The Astrobiology of Alien Worlds: Known and Unknown Forms of Life



Abstract
:1. Introduction
2. The Definition Issue
2.1. Common Components of Modern Definitions
2.2. Argument against a Definition
2.3. Necessity for a Definition
3. Life as We Know It
3.1. Biological Form and Function as Commonly Observed on Earth
3.2. Amorphous Organic Forms
3.3. Amorphous Conglomerate Forms
4. Known Trajectory of Life on Earth
5. Alternative Forms of Life
5.1. Alternative Biochemistries
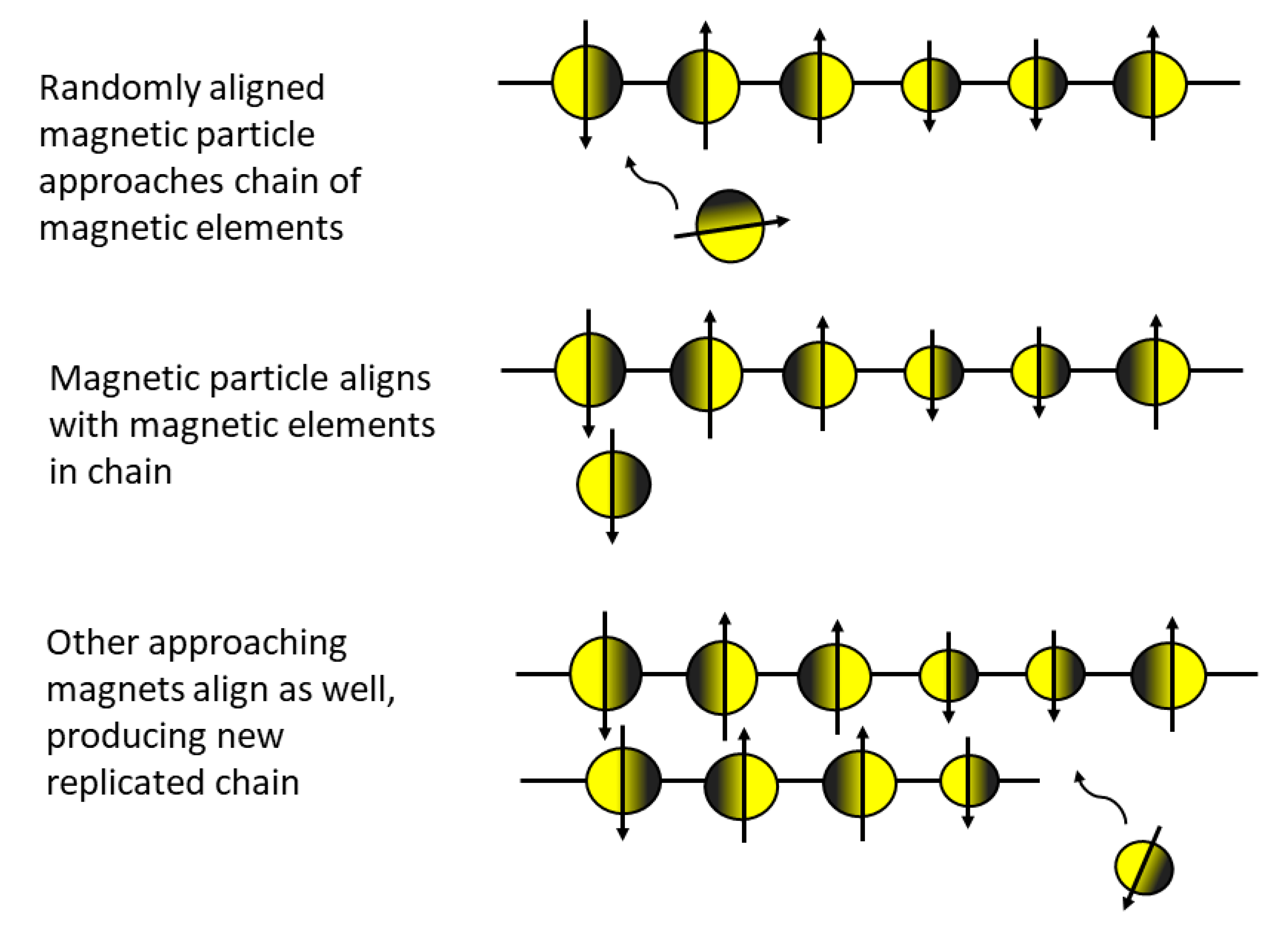
5.2. Unbounded Inorganic Forms
5.3. Amorphous Organic Forms
5.4. Mechanical Forms
6. Plausible Evolutionary Trajectories on Other Worlds
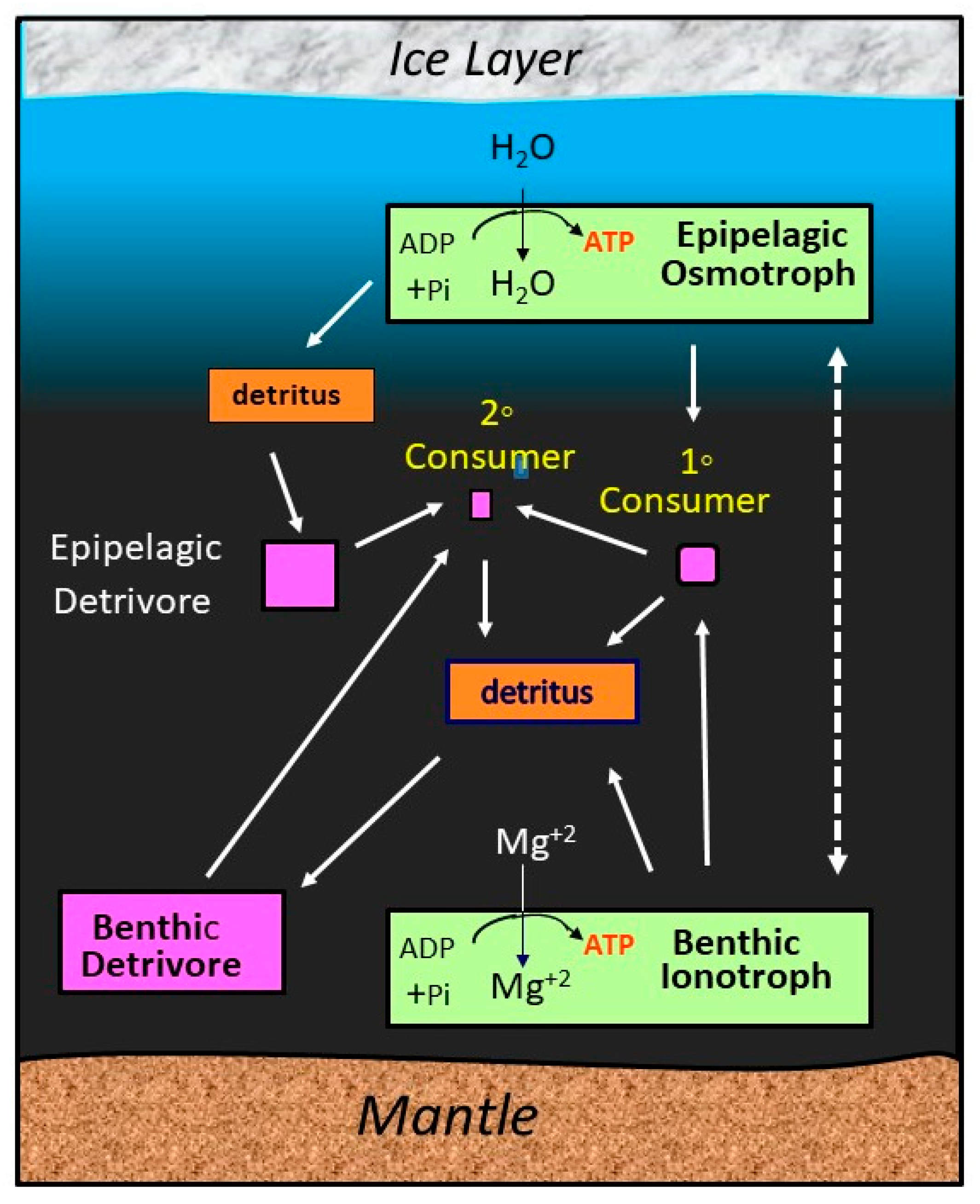
6.1. Life in a Subsurface Ocean of an Icy Planet or Moon
6.2. Life on a Barren Planet
6.3. Life on a Hydrocarbon World
6.4. Life on a Rogue Planet
6.5. Life on a Super-Earth
6.6. Life on a Tidally Locked Red Dwarf Planet
6.7. Life in a Planetary Atmosphere
7. Implications for Fermi Paradox
8. Ongoing Studies and Prospects for Further Investigation
9. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Domagal-Goldman, S.D.; Wright, K.E.; Adamala, K.; de la Rubia, L.A.; Bond, J.; Dartnell, L.R.; Goldman, A.D.; Lynch, K.; Naud, M.E.; Paulino-Lima, I.G.; et al. The Astrobiology Primer v2.0. Astrobiology 2016, 16, 561–653. [Google Scholar] [PubMed] [Green Version]
- Schroedinger, E. What Is Life? The Physical Aspect of the Living Cell, 1st ed.; Cambridge University Press: Cambridge, UK, 1944. [Google Scholar]
- Margulis, L.; Sagan, D. What Is Life; Simon & Schuster: New York, NY, USA, 1995. [Google Scholar]
- Luisi, P.L. About various definitions of life. Orig. Life Evol. Biosph. 1998, 28, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Gánti, T. The Principles of Life; Oxford University Press: New York, NY, USA, 2003. [Google Scholar]
- Irwin, L.N.; Schulze-Makuch, D. Cosmic Biology: How Life Could Evolve on Other Worlds, 1st ed.; Praxis: New York, NY, USA, 2011. [Google Scholar]
- Schulze-Makuch, D.; Irwin, L.N. Life in the Universe: Expectations and Constraints, 3rd ed.; Springer Praxis: Cham, Switzerland, 2018. [Google Scholar]
- Cleland, C.E. The Quest for a Universal Theory of Life: Searching for Life as We don’t Know it; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Pályi, G.; Zucchi, C.; Caglioti, L. Fundamentals of Life; Elsevier SAS: Amsterdam, The Netherlands, 2002; pp. 169–179. [Google Scholar]
- Cleland, C.E.; Chyba, C.F. Defining ‘life’. Orig. Life Evol. Biosph. 2002, 32, 387–393. [Google Scholar] [CrossRef]
- Cleland, C.E. Life without definitions. Synthese 2012, 185, 125–144. [Google Scholar] [CrossRef]
- Cleland, C.E. Moving Beyond Definitions in the Search for Extraterrestrial Life. Astrobiology 2019, 19, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NASA. About Life Detection. 2020. Available online: https://astrobiology.nasa.gov/research/life-detection/about/ (accessed on 11 June 2020).
- Schulze-Makuch, D.; Guan, H.; Irwin, L.N.; Vega, E. Redefining life: An ecological, thermodynamic, and bioinformatic approach. In Fundamentals of Life; Pályi, G., Zucchi, C., Caglioti, L., Eds.; Elsevier SAS: Amsterdam, The Netherlands, 2002; pp. 169–179. [Google Scholar]
- Claeys, P.; Morbidelli, A. Late heavy bombardment. In Encyclopedia of Astrobiology; Gargaud, M., Amils, R., Quintanilla, J.C., Cleaves, H.J., Irvine, W.M., Pinti, D.L., Viso, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 909–912. [Google Scholar]
- Schopf, J.W. Microfossils of the early Archaen apex chert: New evidence of the antiquity of life. Science 1983, 280, 640–646. [Google Scholar]
- Schopf, J.W. Cradle of Life: The Discovery of Earth’s Earliest Fossils; Princeton University Press: Princeton, NJ, USA, 1999; p. 355. [Google Scholar]
- Bell, E.A.; Boehnke, P.; Harrison, T.M.; Mao, W.L. Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. Proc. Natl. Acad. Sci. USA 2015, 112, 14518–14521. [Google Scholar] [CrossRef] [Green Version]
- Dodd, M.S.; Papineau, D.; Grenne, T.; Slack, J.F.; Rittner, M.; Pirajno, F.; O’Neil, J.; Little, C.T. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 2017, 543, 60–64. [Google Scholar] [CrossRef]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubi, E.; Lane, N. The Origin of Life in Alkaline Hydrothermal Vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, K.; Swenson, R. Thermodynamics in the present progressive mode and its role in the context of the origin of life. Biosystems 1999, 51, 53–61. [Google Scholar] [CrossRef]
- Russell, M. First life. Am. Sci. 2006, 94, 32–39. [Google Scholar] [CrossRef]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal Chemistry and the Origin of Cellular Life. Astrobiology 2019, 19, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Locker-Grutjen, O.; Mayer, C. Hypothesis: Origin of life in deep-reaching tectonic faults. Orig. Life Evol. Biosph. 2012, 42, 47–54. [Google Scholar] [CrossRef]
- Mayer, C.; Schreiber, U.; Davila, M.J.; Schmitz, O.J.; Bronja, A.; Meyer, M.; Klein, J.; Meckelmann, S.W. Molecular Evolution in a Peptide-Vesicle System. Life 2018, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Mayer, C.; Schreiber, U.; Davila, M.J. Selection of Prebiotic Molecules in Amphiphilic Environments. Life 2017, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Westall, F.; Hickman-Lewis, K.; Hinman, N.; Gautret, P.; Campbell, K.A.; Breheret, J.G.; Foucher, F.; Hubert, A.; Sorieul, S.; Dass, A.V.; et al. A Hydrothermal-Sedimentary Context for the Origin of Life. Astrobiology 2018, 18, 259–293. [Google Scholar] [CrossRef]
- Ardell, D.H.; Sella, G. On the evolution of redundancy in genetic codes. J. Molec. Evol. 2001, 53, 269–281. [Google Scholar] [CrossRef]
- Zhang, Y.; Baranov, P.V.; Atkins, J.F.; Gladyshev, V.N. Pyrrolysine and selenocysteine use dissimilar coding strategies. J. Biol. Chem. 2005, 280, 20740–20751. [Google Scholar] [CrossRef] [Green Version]
- Beck, C.H.M.; Irwin, L.N. The Evolutionary Imperative: Why Change Happens, Where It Leads, and How We Might Survive; CCB Publishing: Vancouver, BC, Canada, 2016; p. 341. [Google Scholar]
- Schulze-Makuch, D.; Airo, A.; Schirmack, J. The adaptability of life on earth and the diversity of planetary habitats. Front. Microbiol. 2017, 8, 2011. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner, J.T. The Social Amoebae: The Biology of Cellular Slime Molds; Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Erdos, G.W.; Raper, K.B.; Vogen, L.K. Mating types and macrocyst formation in Dictyostelium Discoideum. Proc. Natl. Acad. Sci. USA 1973, 70, 1828–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.A.; Dreisbach, T.A.; Parks, C.G.; Flip, G.M.; Schmitt, C.L. Coarse-scale population structure of pathogenic Armillaria species in a mixed-conifer forest in the Blue Mountains of northeast Oregon. Can. J. For. Res. 2003, 33, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Sipos, G.; Anderson, J.B.; Nagy, L.G. Armillaria. Curr. Biol. 2018, 28, R297–R298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingraham, J.L. March of the Microbes: Sighting the Unseen; Harvard University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Anderson, J.B.; Bruhn, J.N.; Kasimer, D.; Wang, H.; Rodrigue, N.; Smith, M.L. Clonal evolution and genome stability in a 2500-year-old fungal individual. Proc. Biol. Sci. 2018, 285, 20182233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mock, K.E.; Rowe, C.A.; Hooten, M.B.; Dewoody, J.; Hipkins, V.D. Clonal dynamics in western North American aspen (Populus tremuloides). Mol. Ecol. 2008, 17, 4827–4844. [Google Scholar] [CrossRef]
- Honegger, R. Fungal evolution: Symbiosis and morphogenesis. In Symbiosis as a Source of Evolutionary Innovation; Margulis, L., Fester, R., Eds.; MIT Press: Cambridge, MA, USA, 1991; pp. 319–340. [Google Scholar]
- Noffke, N. Ancient sedimentary structures in the <3.7 Ga Gillespie Lake Member, Mars, that resemble macroscopic morphology, spatial associations, and temporal succession in terrestrial microbialites. Astrobiology 2015, 15, 169–192. [Google Scholar]
- Vissche, P.T.; Stolz, J.F. Microbial mats as bioreactors: Populations, processes, and products. In Geobiology: Objectives, Concepts, Perspectives; Noffke, N., Ed.; Elsevier: New York, NY, USA, 2005; pp. 87–100. [Google Scholar]
- Riding, R. Microbial carbonates: The geological record of calcified bacterial–algal mats and biofilms. Sedimentology 2000, 47, 179–214. [Google Scholar] [CrossRef]
- Noffke, N.; Christian, D.; Wacey, D.; Hazen, R.M. Microbially induced sedimentary structures recording an ancient ecosystem in the ca. 3.48 billion-year-old Dresser Formation, Pilbara, Western Australia. Astrobiology 2013, 13, 1103–1124. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.; Ingham, J.; Zedef, V.; Maktav, D.; Sunar, F.; Hall, A.; Fallick, A. Search for signs of ancient life on Mars: Expectations from hydromagnesite microbialites, Salda Lake, Turkey. J. Geol. Soc. 1999, 156, 869–888. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Makuch, D.; Laval, B.; Irwin, L.N. The rise of complexity: Pavilion Lake microbialites suggest a pathway toward macroorganismic communities. Hypoth. Life Sci. 2012, 2, 55–59. [Google Scholar]
- Irwin, L.N.; Méndez, A.; Fairén, A.G.; Schulze-Makuch, D. Assessing the possibility of biological complexity on other worlds, with an estimate of the occurrence of complex life in the Milky Way galaxy. Challenges 2014, 214, 159–174. [Google Scholar] [CrossRef]
- Sheppard, C.R.C.; Davy, S.K.; Pilling, G.M.; Graham, N.A.J. The Biology of Coral Reefs, 2nd ed.; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Murphy, R.C. Coral Reefs: Cities Under the Seas; Darwin Press: Kent, UK, 2002. [Google Scholar]
- Gulko, D. Hawaiian Coral Reef Ecology; Mutual Publishing: Honolulu, HA, USA, 1998. [Google Scholar]
- Szathmáry, E.; Maynard-Smith, J. From replicators to reproducers: The first major transitions leading to life. J. Theor. Biol. 1997, 187, 555–571. [Google Scholar]
- Branciamore, S.; Gallori, E.; Szathmary, E.; Czaran, T. The origin of life: Chemical evolution of a metabolic system in a mineral honeycomb? J. Mol. Evol. 2009, 69, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Bains, W.; Schulze-Makuch, D. The cosmic zoo: The (near) inevitability of the evolution of complex, macroscopic life. Life 2016, 6, 25. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Bains, W. The Cosmic Zoo: Complex. Life on Many Worlds; Springer International: Chichester, UK, 2017. [Google Scholar]
- Wächtershäuser, G. The case for the chemoautotrophic origin of life in an iron-sulfur world. Orig. Life Evol. Biosph. 1990, 20, 173–176. [Google Scholar] [CrossRef]
- Miller, S.L.; Schopf, J.W.; Lazcano, A. Oparin’s “Origin of Life”: Sixty years later. J. Molec. Evol. 1997, 44, 351–353. [Google Scholar] [CrossRef]
- Blankenship, R.E. Origin and early evolution of photosynthesis. Photosynth Res. 1992, 33, 91–111. [Google Scholar] [CrossRef]
- Kondratieva, E.N.; Zhukov, V.G.; Ivanovsky, R.N.; Petushkova, U.P.; Monosov, E.Z. The capacity of phototrophic sulfur bacterium Thiocapsa roseopersicina for chemosynthesis. Arch. Microbiol. 1976, 108, 287–292. [Google Scholar] [CrossRef]
- Dillon, J.G.; Castenholz, R.W. Scytonemin, a cyanobacterial sheath pigment, protects against uvc radiation: Implications for early photosynthetic life. J. Phycol. 1999, 35, 673–681. [Google Scholar] [CrossRef]
- Björn, L.O.; Widell, S.; Wang, T. Evolution of UV-B regulation and protection in plants. Adv. Space Res. 2002, 30, 1557–1562. [Google Scholar] [CrossRef]
- Martin, W. Archaebacteria (Archaea) and the origin of the eukaryotic nucleus. Curr. Opin. Microbiol. 2005, 8, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Bains, W.; Schulze-Makuch, D. Mechanisms of evolutionary innovation point to genetic control logic as the key difference between prokaryotes and eukaryotes. J. Mol. Evol. 2015, 81, 34–53. [Google Scholar] [CrossRef] [Green Version]
- António, M.R.S.; Schulze-Makuch, D. Toward a new understanding of multicellularity. Hypoth. Life Sci. 2012, 2, 4–14. [Google Scholar]
- Ratcliff, W.C.; Herron, M.D.; Howell, K.; Pentz, J.T.; Rosenzweig, F.; Travisano, M. Experimental evolution of an alternating uni- and multicellular life cycle in Chlamydomonas reinhardtii. Nat. Commun. 2013, 4, 2742. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Makuch, D.; Irwin, L.N. Energy cycling and hypothetical organisms in Europa’s ocean. Astrobiology 2002, 2, 105–121. [Google Scholar] [CrossRef]
- Peng, Q.; Tong, H. The physics of strong magnetic fields in neutron stars. Mon. Not. R. Astron. Soc. 2007, 378, 159–162. [Google Scholar] [CrossRef]
- Muller, A.W.J. Were the First Organisms Heat Engines-A New Model for Biogenesis and the Early Evolution of Biological Energy-Conversion. Progr. Biophys. Molec. Biol. 1995, 63, 193–231. [Google Scholar] [CrossRef]
- Muller, A.W. Finding extraterrestrial organisms living on thermosynthesis. Astrobiology 2003, 3, 555–564. [Google Scholar] [CrossRef]
- Gusev, V.A.; Schulze-Makuch, D. Low frequency electromagnetic waves as a supplemental energy source to sustain microbial growth? Naturwissenschaften 2005, 92, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Dadachova, E.; Casadevall, A. Ionizing radiation: How fungi cope, adapt, and exploit with the help of melanin. Curr. Opin. Microbiol. 2008, 11, 525–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Makuch, D.; Grinspoon, D.H. Biologically Enhanced Energy and Carbon Cycling on Titan? Astrobiology 2005, 5, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Eisch, J.J.; Munson, P.R.; Gitua, J.N. The Potential of photochemical transition metal reactions in prebiotic organic synthesis. I. Observed conversion of methanol into ethylene glycol as possible prototype for sugar alcohol formation. Orig. Life Evol. Biosph. 2004, 34, 441–454. [Google Scholar] [CrossRef]
- Petkowski, J.J.; Bains, W.; Seager, S. On the Potential of Silicon as a Building Block for Life. Life 2020, 10, 84. [Google Scholar] [CrossRef]
- Bains, W. Many chemistries could be used to build living systems. Astrobiology 2004, 4, 137–167. [Google Scholar] [CrossRef]
- Ward, P.D.; Benner, S.A. Alien biochemistries. In Planets and Life; Sullivan, W.T., III, Baross, J.A., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 537–544. [Google Scholar]
- Feinberg, G.; Shapiro, R. Life beyond Earth-The Intelligent Earthling’s Guide to Life in the Universe; William Morrow and Company: New York, NY, USA, 1980; p. 464. [Google Scholar]
- Battersby, S. Could Alien Life Exist in the form of Dna-Shaped Dust? New Scientist. 2007. Available online: https://www.newscientist.com/article/dn12466-could-alien-life-exist-in-the-form-of-dna-shaped-dust/#ixzz6UlQmzkHm (accessed on 10 August 2020).
- Cairns-Smith, A.G. Genetic Takeover; Cambridge University Press: London, UK, 1982. [Google Scholar]
- Davis, W.L.; McKay, C.P. Origins of life: A comparison of theories and application to Mars. Orig. Life Evol. Biosph. 1996, 26, 61–73. [Google Scholar] [CrossRef]
- Miyakawa, S.; Joshi, P.C.; Gaffey, M.J.; Gonzalez-Toril, E.; Hyland, C.; Ross, T.; Rybij, K.; Ferris, J.P. Studies in the mineral and salt-catalyzed formation of RNA oligomers. Orig. Life Evol. Biosph. 2006, 36, 343–361. [Google Scholar] [CrossRef]
- Tsytovich, V.N.; Morfill, G.E.; Fortov, V.E.; Gusein-Zade, N.G.; Klumov, B.A.; Vladimirov, S.V. From plasma crystals and helical structures towards inorganic living matter. New J. Phys. 2007, 9, 263. [Google Scholar] [CrossRef]
- Wong, W.S.Y.; Li, M.; Nisbet, D.R.; Craig, V.S.J.; Wang, Z.; Tricoli, A. Mimosa Origami: A nanostructure-enabled directional self-organization regime of materials. Sci. Adv. 2016, 2, 1600417. [Google Scholar] [CrossRef] [Green Version]
- Percec, V.; Wilson, D.A.; Leowanawat, P.; Wilson, C.J.; Hughes, A.D.; Kaucher, M.S.; Hammer, D.A.; Levine, D.H.; Kim, A.J.; Bates, F.S.; et al. Self-assembly of Janus dendrimers into uniform dendrimersomes and other complex architectures. Science 2010, 328, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Evers, C.H.; Luiken, J.A.; Bolhuis, P.G.; Kegel, W.K. Self-assembly of microcapsules via colloidal bond hybridization and anisotropy. Nature 2016, 534, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Zion, M.Y.; He, X.; Maass, C.C.; Sha, R.; Seeman, N.C.; Chaikin, P.M. Self-assembled three-dimensional chiral colloidal architecture. Science 2017, 358, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Sagan, C.; Salpeter, E.E. Particles, environments, and possible ecologies in the jovian atmosphere. Astrophys. J. 1976, 32, 737–755. [Google Scholar] [CrossRef] [Green Version]
- Irwin, L.N. Exotic forms of life on other worlds. In Handbook of Exoplanets; Deeg, H.J., Belmonte, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–13. [Google Scholar]
- Schulze-Makuch, D.; Irwin, L.N. Life in the Universe: Expectations and Constraints, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Schulze-Makuch, D. 100 Million Planets in Our Galaxy May Harbor Complex Life; Air & Space Smithsonian: Washington, DC, USA, 2014. [Google Scholar]
- Kereszturi, A.; Noack, L. Review on the role of planetary factors on habitability. Orig. Life Evol. Biosph. 2016, 46, 473–486. [Google Scholar] [CrossRef]
- Chyba, C.F.; Hand, K.P. Planetary science-Life without photosynthesis. Science 2001, 292, 2026–2027. [Google Scholar] [CrossRef]
- Irwin, L.N.; Schulze-Makuch, D. Strategy for modeling putative multilevel ecosystems on Europa. Astrobiology 2003, 3, 813–821. [Google Scholar] [CrossRef]
- Chyba, C.F. Energy for microbial life on Europa. Nature 2000, 403, 381–382. [Google Scholar] [CrossRef]
- McCollom, T.M. Methanogenesis as a potential source of chemical energy for primary biomass production by autotrophic organisms in hydrothermal systems on Europa. J. Geophys. Res. Planets 1999, 104, 30729–30742. [Google Scholar] [CrossRef]
- Zolotov, M.Y.; Shock, E.L. Energy for biologic sulfate reduction in a hydrothermally formed ocean on Europa. J. Geophys. Res. Planets 2003, 108, 5022. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.; Grima, C.; Blankenship, D.D.; Young, D.A.; Soderlund, K.M. Mobilization of Near-Surface Brine on Europa in Lunar Planetary Laboratory; NASA: Houston, TX, USA, 2017.
- Van Dover, C.L.; Reynolds, G.T.; Chave, A.D.; Tyson, J.A. Light at deep sea hydrothermal vents. Geophys. Res. Lett. 1996, 23, 20149–22052. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Makuch, D.; Irwin, L.N. Alternative energy sources could support life on Europa. Eos Trans. Am. Geophys. Union 2001, 82, 150. [Google Scholar]
- Kargel, J. Mars-A Warmer, Wetter Planet; Praxis Publishing: Chichester, UK, 2004; p. 542. [Google Scholar]
- Kereszturi, A. Review of wet environment types on Mars with focus on duration and volumetric issues. Astrobiology 2012, 12, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Grotzinger, J.P.; Sumner, D.Y.; Kah, L.C.; Stack, K.; Gupta, S.; Edgar, L.; Rubin, D.; Lewis, K.; Schieber, J.; Mangold, N.; et al. A habitable fluvio-lacustrine environment at Yellowknife Bay, Gale crater, Mars. Science 2014, 343, 1242777. [Google Scholar] [CrossRef] [PubMed]
- Spiga, A.; Hinson, D.P.; Madeleine, J.-B.; Navarro, T.; Millour, E.; Forget, F.; Montmessin, F. Snow precipitation on Mars driven by cloud-induced night-time convection. Nat. Geosci. 2017, 10, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Möhlmann, D.T.; Formisano, V.; Savijrvi, H.; Wolkenberg, P.; Niemand, M. Fog phenomena on Mars. Planet. Space Sci. 2009, 57, 1987–1992. [Google Scholar] [CrossRef]
- Martín-Torres, F.J.; Zorzano, M.-P.; Valentín-Serrano, P.; Harri, A.-M.; Genzer, M.; Kemppinen, O.; Rivera-Valentin, E.G.; Jun, I.; Wray, J.; Bo Madsen, M.; et al. Transient liquid water and water activity at Gale crater on Mars. Nat. Geosci. 2015, 8, 357–361. [Google Scholar] [CrossRef]
- Pál, B.; Kereszturi, A. Annual and daily ideal periods for deliquescence at the landing site of InSight based on GCM model calculations. Icarus 2020, 340, 113639. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Wagner, D.; Kounaves, S.P.; Mangelsdorf, K.; Devine, K.G.; de Vera, J.P.; Schmitt-Kopplin, P.; Grossart, H.P.; Parro, V.; Kaupenjohann, M.; et al. Transitory microbial habitat in the hyperarid Atacama Desert. Proc. Natl. Acad. Sci. USA 2018, 115, 2670–2675. [Google Scholar] [CrossRef] [Green Version]
- Davila, A.F.; Hawes, I.; Ascaso, C.; Wierzchos, J. Salt deliquescence drives photosynthesis in the hyperarid Atacama Desert. Environ. Microbiol. Rep. 2013, 5, 583–587. [Google Scholar] [CrossRef]
- Maus, D.; Heinz, J.; Schirmack, J.; Airo, A.; Kounaves, S.P.; Wagner, D.; Schulze-Makuch, D. Methanogenic archaea can produce methane in deliquescence-driven Mars analog environments. Sci. Rep. 2020, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; McKay, P.; Omelon, C. Polar endoliths–An anti-correlation of climatic extremes and microbial biodiversity. Intl. J. Astrobiol. 2002, 1, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Wharton, D.A. The environmental physiology of Antarctic terrestrial nematodes: A review. J. Comp. Physiol. B 2003, 173, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Makuch, D.; Irwin, L.N.; Lipps, J.H.; LeMone, D.; Dohm, J.M.; Fairén, A.G. Scenarios for the evolution of life on Mars. J. Geophys. Res. Planets 2005, 110, E12S23. [Google Scholar] [CrossRef] [Green Version]
- Tokano, T.; McKay, C.P.; Neubauer, F.M.; Atreya, S.K.; Ferri, F.; Fulchignoni, M.; Niemann, H.B. Methane drizzle on Titan. Nature 2006, 442, 432–435. [Google Scholar] [CrossRef]
- Turse, C.; Leitner, J.; Firneis, M.; Schulze-Makuch, D. Simulations of Prebiotic Chemistry under Post-Impact Conditions on Titan. Life 2013, 3, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Horst, S.M.; Yelle, R.V.; Buch, A.; Carrasco, N.; Cernogora, G.; Dutuit, O.; Quirico, E.; Sciamma-O’Brien, E.; Smith, M.A.; Somogyi, A.; et al. Formation of amino acids and nucleotide bases in a Titan atmosphere simulation experiment. Astrobiology 2012, 12, 809–817. [Google Scholar] [CrossRef] [Green Version]
- Meckenstock, R.U.; von Netzer, F.; Stumpp, C.; Lueders, T.; Himmelberg, A.M.; Hertkorn, N.; Schmitt-Kopplin, P.; Harir, M.; Hosein, R.; Haque, S.; et al. Oil biodegradation. Water droplets in oil are microhabitats for microbial life. Science 2014, 345, 673–676. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Haque, S.; de Sousa Antonio, M.R.; Ali, D.; Hosein, R.; Song, Y.C.; Yang, J.; Zaikova, E.; Beckles, D.M.; Guinan, E.; et al. Microbial life in a liquid asphalt desert. Astrobiology 2011, 11, 241–258. [Google Scholar] [CrossRef]
- Sandstrom, H.; Rahm, M. Can polarity-inverted membranes self-assemble on Titan? Sci. Adv. 2020, 6, eaax0272. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, J.; Lunine, J.; Clancy, P. Membrane alternatives in worlds without oxygen: Creation of an azotosome. Sci. Adv. 2015, 1, e1400067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strigari, L.E.; Barnabè, M.; Marshall, P.J.; Blandford, R.D. Nomads of the Galaxy. Mon. Not. R. Astron. Soc. 2012, 423, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Delorme, P.; Gagné, J.; Malo, L.; Reylé, C.; Artigau, E.; Albert, L.; Forveille, T.; Delfosse, X.; Allard, F.; Homeier, D. CFBDSIR2149–0403: A 4–7 Jupiter-mass free-floating planet in the young moving group AB Doradus? Astron. Astrophys. 2012, 548, A26. [Google Scholar] [CrossRef] [Green Version]
- Lineweaver, C.H.; Fenner, Y.; Gibson, B.K. The galactic habitable zone and the age distribution of complex life in the Milky Way. Science 2004, 303, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Wada, K.; Tsukamoto, Y.; Kokubo, E. Planet formation around supermassive black holes in the active galactic nuclei. Astrophys. J. 2019, 886, 107. [Google Scholar] [CrossRef]
- Lingam, M.; Ginsburg, I.; Bialy, S. Active galactic nuclei: Boon or bane for biota? Astrophys. J. 2019, 877, 62. [Google Scholar] [CrossRef] [Green Version]
- Iorio, L. Effects of general relativistic spin precessions on the habitability of rogue planets orbiting supermassive black holes. Astrophys. J. 2020, 896, 82. [Google Scholar]
- Schnittman, J.D. Life on Miller’s Planet: The habitable zone around supermassive black holes. arXiv 2019, arXiv:1910.00940. [Google Scholar]
- Abbot, D.S.; Switzer, E.R. The Steppenwolf: A proposal for a habitable planet in interstellar space. Astrophys. J. Lett. 2011, 735, L27–L31. [Google Scholar] [CrossRef] [Green Version]
- Muller, A.W.; Schulze-Makuch, D. Thermal energy and the origin of life. Orig. Life Evol. Biosph. 2006, 36, 177–189. [Google Scholar] [CrossRef]
- Bada, J.L. State-of-the-art instruments for detecting extraterrestrial life. Proc. Natl. Acad. Sci. USA 2001, 98, 797–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, D.J. Life-sustaining planets in interstellar space? Nature 1999, 400, 32. [Google Scholar] [CrossRef] [PubMed]
- Seager, S.; Huang, J.; Petkowski, J.J.; Pajusalu, M. Laboratory studies on the viability of life in H2-dominated exoplanet atmospheres. Nat. Astron. 2020, 4, 802–806. [Google Scholar] [CrossRef]
- Zeng, L.; Sasselov, D.D.; Jacobsen, S.B. Mass-radius relation for rocky planets based on PREM. Astrophys. J. 2016, 819, 127. [Google Scholar] [CrossRef]
- Heller, R.; Armstrong, J. Superhabitable worlds. Astrobiology 2014, 14, 50–66. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Makuch, D.; Heller, R.; Guinan, E.F. In search for a planet better than Earth: Top contenders for a superhabitable world. Astrobiology 2020, in press. [Google Scholar]
- Guinan, E.F.; Engle, S.; Ribas, I.; Harper, G. FUSE observations of young to old dG, dK & dM stars: Critical tests of dynamos, X-FUV irradiances and impacts on planetary environments and the development of life. Bull. Am. Astron. Soc. 2005, 37, 1490. [Google Scholar]
- Luger, R.; Barnes, R. Extreme water loss and abiotic O2 buildup on planets throughout the habitable zones of M dwarfs. Astrobiology 2015, 15, 119–143. [Google Scholar] [CrossRef] [Green Version]
- Kasting, J.F.; Whitmire, D.P.; Reynolds, R.T. Habitable Zones around Main-Sequence Stars. Icarus 1993, 101, 108–128. [Google Scholar] [CrossRef]
- Heller, R.; Leconte, J.; Barnes, R. Tidal obliquity evolution of potentially habitable planets. Astron. Astrophys. 2011, 528, A27. [Google Scholar] [CrossRef] [Green Version]
- Lingam, M.; Loeb, A. Physical constraints on the likelihood of life on exoplanets. I17:116–126. Int. J. Astrobiol. 2018, 17, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Lingam, M.; Loeb, A. Is life most likely around Sun-like stars? J. Cosmol. Astropart. Phys. 2019, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Barnes, R. Tidal locking of habitable exoplanets. Celest. Mech. Dynam. Astron. 2017, 129, 509–536. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.M.; Haberle, R.M.; Reynolds, R.T. Simulations of the atmospheres of synchronously rotating terrestrial planets orbiting M dwarfs: Conditions for atmospheric collapse and the implications for habitability. Icarus 1997, 129, 450–465. [Google Scholar] [CrossRef] [Green Version]
- Tarter, J.C.; Backus, P.R.; Mancinelli, R.L.; Aurnou, J.M.; Backman, D.E.; Basri, G.S.; Boss, A.P.; Clarke, A.; Deming, D.; Doyle, L.R.; et al. A reappraisal of the habitability of planets around M dwarf stars. Astrobiology 2007, 7, 30–65. [Google Scholar] [CrossRef]
- Edson, A.R.; Kasting, J.F.; Pollard, D.; Lee, S.; Bannon, P.R. The carbonate-silicate cycle and CO2/climate feedbacks on tidally locked terrestrial planets. Astrobiology 2012, 12, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Wandel, A.; Gale, J. The bio-habitable zone and atmospheric properties for planets of red dwarfs. Int. J. Astrobiol. 2020, 19, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Boutle, I.A.; Joshi, M.; Lambert, F.H.; Mayne, N.J.; Lyster, D.; Manners, J.; Ridgway, R.; Kohary, K. Mineral dust increases the habitability of terrestrial planets but confounds biomarker detection. Nat. Commun. 2020, 11, 2731. [Google Scholar] [CrossRef]
- Checlair, J.H.; Salazar, A.M.; Paradise, A.; Menou, K.; Abbot, D.S. No Snowball Cycles at the Outer Edge of the Habitable Zone for Habitable Tidally Locked Planets. Astrophys. J. 2019, 887, L3–L8. [Google Scholar] [CrossRef]
- Checlair, J.; Menou, K.; Abbot, D.S. No Snowball on Habitable Tidally Locked Planets. Astrophys. J. 2017, 845, 132. [Google Scholar] [CrossRef]
- Vaitilingom, M.; Attard, E.; Gaiani, N.; Sancelme, M.; Deguillaume, L.; Flossmann, A.I.; Amato, P.; Delort, A.-M. Long-term features of cloud microbiology at the puy de Dôme (France). Atmosph. Environ. 2012, 56, 88–100. [Google Scholar] [CrossRef]
- Bryan, N.C.; Christner, B.C.; Guzik, T.G.; Granger, D.J.; Steward, M.F. Abundance and survival of microbial aerosols in the troposphere and stratosphere. ISME J. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Amato, P.; Joly, M.; Besaury, L.; Oudart, A.; Taib, N.; Mone, A.I.; Deguillaume, L.; Delort, A.M.; Debroas, D. Active microorganisms thrive among extremely diverse communities in cloud water. PLoS ONE 2017, 12, e0182869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, P.; Besaury, L.; Joly, M.; Penaud, B.; Deguillaume, L.; Delort, A.M. Metatranscriptomic exploration of microbial functioning in clouds. Sci. Rep. 2019, 9, 4383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seager, S.; Petkowski, J.J.; Gao, P.; Bains, W.; Bryan, N.C.; Ranjan, S.; Greaves, J. The Venusian lower atmosphere haze as a depot for desiccated microbial life: A proposed life cycle for persistence of the Venusian aerial biosphere. Astrobiology 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Burrows, S.M.; Butler, T.; Jöckel, P.; Tost, H.; Kerkweg, A.; Pöschl, U.; Lawrence, M.G. Bacteria in the global atmosphere-Part 2: Modeling of emissions and transport between different ecosystems. Atmosph. Chem. Phys. 2009, 9, 9281–9297. [Google Scholar] [CrossRef] [Green Version]
- Way, M.J.; Del Genio, A.D.; Kiang, N.Y.; Sohl, L.E.; Grinspoon, D.H.; Aleinov, I.; Kelley, M.; Clune, T. Was Venus the first habitable world of our solar system? Geophys Res. Lett. 2016, 43, 8376–8383. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Irwin, L.N.; Fairén, A.G. Drastic environmental change and its effects on a planetary biosphere. Icarus 2013, 225, 275–280. [Google Scholar] [CrossRef]
- Grinspoon, D.H. Venus Revealed: A New Look Below the Clouds of Our Mysterious Twin Planet; Perseus Publishing: Cambridge, MA, USA, 1997. [Google Scholar]
- Limaye, S.S.; Mogul, R.; Smith, D.J.; Ansari, A.H.; Slowik, G.P.; Vaishampayan, P. Venus’ Spectral Signatures and the Potential for Life in the Clouds. Astrobiology 2018, 18, 1181–1198. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Grinspoon, D.H.; Abbas, O.; Irwin, L.N.; Bullock, M.A. A sulfur-based survival strategy for putative phototrophic life in the venusian atmosphere. Astrobiology 2004, 4, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Franck, S.; Block, A.; von Bloh, W.; Bounama, C.; Garrido, I.; Schellnhuber, H.J. Planetary habitability: Is Earth commonplace in the Milky Way? Naturwissenschaften 2001, 88, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.H. The Fermi Paradox Is neither Fermi’s nor a paradox. Astrobiology 2015, 15, 195–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, S. Where Is Everybody? Fifty Solutions to the Fermi Paradox and the Problem of Extraterrestrial Life; Copernicus: New York, NY, USA, 2002. [Google Scholar]
- Ward, P.D.; Brownlee, D. Rare Earth: Why Complex. Life Is Uncommon in the Universe; Springer: New York, NY, USA, 2000; p. 315. [Google Scholar]
- Ward, P.D. The Medea Hypothesis: Is Life on Earth Ultimately Self-Destructive? Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- NASA. Mars 2020 Perseverance Rover. 2020. Available online: https://www.jpl.nasa.gov/missions/mars-2020-perseverance-rover/ (accessed on 30 July 2020).
- Kereszturi, A.; Bradak, B.; Chatzitheodoridis, E.; Ujvari, G. Indicators and methods to understand past environments from ExoMars rover drills. Orig. Life Evol. Biosph. 2016, 46, 435–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairen, A.G.; Davila, A.F.; Lim, D.; Bramall, N.; Bonaccorsi, R.; Zavaleta, J.; Uceda, E.R.; Stoker, C.; Wierzchos, J.; Dohm, J.M.; et al. Astrobiology through the ages of Mars: The study of terrestrial analogues to understand the habitability of Mars. Astrobiology 2010, 10, 821–843. [Google Scholar] [CrossRef]
- Popa, R.; Smith, A.R.; Popa, R.; Boone, J.; Fisk, M. Olivine-respiring bacteria isolated from the rock-ice interface in a lava-tube cave, a Mars analog environment. Astrobiology 2012, 12, 9–18. [Google Scholar] [CrossRef]
- Sandford, S.A.; Aleon, J.; Alexander, C.M.; Araki, T.; Bajt, S.; Baratta, G.A.; Borg, J.; Bradley, J.P.; Brownlee, D.E.; Brucato, J.R.; et al. Organics captured from comet 81P/Wild 2 by the Stardust spacecraft. Science 2006, 314, 1720–1724. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Makuch, D.; Irwin, L.N. Reassessing the possibility of life on Venus: Proposal for an astrobiology mision. Astrobiology 2002, 2, 197–202. [Google Scholar] [CrossRef]
- Jones, R.M.; Goordial, J.M.; Orcutt, B.N. Low energy subsurface environments as extraterrestrial analogs. Front. Microbiol. 2018, 9, 1605. [Google Scholar] [CrossRef]
- Naganuma, T. Search for life in deep biospheres. Biol. Sci. Space 2003, 17, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Gleeson, D.F.; Pappalardo, R.T.; Anderson, M.S.; Grasby, S.E.; Mielke, R.E.; Wright, K.E.; Templeton, A.S. Biosignature detection at an Arctic analog to Europa. Astrobiology 2012, 12, 135–150. [Google Scholar] [CrossRef]
- Prieto-Ballesteros, O.; Rodriguez, N.; Kargel, J.S.; Kessler, C.G.; Amils, R.; Remolar, D.F. Tirez lake as a terrestrial analog of Europa. Astrobiology 2003, 3, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Klenner, F.; Postberg, F.; Hillier, J.; Khawaja, N.; Cable, M.L.; Abel, B.; Kempf, S.; Glein, C.R.; Lunine, J.I.; Hodyss, R.; et al. Discriminating abiotic and biotic fingerprints of amino acids and fatty acids in ice grains relevant to ocean worlds. Astrobiology 2020, 20, 12. [Google Scholar]
- Nadeau, J.; Lindensmith, C.; Deming, J.W.; Fernandez, V.I.; Stocker, R. Microbial morphology and motility as biosignatures for outer planet missions. Astrobiology 2016, 16, 755–774. [Google Scholar] [CrossRef] [PubMed]
- McKay, C.P.; Smith, H.D. Possibilities for methanogenic life in liquid methane on the surface of Titan. Icarus 2005, 178, 274–276. [Google Scholar] [CrossRef] [Green Version]
- Khare, B.N.; Bakes, E.L.; Cruikshank, D.; McKay, C.P. Solid organic matter in the atmosphere and on the surface of outer Solar System bodies. Adv. Space Res. 2001, 27, 299–307. [Google Scholar] [CrossRef]
- Spilker, L. Cassini-Huygens’ exploration of the Saturn system: 13 years of discovery. Science 2019, 364, 1046–1051. [Google Scholar] [CrossRef]
- Bohn, R.B.; Sandford, S.A.; Allamandola, L.J.; Cruikshank, D.P. Infrared spectroscopy of Triton and Pluto ice analogs: The case for saturated hydrocarbons. Icarus 1994, 111, 151–173. [Google Scholar] [CrossRef]
- Agnor, C.B.; Hamilton, D.P. Neptune’s capture of its moon Triton in a binary-planet gravitational encounter. Nature 2006, 441, 192–194. [Google Scholar] [CrossRef]
- Kerr, R.A. Planetary science. How wet the moon? Just damp enough to be interesting. Science 2010, 330, 434. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Crawford, I. Was there an early habitability window for our Moon? Astrobiology 2018, 18, 985–988. [Google Scholar] [CrossRef] [Green Version]
- Cowan, N.B. The exoplanet opportunity: Top-down planetary science. Eos Trans. Am. Geophys. Union 2014, 95, 209–210. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Guinan, E. Another Earth 2.0? Not So Fast. Astrobiology 2016, 16, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Gaidos, E. Detecting the glint of starlight on the oceans of distant planets. Icarus 2008, 195, 927–937. [Google Scholar] [CrossRef] [Green Version]
- Maccone, C. Deep Space Flight and Communications: Exploiting the Sun as a Gravitational Lens; Springer-Praxis: Chichester, UK, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Habitat | Solar System Examples | Life Forms Possible 1 | Observational Possibilities with Current Technologies | Experimental Possibilities |
|---|---|---|---|---|
| Rocky/water surface with dense N2/O2 atmosphere | Earth | Micro/macro individuals Amorphous inorganic Amorphous organic | Direct, current, ongoing | Direct, current, ongoing |
| “Super Earth” (larger and warmer than current Earth) | None | Micro/macro individuals Amorphous inorganic Amorphous organic | Remote Analog habitats on Earth | Lab simulations Studies of analogs |
| Barren rocky, with previous water, no or little atmosphere | Mars | Micro/macro individuals Amorphous inorganic Amorphous organic | Robotic Human exploration Analog habitats on Earth | Analysis of return samples In situ instrument analysis Lab simulations Studies of analogs |
| Barren rocky, no previous water, with little or no atmosphere | Mercury | Amorphous inorganic | Robotic | Lab simulations |
| Ice-covered global ocean | Europa Ceres (?) Enceladus | Micro/macro individuals Amorphous inorganic Amorphous organic | Analog habitats on Earth Robotic | Studies of analogs In situ instrument analysis Lab simulations |
| Rocky/hydrocarbon surface with dense N2/CH4 atmosphere | Titan | Micro/macro individuals with exotic biochemistry Amorphous inorganic Amorphous organic | Robotic | In situ instrument analysis Lab simulations Studies of analogs |
| Rogue planet with various surfaces and atmospheres | Triton (ancestrally) | Dependent on surface, atmosphere, and planetary history | Very remote | Lab simulations |
| Tidally locked with planet or Red Dwarf | Moon, Io | Dependent on surface, atmosphere, and history | Direct human exploration of Moon Robotic on Io Remote for exoplanets | Analysis of sample return from Moon Study of analog habitats for Io Lab simulations |
| Gas giant or rocky, with dense atmosphere | Gas giants, Venus | Microbial Amorphous gaseous | Robotic Spectral signatures | Robotic probe of gas giants Sample return from clouds of Venus |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irwin, L.N.; Schulze-Makuch, D. The Astrobiology of Alien Worlds: Known and Unknown Forms of Life. Universe 2020, 6, 130. https://doi.org/10.3390/universe6090130
Irwin LN, Schulze-Makuch D. The Astrobiology of Alien Worlds: Known and Unknown Forms of Life. Universe. 2020; 6(9):130. https://doi.org/10.3390/universe6090130
Chicago/Turabian StyleIrwin, Louis N., and Dirk Schulze-Makuch. 2020. "The Astrobiology of Alien Worlds: Known and Unknown Forms of Life" Universe 6, no. 9: 130. https://doi.org/10.3390/universe6090130
APA StyleIrwin, L. N., & Schulze-Makuch, D. (2020). The Astrobiology of Alien Worlds: Known and Unknown Forms of Life. Universe, 6(9), 130. https://doi.org/10.3390/universe6090130