NMR Metabolomic Analysis of Skeletal Muscle, Heart, and Liver of Hatchling Loggerhead Sea Turtles (Caretta caretta) Experimentally Exposed to Crude Oil and/or Corexit

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Metabolic Profiling
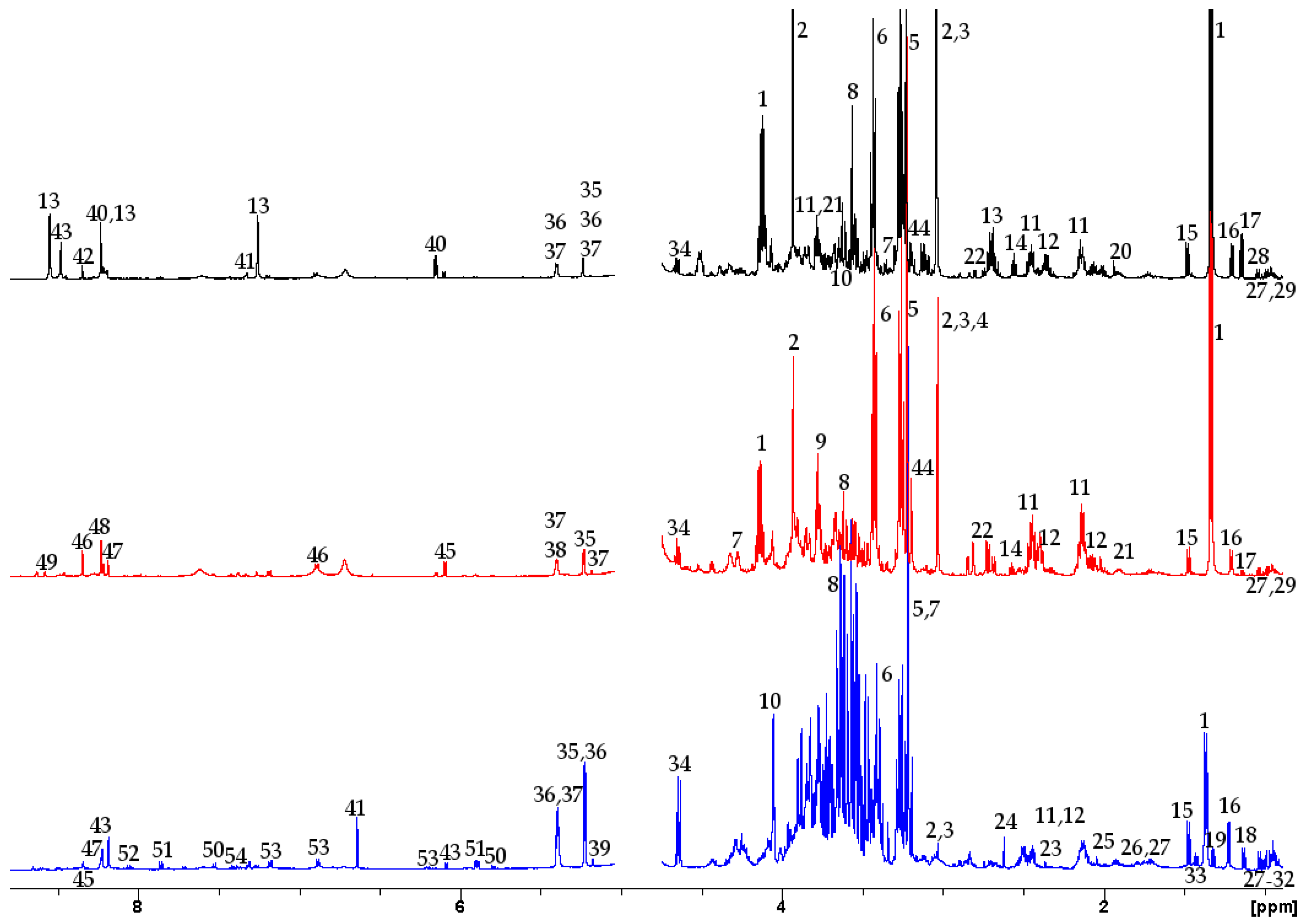
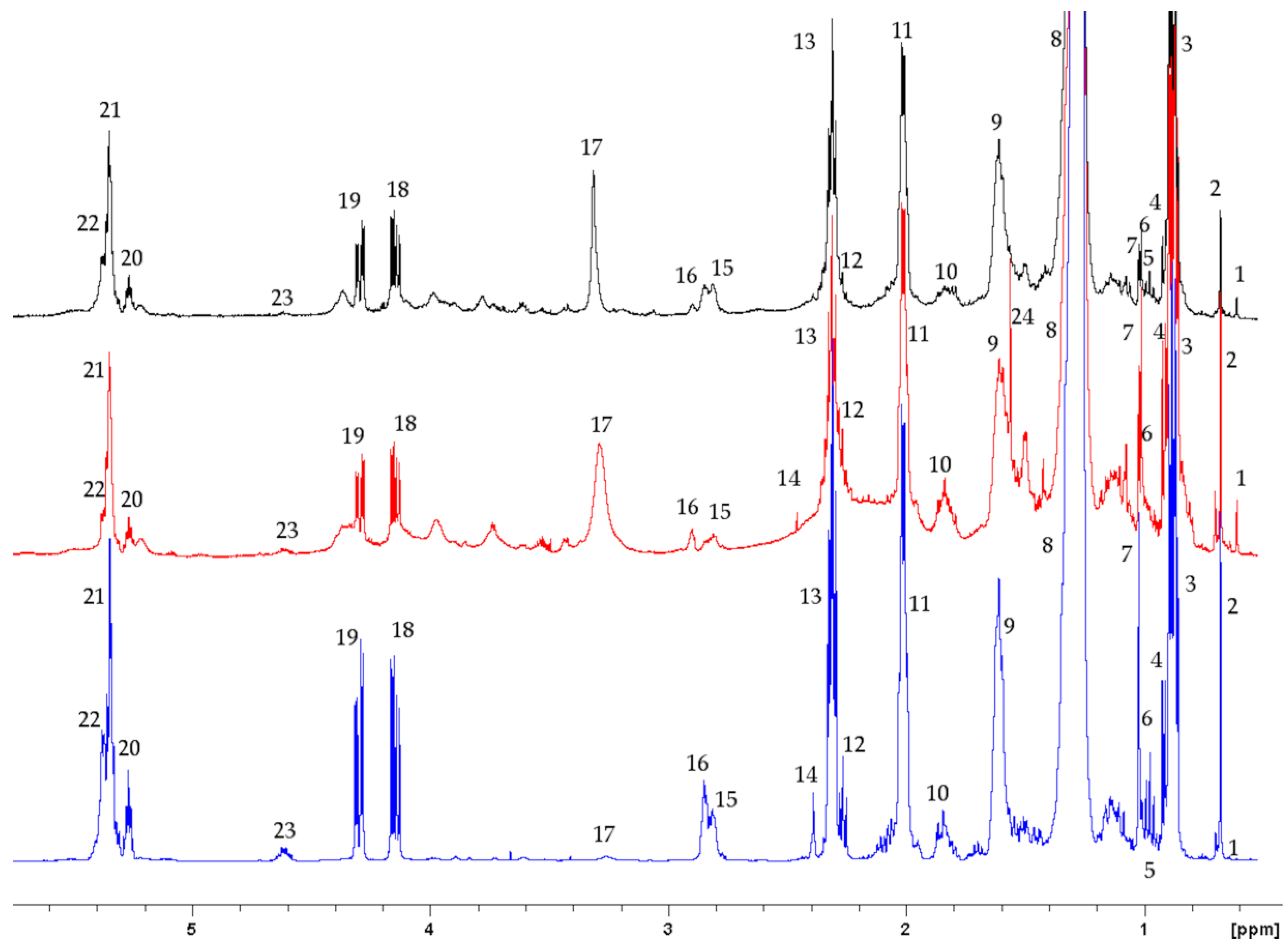
2.1.1. Aqueous Data
2.1.2. Lipophilic Data
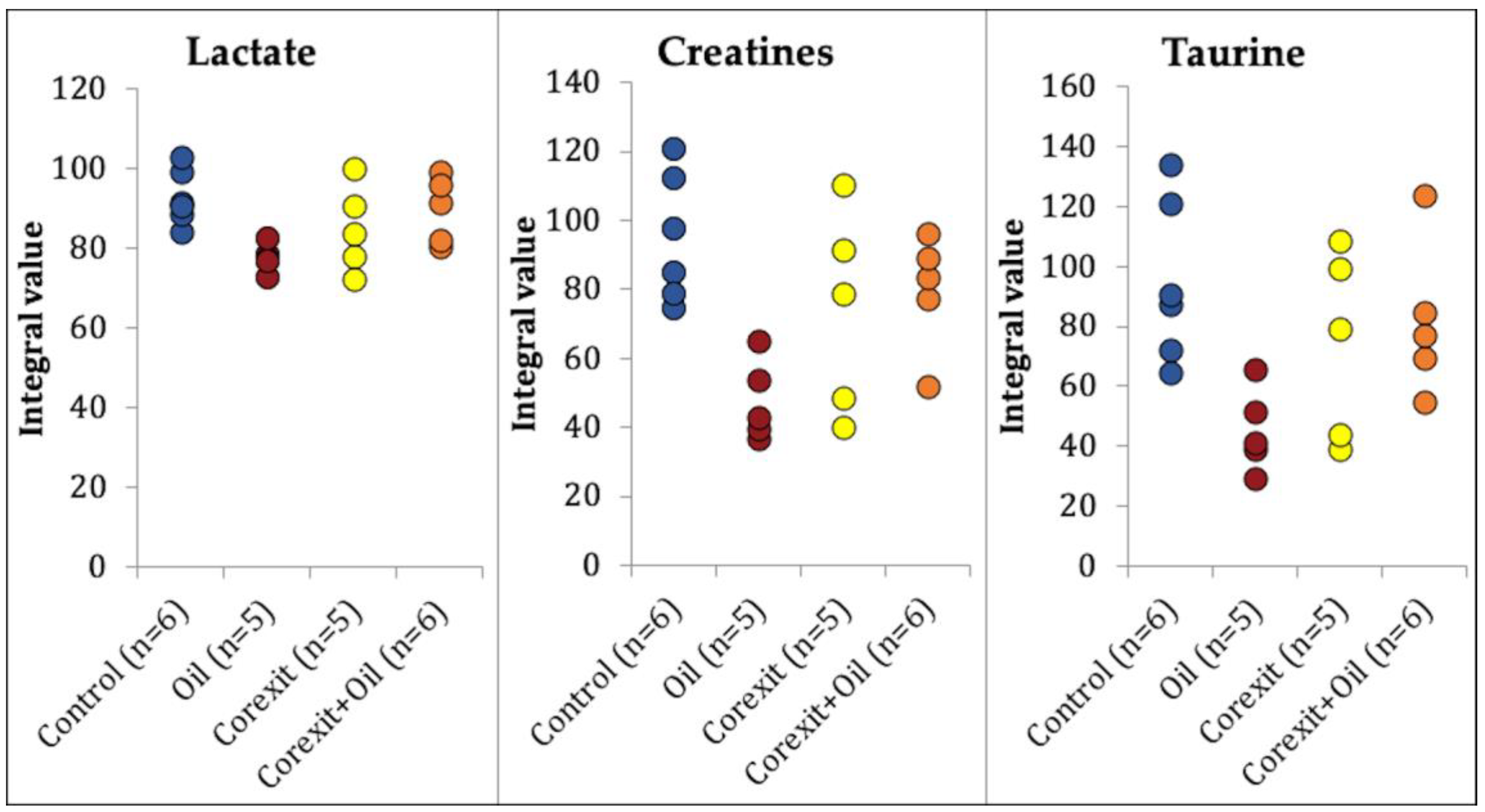
2.2. Statistical Analysis
2.2.1. Aqueous Data
2.2.2. Lipophilic Data
3. Discussion
4. Materials and Methods
4.1. Organisms, Oil/Dispersant Exposure, and Tissue Collection
4.2. Metabolite Extraction
4.3. 1H-NMR Metabolomics and Spectral Pre-Processing
4.4. 2D NMR and 1D 31P NMR Methods
4.5. NMR Data Processing and Analysis
4.6. Statistical Analysis of NMR Data
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
NMR Data Acquisition Methods
Skeletal Muscle
Heart
Liver
References
- Wallace, B.P.; Stacy, B.A.; Rissing, M.; Cacela, D.; Garrison, L.P.; Graettinger, G.D.; Holmes, J.V.; McDonald, T.; McLamb, D.; Schroeder, B. Estimating sea turtle exposures to Deepwater Horizon oil. Endang. Species Res. 2017, 33, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Sydeman, W.J.; Poloczanska, E.; Reed, T.E.; Thompson, S.A. Climate change and marine vertebrates. Science 2015, 350, 772–777. [Google Scholar] [CrossRef] [PubMed]
- The IUCN Red List of Threatened Species, Version 2018-1. Available online: www.iucnredlist.org (accessed on 28 August 2018).
- Campagna, C.; Short, F.T.; Polidoro, B.A.; McManus, R.; Collette, B.B.; Pilcher, N.J.; Sadovy de Mitcheson, Y.; Stuart, S.N.; Carpenter, K.E. Gulf of Mexico oil blowout increases risks to globally threatened species. Bioscience 2011, 61, 393–397. [Google Scholar] [CrossRef]
- Ylitalo, G.M.; Collier, T.K.; Anulacion, B.F.; Juaire, K.; Boyer, R.H.; da Silva, D.A.; Keene, J.L.; Stacy, B.A. Determining oil and dispersant exposure in sea turtles from the northern Gulf of Mexico resulting from the Deepwater Horizon oil spill. Endang. Species Res. 2017, 33, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3. [Google Scholar] [CrossRef]
- Ryan, D.; Robards, K. Metabolomics: The greatest omics of them all? Anal. Chem. 2006, 78, 7954–7958. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Doskocz, M.; Jodłowska, E.; Zacharzewska, A.; Łakomiec, J.; Czaja, K.; Kujawski, J. NMR techniques in metabolomic studies: A quick overview on examples of utilization. Appl. Magn. Reson. 2017, 48, 1–21. [Google Scholar] [CrossRef]
- Lin, C.Y.; Anderson, B.S.; Phillips, B.M.; Peng, A.C.; Clark, S.; Voorhees, J.; Wu, H.I.; Martin, M.J.; McCall, J.; Todd, C.R.; et al. Characterization of the metabolic actions of crude versus dispersed oil in salmon smolts via NMR-based metabolomics. Aquat. Toxicol. 2009, 95, 230–238. [Google Scholar] [CrossRef]
- Van Scoy, A.R.; Lin, C.Y.; Anderson, B.S.; Philips, B.M.; Martin, M.J.; McCall, J.; Todd, C.R.; Crane, D.; Sowby, M.L.; Viant, M.R.; et al. Metabolic responses produced by crude versus dispersed oil in Chinook salmon pre-smolts via NMR-based metabolomics. Ecotoxicol. Environ. Saf. 2010, 73, 710–717. [Google Scholar] [CrossRef]
- Stieglitz, J.D.; Mager, E.M.; Hoenig, R.H.; Benetti, D.D.; Grosell, M. Impacts of Deepwater Horizon crude oil exposure on adult mahi-mahi (Coryphaena hippurus) swim performance. Environ. Toxicol. Chem. 2016, 35, 2613–2622. [Google Scholar] [CrossRef]
- Brette, F.; Shiels, H.A.; Galli, G.L.J.; Cros, C.; Incardona, J.P.; Scholz, N.L.; Block, B.A. A novel cardiotoxic mechanism for a pervasive global pollutant. Sci. Rep. 2017, 7, 41476. [Google Scholar] [CrossRef] [PubMed]
- Lutcavage, M.E.; Lutz, P.L.; Bossart, G.D.; Hudson, D.M. Physiologic and clinicopathologic effects of crude oil on loggerhead sea turtles. Arch. Environ. Contam. Toxicol. 1995, 28, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Harms, C.A.; McClellan-Green, P.; Godfrey, M.H.; Christiansen, E.F.; Broadhurst, H.J.; Godard-Codding, C. Clinical pathology effects of crude oil and dispersant on hatchling loggerhead sea turtles (Caretta caretta). In Proceedings of the 45th Annual International Association for Aquatic Animal Medicine, Gold Coast, Australia, 17–22 May 2014; pp. 17–22. [Google Scholar]
- Hill, J.; Olson, E. An Introduction to Muscle. In Muscle: Fundamental Biology and Mechanisms of Disease; Hill, J., Olson, E., Eds.; Elsevier Science & Technology: San Diego, CA, USA, 2012; Chapter 1; pp. 3–9. [Google Scholar]
- Rubio, R.; Berne, R.M.; Dobson, J.G., Jr. Sites of adenosine production in cardiac and skeletal muscle. Am. J. Physiol. 1973, 225, 938–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleeson, T.T.; Dalessio, P.M. Lactate and glycogen metabolism in the lizard Dipsosaurus dorsalis following exhaustive exercise. J. Exp. Biol. 1989, 144, 377–393. [Google Scholar]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.; Blanco, G. Metabolism in Some Tissues. In Medical Biochemistry; Blanco, A., Blanco, G., Eds.; Academic Press: London, UK, 2017; Chapter 20; pp. 447–463. [Google Scholar]
- Gleeson, T.T.; Dalessio, P.M. Lactate: A substrate for reptilian muscle gluconeogenesis following exhaustive exercise. J. Comp. Physiol. B 1990, 160, 331–338. [Google Scholar] [CrossRef]
- Gagnon, M.M.; Holdway, D.A. Metabolic enzyme activities in fish gills as biomarkers of exposure to petroleum hydrocarbons. Ecotoxicol. Environ. Saf. 1999, 44, 92–99. [Google Scholar] [CrossRef]
- Van Pilsum, J.F.; Stephens, G.C.; Taylor, D. Distribution of creatine, guanidinoacetate and the enzymes for their biosynthesis in the animal kingdom. Implications for phylogeny. Biochem. J. 1972, 126, 325–345. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, S.W.; Ju Jong, C.; Ramila, K.C.; Azuma, J. Physiological roles of taurine in heart and muscle. J. Biomed. Sci. 2010, 17, S2. [Google Scholar] [CrossRef]
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Purine and pyrimidine metabolism. In Medical Biochemistry; Blanco, A., Blanco, G., Eds.; Academic Press: London, UK, 2017; Chapter 18; pp. 413–423. [Google Scholar]
- Incardona, J.P.; Gardner, L.D.; Linbo, T.L.; Brown, T.L.; Esbaugh, A.J.; Mager, E.M.; Stieglitz, J.D.; French, B.L.; Labenia, J.S.; Laetz, C.A.; et al. Deepwater Horizon crude oil impacts the developing hearts of large predatory pelagic fish. Proc. Natl. Acad. Sci. USA 2014, 201320950. [Google Scholar] [CrossRef]
- Esbaugh, A.J.; Mager, E.M.; Stieglitz, J.D.; Hoenig, R.; Brown, T.L.; French, B.L.; Linbo, T.L.; Lay, C.; Forth, H.; Scholz, N.L.; et al. The effects of weathering and chemical dispersion on Deepwater Horizon crude oil toxicity to mahi-mahi (Coryphaena hippurus) early life stages. Sci. Total Environ. 2016, 543, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Derickson, W.K. Lipid storage and utilization in reptiles. Am. Zool. 1976, 16, 711–723. [Google Scholar] [CrossRef]
- Price, E.R. The physiology of lipid storage and use in reptiles. Biol. Rev. 2017, 92, 1406–1426. [Google Scholar] [CrossRef] [PubMed]
- Committee on Oil in the Sea: Inputs, Fates, and Effects, Staff National Research Council, National Research Council, and National Academy of Sciences. Oil in the Sea III: Inputs, Fates, and Effects; National Academies Press: Washington, DC, USA, 2003; 277p. [Google Scholar]
- Korn, S.; Moles, D.A.; Rice, S.D. Effects of temperature on the median tolerance limit of pink salmon and shrimp exposed to toluene, naphthalene, and Cook Inlet crude oil. Bull. Environ. Contam. Toxicol. 1979, 21, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Bembenek Bailey, S.A.; Niemuth, J.N.; McClellan-Green, P.D.; Godfrey, M.H.; Harms, C.A.; Stoskopf, M.K. 1H-NMR metabolomic study of whole blood from hatchling loggerhead sea turtles (Caretta caretta) exposed to crude oil and/or Corexit. R. Soc. Open Sci. 2017, 4, 171433. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E. HMDB 3.0—The human metabolome database in 2013. Nucleic Acids Res. 2012, 41, D801–D807. [Google Scholar] [CrossRef]
- AOCS Lipid Library: NMR Spectroscopy of Fatty Acids and Their Derivatives. Available online: http://lipidlibrary.aocs.org/Analysis/content.cfm?ItemNumber=40808 (accessed on 28 August 2018).
- Oostendorp, M.; Engelke, U.; Willemsen, M.; Wevers, R. Diagnosing inborn errors of lipid metabolism with proton nuclear magnetic resonance spectroscopy. Clin. Chem. 2006, 52, 1395–1405. [Google Scholar] [CrossRef]
- Subramanian, A.; Shankar Joshi, B.; Roy, A.D.; Roy, R.; Gupta, V.; Dang, R.S. NMR spectroscopic identification of cholesterol esters, plasmalogen and phenolic glycolipids as fingerprint markers of human intracranial tuberculomas. NMR Biomed. 2008, 21, 272–288. [Google Scholar] [CrossRef]
- Ekman, D.R.; Teng, Q.; Villeneuve, D.L.; Kahl, M.D.; Jensen, K.M.; Durhan, E.J.; Ankley, G.T.; Collette, T.W. Profiling lipid metabolites yields unique information on sex-and time-dependent responses of fathead minnows (Pimephales promelas) exposed to 17α-ethynylestradiol. Metabolomics 2009, 5, 22. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillén, M.D. A method based on 1H NMR spectral data useful to evaluate the hydrolysis level in complex lipid mixtures. Food Res. Int. 2014, 66, 379–387. [Google Scholar] [CrossRef]
- Nakagawa, S.; Cuthill, I.C. Effect size, confidence interval and statistical significance: A practical guide for biologists. Biol. Rev. 2007, 82, 591–605. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Metabolite | Tissue | Metabolite | Tissue |
|---|---|---|---|
| Organic acids/Osmolytes | |||
| Acetate | S* | Glutathione | H |
| Carnosine | S | Glycerophosphocholine | S, H, L* |
| Choline | S, H, L* | Lactate | S#, H, L* |
| Creatinine | H* | Myo-inositol | S, L* |
| Formate | S | Phosphocholine | S, H, L* |
| Energy compounds | |||
| Creatine | S#, H, L* | Mannose | L |
| Glucose (α & β) | S, H, L* | Ribose | S, H, L* |
| Glycogen | H* | Phosphocreatine | S#, H, L* |
| Maltose | S, L* | ||
| Amino acids | |||
| Alanine | S, H, L* | Histidine | S, L * |
| Alanine dimer | L* | Homoserine | S, H |
| Alloisoleucine | L* | Isoleucine | S, H, L* |
| Arginine | L* | Leucine | S, H, L* |
| Aspartate | S, H* | Methylamine | L* |
| Beta-alanine | S, H* | Phenylalanine | L* |
| Glutamate | S, H, L* | Taurine | S#, H, L* |
| Glutamine | S, H, L* | Threonine | L* |
| Glycine | S, H, L* | Tyrosine | L* |
| Glycylproline | L* | Valine | S, H, L* |
| Ketone bodies | |||
| 3-hydroxybutyrate | S, H, L* | 2-hydroxyvalerate | L |
| 2-hydroxybutyrate | L* | Pyruvate | L* |
| Nucleosides & Nucleotides | |||
| Adenine | H, (L*) | Inosine | S, L* |
| Adenosine | H | Inosine monophosphate | S |
| Adenosine diphosphate | H | Nicotinamide adenine dinucleotide | H |
| Adenosine triphosphate | H, L* | Uracil | L* |
| Cytidine | L* | Uridine | L* |
| Other | |||
| Propylene glycol | S, H* | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bembenek-Bailey, S.A.; Niemuth, J.N.; McClellan-Green, P.D.; Godfrey, M.H.; Harms, C.A.; Gracz, H.; Stoskopf, M.K. NMR Metabolomic Analysis of Skeletal Muscle, Heart, and Liver of Hatchling Loggerhead Sea Turtles (Caretta caretta) Experimentally Exposed to Crude Oil and/or Corexit. Metabolites 2019, 9, 21. https://doi.org/10.3390/metabo9020021
Bembenek-Bailey SA, Niemuth JN, McClellan-Green PD, Godfrey MH, Harms CA, Gracz H, Stoskopf MK. NMR Metabolomic Analysis of Skeletal Muscle, Heart, and Liver of Hatchling Loggerhead Sea Turtles (Caretta caretta) Experimentally Exposed to Crude Oil and/or Corexit. Metabolites. 2019; 9(2):21. https://doi.org/10.3390/metabo9020021
Chicago/Turabian StyleBembenek-Bailey, Stasia A., Jennifer N. Niemuth, Patricia D. McClellan-Green, Matthew H. Godfrey, Craig A. Harms, Hanna Gracz, and Michael K. Stoskopf. 2019. "NMR Metabolomic Analysis of Skeletal Muscle, Heart, and Liver of Hatchling Loggerhead Sea Turtles (Caretta caretta) Experimentally Exposed to Crude Oil and/or Corexit" Metabolites 9, no. 2: 21. https://doi.org/10.3390/metabo9020021
APA StyleBembenek-Bailey, S. A., Niemuth, J. N., McClellan-Green, P. D., Godfrey, M. H., Harms, C. A., Gracz, H., & Stoskopf, M. K. (2019). NMR Metabolomic Analysis of Skeletal Muscle, Heart, and Liver of Hatchling Loggerhead Sea Turtles (Caretta caretta) Experimentally Exposed to Crude Oil and/or Corexit. Metabolites, 9(2), 21. https://doi.org/10.3390/metabo9020021