 86. [Google Scholar] [CrossRef]
- Southam, A.D.; Payne, T.G.; Cooper, H.J.; Arvanitis, T.N.; Viant, M.R. Dynamic Range and Mass Accuracy of Wide-Scan Direct Infusion Nanoelectrospray Fourier Transform Ion Cyclotron Resonance Mass Spectrometry-Based Metabolomics Increased by the Spectral Stitching Method. Anal. Chem. 2007, 79, 4595–4602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, J.; Liebeke, M.; Sommer, U.; Viant, M.R.; Bundy, J.G.; Ebbels, T.M.D. Statistical correlations between NMR spectroscopy and direct infusion FT-ICR mass spectrometry aid annotation of unknowns in metabolomics. Anal. Chem. 2016, 88, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Lawson, T.N.; Weber, R.J.M.; Dunn, W.B.; Viant, M.R. Deep Metabolome Annotation of the Model Organism Daphnia magna. Unpublished work. 2018. [Google Scholar]
- Edison, A.S.; Hall, R.D.; Junot, C.; Karp, P.D.; Kurland, I.J.; Mistrik, R.; Reed, L.K.; Saito, K.; Salek, R.M.; Steinbeck, C. The time is right to focus on model organism metabolomes. Metabolites 2016, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L. Promoting health and longevity through diet: From model organisms to humans. Cell 2015, 161, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Pallanca, J.E. Ascorbate metabolism in relation to oxidative stress. Biochem. Soc. Trans. 1996, 24, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frei, B.; England, L.; Ames, B.N. Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Natl. Acad. Sci. USA 1989, 86, 6377–6381. [Google Scholar] [CrossRef] [PubMed]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress part 1: Mechanisms involved in metal-induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Lester, R.; Carey, M.C.; Little, J.; Cooperstein, L.A.; Down, S.R. Crustacean intenstinal detergent promotes sterol solubilisation. Science 1975, 189, 1098–1100. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, S.; Miyachi, S. Sulfolipid metabolism in Chlorella. Plant Physiol. 1966, 41, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Van den Oord, A.; Danielsson, H.; Ryhage, R. On the structure of the emulsifiers in gastric juice from the crab, Cancer pagurus L. J. Biol. Chem. 1965, 240, 2242–2247. [Google Scholar] [PubMed]
- Collatz, K.-G.; Mommsen, G.; Mommsen, T. Alkylsulfuric acid and glyceride type emulsifiers in the intestinal juice of the snail Helix pomatia L. J. Comp. Physiol. 1975, 96, 123–129. [Google Scholar] [CrossRef]
- De Coen, W.; Janssen, C.R. The missing biomarker link: Relationships between effects on the cellular energy allocation biomarker of toxicant-stressed Daphnia magna and corresponding population characteristics. Environ. Toxicol. Chem. 2003, 22, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- MEtabolomics standaRds Initiative in Toxicology (MERIT) Project. Available online: http://www.ecetoc.org/topics/standardisation-metabolomics-assays-regulatory-toxicology/ (accessed on 6 June 2018).
- Terry, A.; Cambridge Environmental Assessments, Cambridge, UK. Personal communication, 2018.
- Weber, R.J.M.; Viant, M.R. MI-Pack: Increased confidence of metabolite identification in mass spectra by integrating accurate masses and metabolic pathways. Chemometr. Intell. Lab. 2010, 104, 75–82. [Google Scholar] [CrossRef]
- Wold, S.; Sjostrom, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemometr. Intell. Lab. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.J.; van Duijnhoven, J.P.M.; van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank Order of Peak Importance in PLS-R Model | Unique Empirical Formulae | Metabolite Name(s) | MSI Level | Measured m/z (Intensity Correlation between Peak-Pairs) | Ion Form(s) | m/z Error (ppm) | Supporting Peak Annotations (Other Adducts or Isotopes) |
|---|---|---|---|---|---|---|---|
| 5 | C6H8O6 | Ascorbate | 1 (compared to authentic chemical standard) | 175.02480 | [M − H]− | −0.08 | [M + Na − 2H]− m/z 197.00675 (−0.04 ppm error), [M + Cl]− m/z 211.00136 (−0.62 ppm) |
| 13, 6 | C12H18O6 | Unknown | 2 (accurate m/z, peak correlation, isotope pattern) | 257.10306, 258.10642 (r = 0.996) | [M − H]−, [M(13C) − H]− | −0.01, 0.01 | − |
| 9 | C12H24SO4 | Sulfonated lipid | 2 (accurate m/z, MS/MS; but no standard available) | 263.13227 | [M − H]− | 0.05 | − |
| 17 | C12H24SO6 | Sulfonated lipid | 2 (accurate m/z, MS/MS; but no standard available) | 295.12191 | [M − H]− | −0.60 | [M + Na − 2H]− m/z 317.10402 (−0.03 ppm), [M + K − 2H]− m/z 333.07797 (0.00 ppm) |
| 23, 31 | C7H10O7 | 3-Hydroxybutane-1,2,3-tricarboxylate, 2-hydroxybutane-1,2,4-tricarboxylate, 2-methylcitrate and/or homoisocitrate | 2 (accurate m/z, peak correlation, isotope pattern) | 242.01539, 243.00911 (r = 0.970) | [M(13C) + Cl]−, [M + (37Cl)]− | −0.09, 0.01 | − |
| 25 | C11H24SO4 | Sulfonated lipid | 2 (accurate m/z, MS/MS, isotope pattern; but no standard available) | 252.13562 | [M(13C) − H]− | 0.04 | [M − H]− m/z 251.13225 (−0.02 ppm) |
| 26 | C10H20SO5 | Sulfonated lipid | 2 (accurate m/z, MS/MS; but no standard available) | 251.09579 | [M − H]− | −0.32 | − |
| 43 | C8H15NO6 | N-Acetyl-D-galactosamine, N-acetyl-D-glucosamine and/or N-acetyl-D-mannosamine | 2 (accurate m/z, isotope pattern) | 258.05642 | [M + (37Cl)]− | 0.11 | [M − H]− m/z 220.08268 (0.08 ppm), [M + Cl]− m/z 256.05935 (0.04 ppm), [M + K − 2H]− m/z 258.03844 (−0.41 ppm) |
| 49 | C14H23N10OP | Unknown | 2 (accurate m/z, isotope pattern) | 415.14588 | [M + (37Cl)]− | 0.09 | [M − H]− m/z 377.17266 (1.44 ppm), [M + Cl]− m/z 413.14865 (−0.35 ppm), [M(13C)+ Cl]− m/z 414.15214 (−0.02 ppm), [M(13C)+(37Cl)]− m/z 416.14882 (−0.91 ppm) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, N.S.; Gavin, A.; Viant, M.R. Metabolomics Discovers Early-Response Metabolic Biomarkers that Can Predict Chronic Reproductive Fitness in Individual Daphnia magna. Metabolites 2018, 8, 42. https://doi.org/10.3390/metabo8030042
Taylor NS, Gavin A, Viant MR. Metabolomics Discovers Early-Response Metabolic Biomarkers that Can Predict Chronic Reproductive Fitness in Individual Daphnia magna. Metabolites. 2018; 8(3):42. https://doi.org/10.3390/metabo8030042
Chicago/Turabian StyleTaylor, Nadine S., Alex Gavin, and Mark R. Viant. 2018. "Metabolomics Discovers Early-Response Metabolic Biomarkers that Can Predict Chronic Reproductive Fitness in Individual Daphnia magna" Metabolites 8, no. 3: 42. https://doi.org/10.3390/metabo8030042
APA StyleTaylor, N. S., Gavin, A., & Viant, M. R. (2018). Metabolomics Discovers Early-Response Metabolic Biomarkers that Can Predict Chronic Reproductive Fitness in Individual Daphnia magna. Metabolites, 8(3), 42. https://doi.org/10.3390/metabo8030042