Reconsideration of the Semaphorin-3A Binding Motif Found in Chondroitin Sulfate Using Galnac4s-6st-Knockout Mice

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Enzyme-Linked Immunosorbent Assay
2.2. Animals
2.3. Real-Time PCR Analysis
2.4. Immunohistochemistry
2.5. Immunoblotting
2.6. Statistical Analysis
3. Results
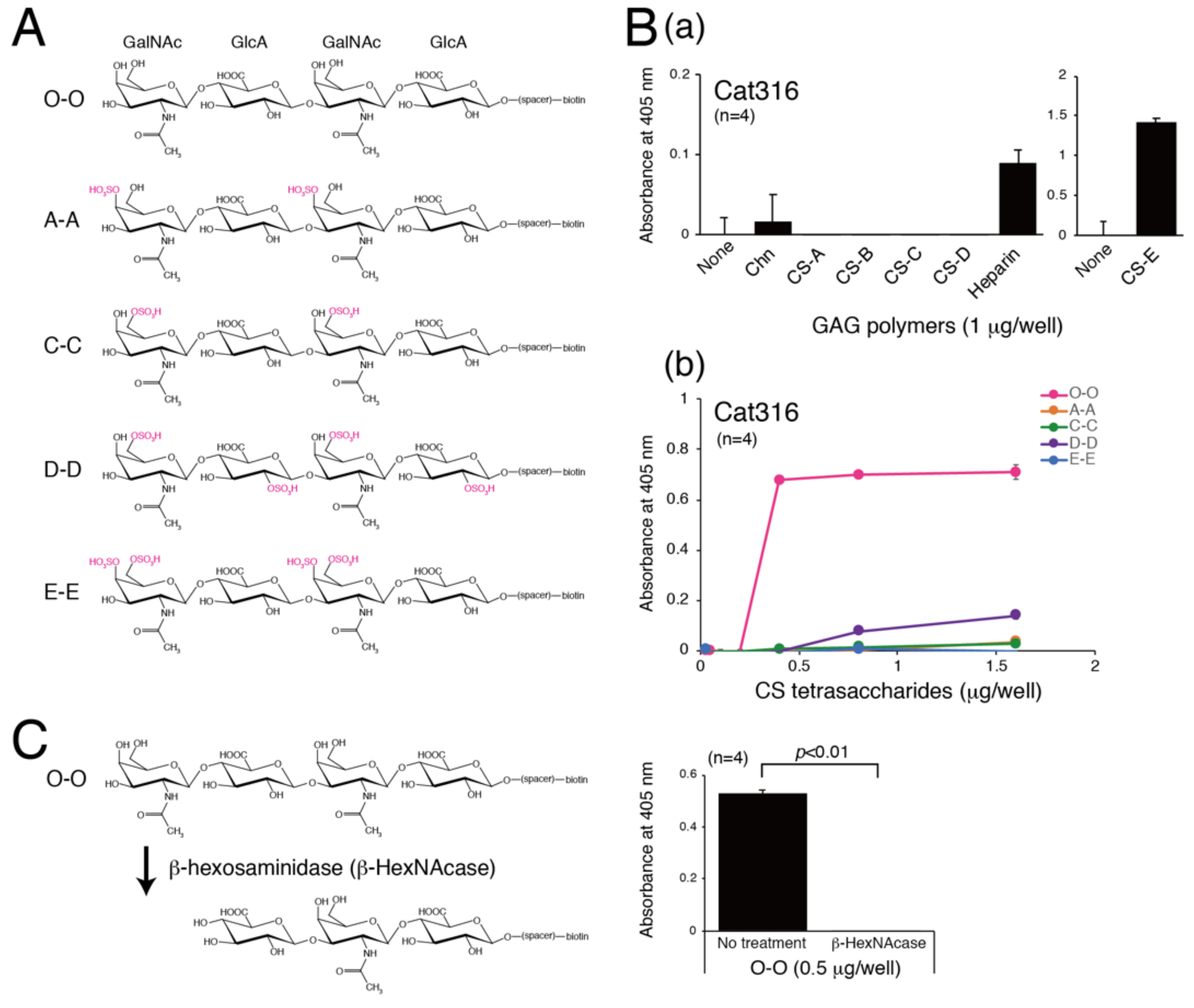
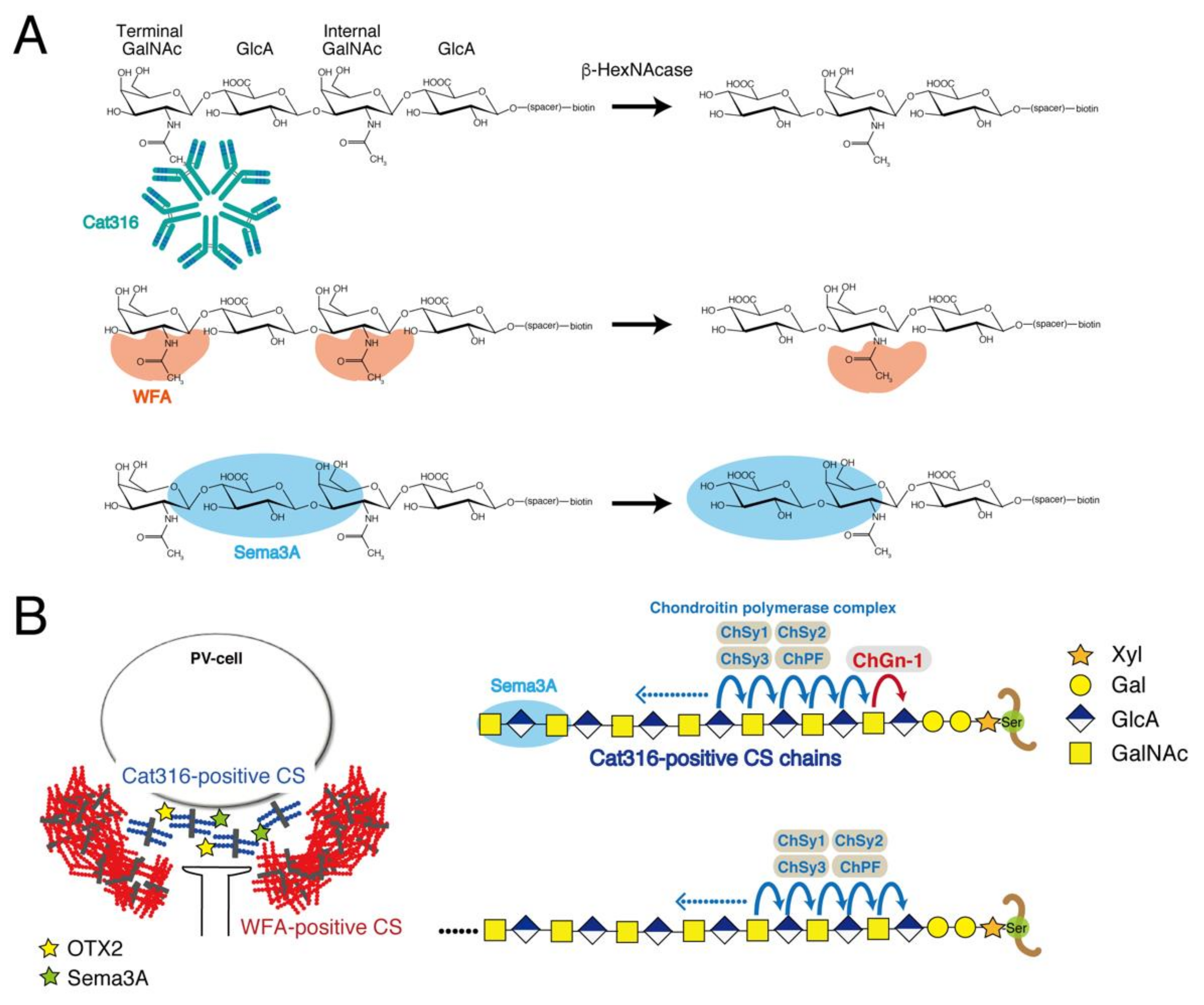
3.1. Determination of the Epitope of the Cat316 Antibody
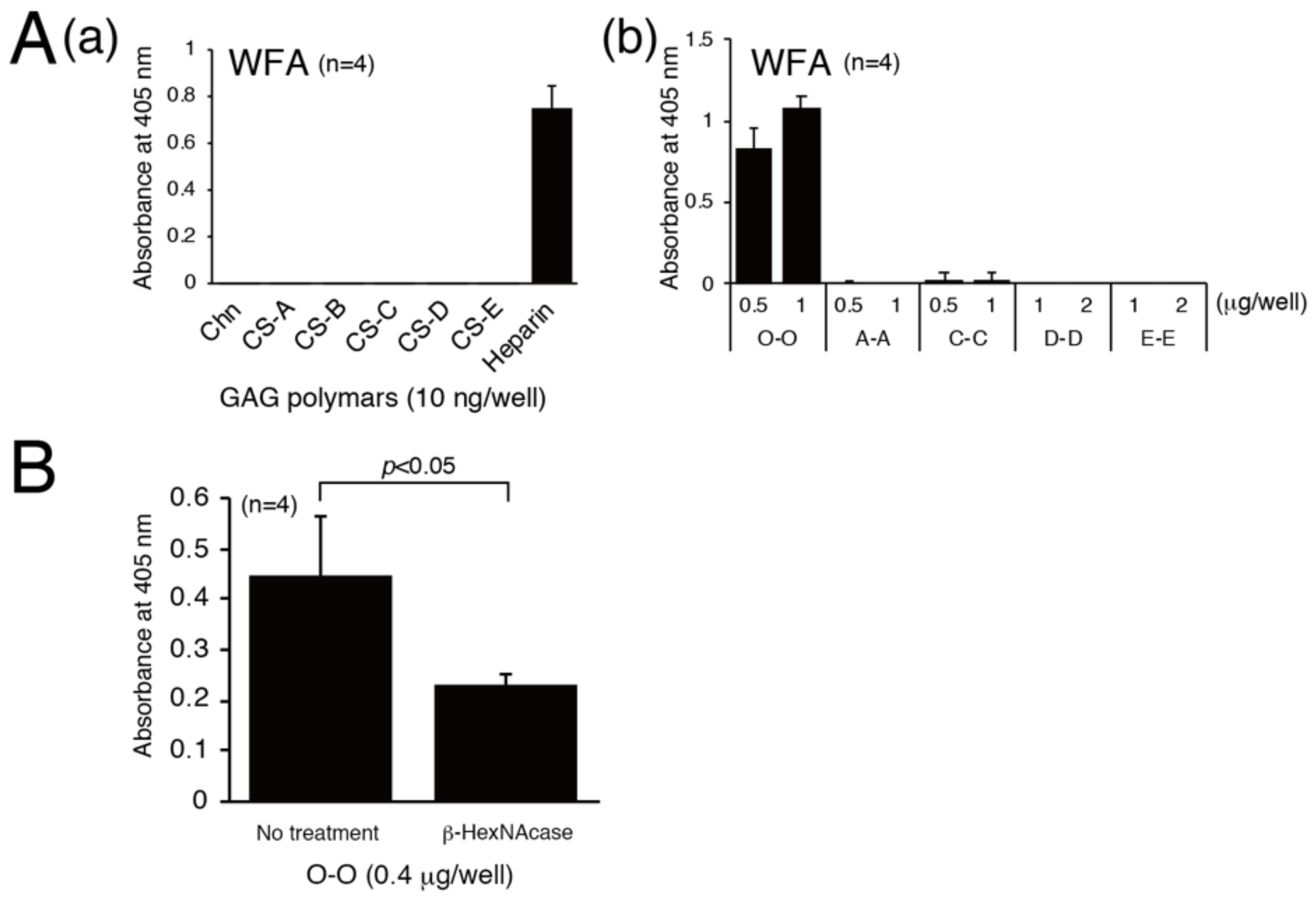
3.2. Determination of Structural Requirements for Recognition by WFA
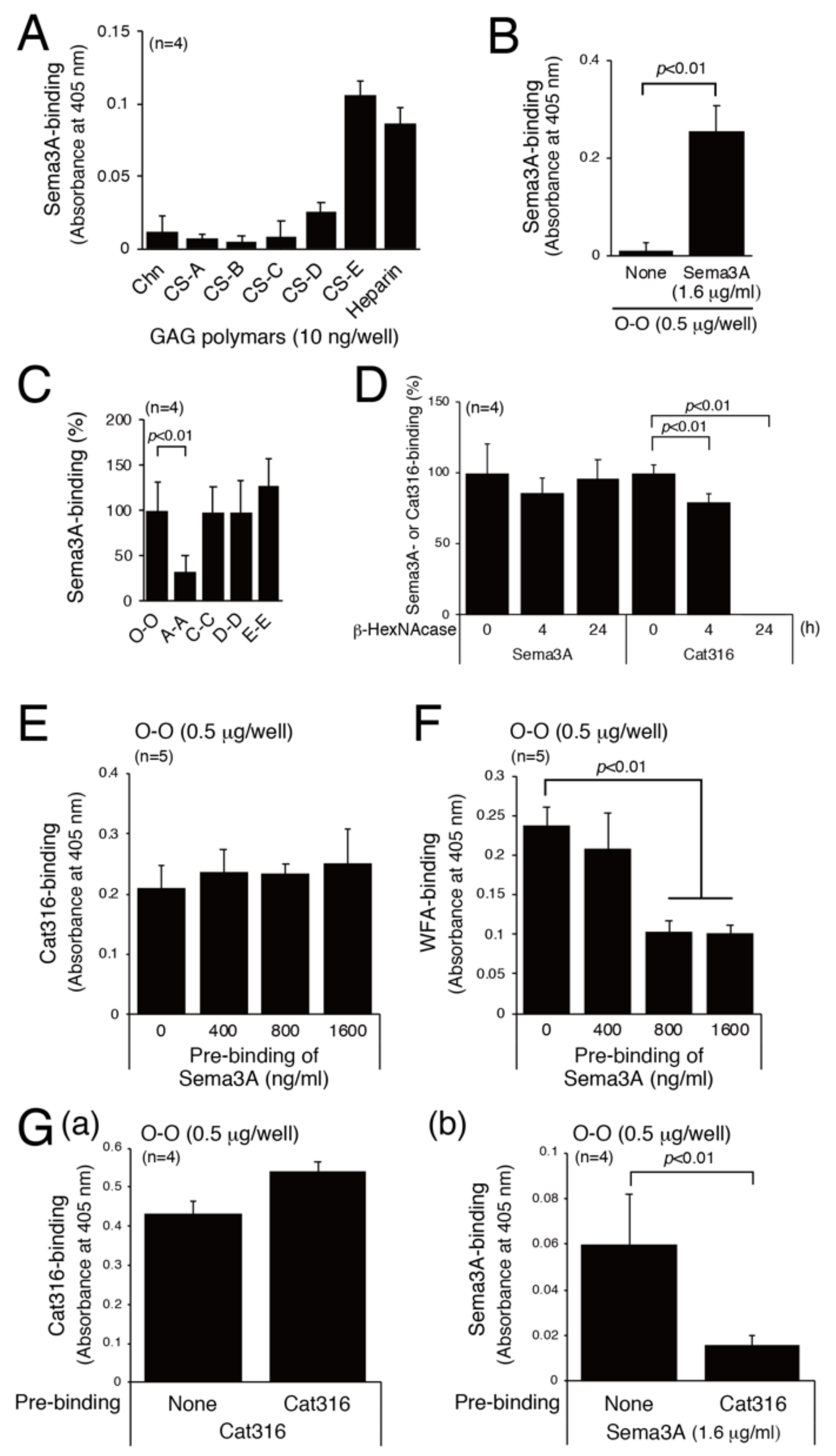
3.3. Examination of the Sema3A-Binding Motif
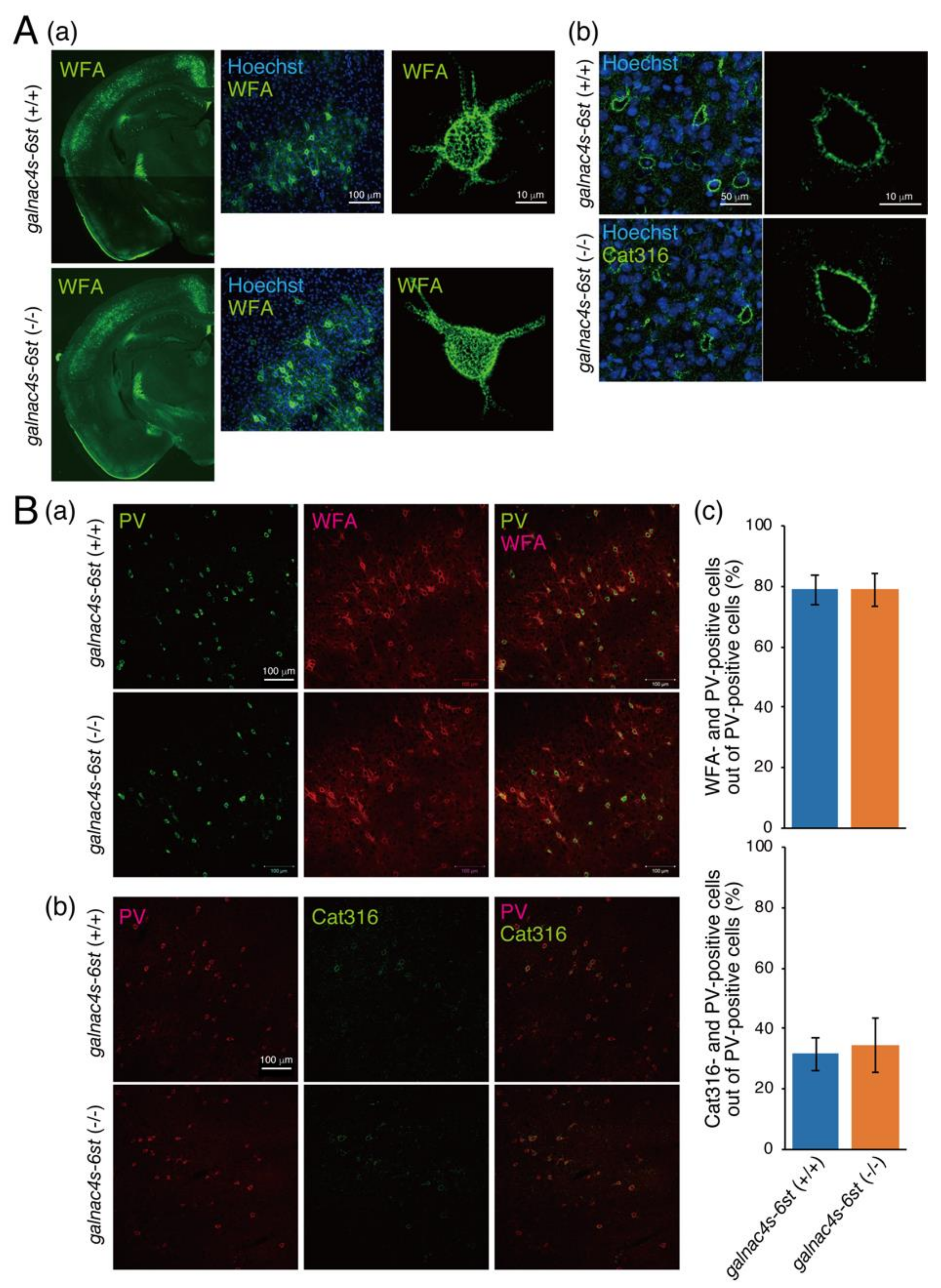
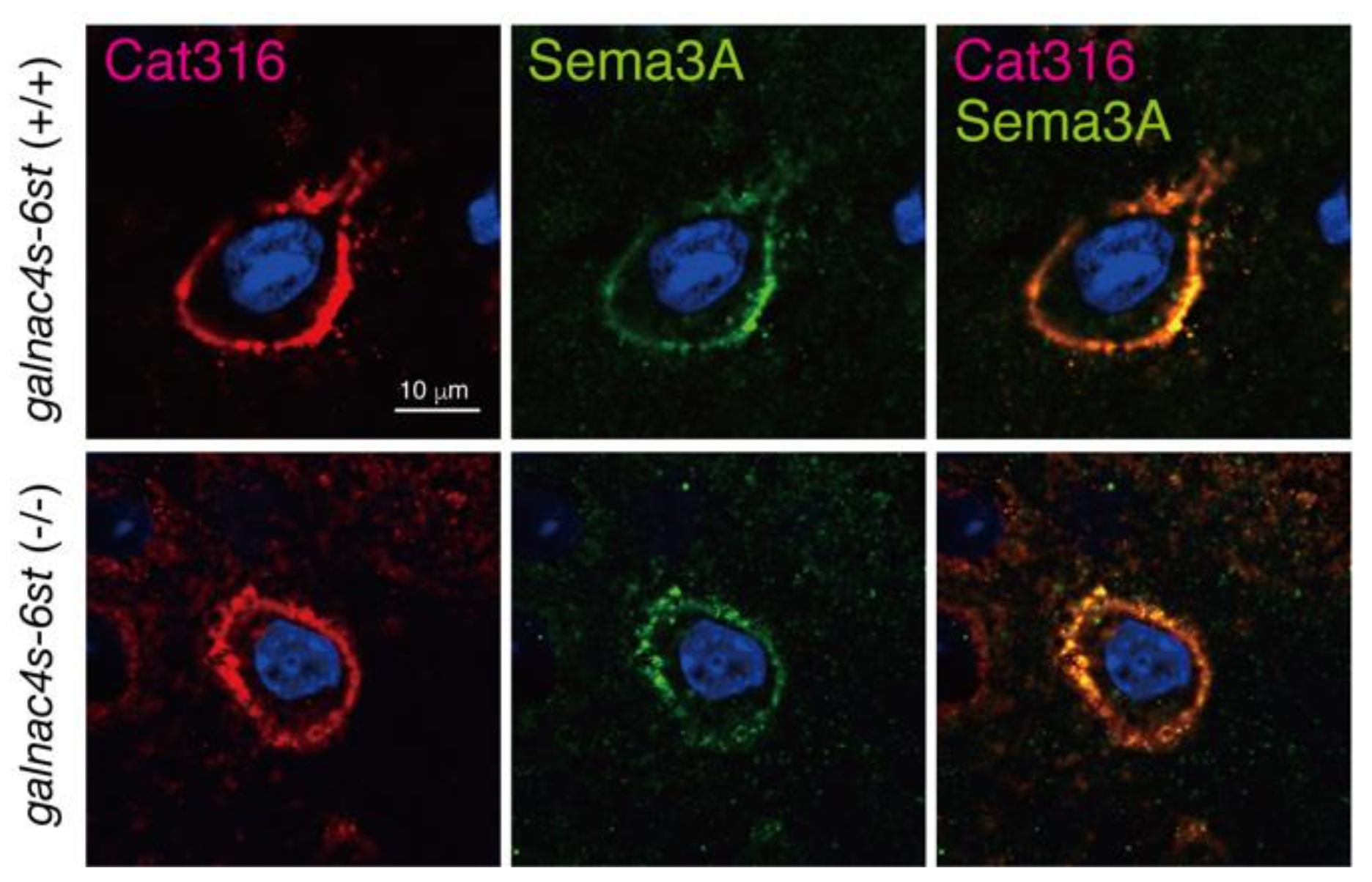
3.4. Analysis of Physiological Significance of the Binding of Sema3A to CS-E
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sigal, Y.M.; Bae, H.; Bogart, L.J.; Hensch, T.K.; Zhuang, X. Structural maturation of cortical perineuronal nets and their perforating synapses revealed by superresolution imaging. Proc. Natl. Acad. Sci. USA 2019, 116, 7071–7076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takesian, A.E.; Hensch, T.K. Balancing Plasticity/Stability Across Brain Development. Prog. Brain Res. 2013, 207, 3–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Kirkwood, A.; Pizzorusso, T.; Porciatti, V.; Morales, B.; Bear, M.F.; Maffei, L.; Tonegawa, S. BDNF Regulates the Maturation of Inhibition and the Critical Period of Plasticity in Mouse Visual Cortex. Cell 1999, 98, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyaya, B.; Di Cristo, G.; Higashiyama, H.; Knott, G.W.; Kuhlman, S.J.; Welker, E.; Huang, Z.J. Experience and Activity-Dependent Maturation of Perisomatic GABAergic Innervation in Primary Visual Cortex during a Postnatal Critical Period. J. Neurosci. 2004, 24, 9598–9611. [Google Scholar] [CrossRef] [PubMed]
- Hensch, T.K. Critical period plasticity in local cortical circuits. Nat. Rev. Neurosci. 2005, 6, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.W.; Maffei, L. Reactivation of Ocular Dominance Plasticity in the Adult Visual Cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef] [Green Version]
- Berardi, N.; Pizzorusso, T.; Maffei, L. Extracellular Matrix and Visual Cortical Plasticity. Neuron 2004, 44, 905–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichelt, A.C.; Hare, D.J.; Bussey, T.J.; Saksida, L.M. Perineuronal Nets: Plasticity, Protection, and Therapeutic Potential. Trends Neurosci. 2019, 42, 458–470. [Google Scholar] [CrossRef]
- Baroncelli, L.; Sale, A.; Viegi, A.; Vetencourt, J.F.M.; De Pasquale, R.; Baldini, S.; Maffei, L. Experience-dependent reactivation of ocular dominance plasticity in the adult visual cortex. Exp. Neurol. 2010, 226, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.R.; Gao, W.-J. PV Interneurons: Critical Regulators of E/I Balance for Prefrontal Cortex-Dependent Behavior and Psychiatric Disorders. Front. Neural Circuits 2018, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Marín, O. Developmental timing and critical windows for the treatment of psychiatric disorders. Nat. Med. 2016, 22, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Pantazopoulos, H.; Woo, T.-U.W.; Lim, M.P.; Lange, N.; Berretta, S. Extracellular Matrix-Glial Abnormalities in the Amygdala and Entorhinal Cortex of Subjects Diagnosed With Schizophrenia. Arch. Gen. Psychiatry 2010, 67, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, S.; Kitagawa, H. Mechanisms for modulation of neural plasticity and axon regeneration by chondroitin sulphate. J. Biochem. 2014, 157, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Yoshioka, N.; Tsukano, H.; Sakai, A.; Miyata, S.; Watanabe, Y.; Yanagawa, Y.; Sakimura, K.; Takeuchi, K.; Kitagawa, H.; et al. Chondroitin Sulfate Is Required for Onset and Offset of Critical Period Plasticity in Visual Cortex. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef]
- Nadanaka, S.; Ishida, M.; Ikegami, M.; Kitagawa, H. Chondroitin 4-O-Sulfotransferase-1 Modulates Wnt-3a Signaling through Control of E Disaccharide Expression of Chondroitin Sulfate. J. Biol. Chem. 2008, 283, 27333–27343. [Google Scholar] [CrossRef] [Green Version]
- Nadanaka, S.; Kinouchi, H.; Kitagawa, H. Chondroitin sulfate–mediated N-cadherin/β-catenin signaling is associated with basal-like breast cancer cell invasion. J. Biol. Chem. 2017, 293, 444–465. [Google Scholar] [CrossRef] [Green Version]
- Nadanaka, S.; Hashiguchi, T.; Kitagawa, H. Aberrant glycosaminoglycan biosynthesis by tumor suppressor EXTL2 deficiency promotes liver inflammation and tumorigenesis through Toll-like 4 receptor signaling. Faseb J. 2020, 34, 8385–8401. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.; Tan, C.L.; Alves, J.N.; Ehlert, E.M.E.; Miller, G.M.; Hsieh-Wilson, L.C.; Sugahara, K.; Oosterhof, A.; Van Kuppevelt, T.H.; Verhaagen, J.; et al. Semaphorin 3A Binds to the Perineuronal Nets via Chondroitin Sulfate Type E Motifs in Rodent Brains. J. Biol. Chem. 2013, 288, 27384–27395. [Google Scholar] [CrossRef] [Green Version]
- Beurdeley, M.; Spatazza, J.; Lee, H.H.C.; Sugiyama, S.; Bernard, C.; Di Nardo, A.A.; Hensch, T.K.; Prochiantz, A. Otx2 Binding to Perineuronal Nets Persistently Regulates Plasticity in the Mature Visual Cortex. J. Neurosci. 2012, 32, 9429–9437. [Google Scholar] [CrossRef] [Green Version]
- De Winter, F.; Kwok, J.C.F.; Fawcett, J.W.; Vo, T.T.; Carulli, D.; Verhaagen, J. The Chemorepulsive Protein Semaphorin 3A and Perineuronal Net-Mediated Plasticity. Neural Plast. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Hilton, S.; Alves, J.N.; Saksida, L.; Bussey, T.; Matthews, R.T.; Kitagawa, H.; Spillantini, M.G.; Kwok, J.; Fawcett, J.W. Antibody recognizing 4-sulfated chondroitin sulfate proteoglycans restores memory in tauopathy-induced neurodegeneration. Neurobiol. Aging 2017, 59, 197–209. [Google Scholar] [CrossRef]
- Tamura, J.-I.; Tsutsumishita-Nakai, N.; Nakao, Y.; Kawano, M.; Kato, S.; Takeda-Okuda, N.; Nadanaka, S.; Kitagawa, H. Synthesis and interaction with midkine of biotinylated chondroitin sulfate tetrasaccharides. Bioorg. Med. Chem. Lett. 2012, 22, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Nakata, T.; Takeda-Okuda, N.; Nadanaka, S.; Kitagawa, H.; Tamura, J.-I. Synthesis of chondroitin sulfate CC and DD tetrasaccharides and interactions with 2H6 and LY111. Bioorg. Med. Chem. 2018, 26, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Nadanaka, S.; Purunomo, E.; Takeda, N.; Tamura, J.-I.; Kitagawa, H. Heparan Sulfate Containing Unsubstituted Glucosamine Residues. J. Biol. Chem. 2014, 289, 15231–15243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtake-Niimi, S.; Kondo, S.; Ito, T.; Kakehi, S.; Ohta, T.; Habuchi, H.; Kimata, K.; Habuchi, O. Mice Deficient inN-Acetylgalactosamine 4-Sulfate 6-O-Sulfotransferase Are Unable to Synthesize Chondroitin/Dermatan Sulfate containingN-Acetylgalactosamine 4,6-Bissulfate Residues and Exhibit Decreased Protease Activity in Bone Marrow-derived Mast Cells. J. Biol. Chem. 2010, 285, 20793–20805. [Google Scholar] [CrossRef] [Green Version]
- Miyata, S.; Nadanaka, S.; Igarashi, M.; Kitagawa, H. Structural Variation of Chondroitin Sulfate Chains Contributes to the Molecular Heterogeneity of Perineuronal Nets. Front. Integr. Neurosci. 2018, 12, 3. [Google Scholar] [CrossRef]
- Matthews, R.T.; Kelly, G.M.; Zerillo, C.A.; Gray, G.; Tiemeyer, M.; Hockfield, S. Aggrecan Glycoforms Contribute to the Molecular Heterogeneity of Perineuronal Nets. J. Neurosci. 2002, 22, 7536–7547. [Google Scholar] [CrossRef]
- Miyata, S.; Komatsu, Y.; Yoshimura, Y.; Taya, C.; Kitagawa, H. Persistent cortical plasticity by upregulation of chondroitin 6-sulfation. Nat. Neurosci. 2012, 15, 414–422. [Google Scholar] [CrossRef]
- Tasic, B.; Yao, Z.; Graybuck, L.T.; Smith, K.A.; Nguyen, T.N.; Bertagnolli, D.; Goldy, J.; Garren, E.; Economo, M.N.; Viswanathan, S.; et al. Shared and distinct transcriptomic cell types across neocortical areas. Nat. Cell Biol. 2018, 563, 72–78. [Google Scholar] [CrossRef]
- Giamanco, K.; Morawski, M.; Matthews, R. Perineuronal net formation and structure in aggrecan knockout mice. Neuroscience 2010, 170, 1314–1327. [Google Scholar] [CrossRef]
- Vo, T.; Ecarulli, D.; Ehlert, E.M.; Kwok, J.C.; Dick, G.; Mecollari, V.; Moloney, E.B.; Neufeld, G.; De Winter, F.; Fawcett, J.W.; et al. The chemorepulsive axon guidance protein semaphorin3A is a constituent of perineuronal nets in the adult rodent brain. Mol. Cell. Neurosci. 2013, 56, 186–200. [Google Scholar] [CrossRef]
- Ueno, H.; Fujii, K.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Aoki, S.; Okamoto, M.; Ishihara, T.; Takao, K. Expression of aggrecan components in perineuronal nets in the mouse cerebral cortex. IBRO Rep. 2018, 4, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, M.; Yada, T.; Sato, T.; Akashima, T.; Iwasaki, H.; Mochizuki, H.; Inaba, N.; Togayachi, A.; Kudo, T.; Watanabe, H.; et al. Molecular Cloning and Characterization of a Novel Chondroitin Sulfate Glucuronyltransferase That Transfers Glucuronic Acid toN-Acetylgalactosamine. J. Biol. Chem. 2002, 277, 38179–38188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyama, T.; Kitagawa, H.; Ji, J.-I.T.; Sugahara, K. Molecular Cloning and Expression of Human ChondroitinN-Acetylgalactosaminyltransferase. J. Biol. Chem. 2002, 277, 8841–8846. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Takeuchi, K.; Onaga, S.H.; Sato, M.; Tsujita, M.; Abe, M.; Natsume, R.; Li, M.; Furuichi, T.; Saeki, M.; et al. Chondroitin sulfate N-acetylgalactosaminyltransferase-1 is required for normal cartilage development. Biochem. J. 2010, 432, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, Y.; Meng, H.; Du, J.; Luan, G.; Wang, H.-Q.; Yang, M.; Luo, Z. Role of miR-155 in the regulation of MMP-16 expression in intervertebral disc degeneration. J. Orthop. Res. 2017, 35, 1323–1334. [Google Scholar] [CrossRef] [Green Version]
- Sztrolovics, R.; Alini, M.; Roughley, P.J.; Mort, J.S. Aggrecan degradation in human intervertebral disc and articular cartilage. Biochem. J. 1997, 326, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, S.M.; Razak, K.; Ethell, I.M. A delicate balance: Role of MMP-9 in brain development and pathophysiology of neurodevelopmental disorders. Front. Cell. Neurosci. 2015, 9, 280. [Google Scholar] [CrossRef] [Green Version]
- Ripke, S.; O’Dushlaine, C.; Chambert, K.; Moran, J.L.; Kähler, A.K.; Akterin, S.; Bergen, S.E.; Collins, A.L.; Crowley, J.J.; Fromer, M.; et al. Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat. Genet. 2013, 45, 1150–1159. [Google Scholar] [CrossRef]
- Lepeta, K.; Kaczmarek, L. Matrix Metalloproteinase-9 as a Novel Player in Synaptic Plasticity and Schizophrenia. Schizophr. Bull. 2015, 41, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.C.; Tang, A.T.; Detter, M.R.; Choi, J.P.; Wang, R.; Yang, X.; Guerrero, A.A.; Wittig, C.F.; Hobson, N.; Girard, R.; et al. Cerebral cavernous malformations are driven by ADAMTS5 proteolysis of versican. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Miyata, S.; Kitagawa, H. Chondroitin 6-Sulfation Regulates Perineuronal Net Formation by Controlling the Stability of Aggrecan. Neural Plast. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratta, M.A.; Tortorella, M.D.; Arner, E.C. Age-related Changes in Aggrecan Glycosylation Affect Cleavage by Aggrecanase. J. Biol. Chem. 2000, 275, 39096–39102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struglics, A.; Larsson, S.; Pratta, M.; Kumar, S.; Lark, M.; Lohmander, L.; Lohmander, L.S. Human osteoarthritis synovial fluid and joint cartilage contain both aggrecanase- and matrix metalloproteinase-generated aggrecan fragments. Osteoarthr. Cartil. 2006, 14, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Takagi, H.; Asano, Y.; Yamakawa, N.; Matsumoto, I.; Kimata, K. Annexin 6 is a putative cell surface receptor for chondroitin sulfate chains. J. Cell Sci. 2002, 115, 3309–3318. [Google Scholar]
- Ishitsuka, R.; Kojima, K.; Utsumi, H.; Ogawa, H.; Matsumoto, I. Glycosaminoglycan Binding Properties of Annexin IV, V, and VI. J. Biol. Chem. 1998, 273, 9935–9941. [Google Scholar] [CrossRef] [Green Version]
- Horlacher, T.; Noti, C.; De Paz, J.L.; Bindschädler, P.; Hecht, M.-L.; Smith, D.F.; Fukuda, M.N.; Seeberger, P.H. Characterization of Annexin A1 Glycan Binding Reveals Binding to Highly Sulfated Glycans with Preference for Highly Sulfated Heparan Sulfate and Heparin. Biochemistry 2011, 50, 2650–2659. [Google Scholar] [CrossRef] [Green Version]
- Grindheim, A.K.; Saraste, J.; Vedeler, A. Protein phosphorylation and its role in the regulation of Annexin A2 function. Biochim. Biophys. Gen. Subj. 2017, 1861, 2515–2529. [Google Scholar] [CrossRef]
- Zhao, W.; Lu, B. Expression of annexin A2 in GABAergic interneurons in the normal rat brain. J. Neurochem. 2006, 100, 1211–1223. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay ID | Reporter Dye | Context Sequence | Gene Symbol | Gene Name | Species | NCBI Gene Reference |
|---|---|---|---|---|---|---|
| Mm00545794_m1 | FAM | 5′-CTCAGAAGAAGTTCCAGACCATGAC-3′ | Acan | aggrecan | Mus musculus | NM_007424.2 L07049.1 |
| Mm00476090_m1 | FAM | 5′-TCTACTGCTTCCGAGACTCTGCCCA-3′ | Bcan | brevican | Mus musculus | NM_007529.2 NM_001109758.1 X87096.1 BC052032.1 AK144405.1 |
| Mm00484007_m1 | FAM | 5′-AGGAAGATCCCACAGATCCCTGCGA-3′ | Ncan | neurocan | Mus musculus | NM_007789.3 X84727.1 BC065118.1 |
| Mm01283063_m1 | FAM | 5′-CACTGGCTGTGGATGGTGTTGTGTT-3′ | Vcan | versican | Mus musculus | NM_172955.1 NM_001134474.1 NM_001134475.1 NM_019389.2 NM_001081249.1 D28599.1 D32040.1 D16263.1 BC021652.1 AK014525.1 AK034871.1 BC096495.1 AK135324.1 AK163927.1 AK165069.1 |
| Mm00478484_m1 | FAM | 5′-GTCCTACACAGGAGCACTAAATCAA-3′ | Ptprz1 (Pcan) | protein tyrosine phosphatase, receptor type Z, polypeptide 1 (Phosphacan) | Mus musculus | NM_001081306.1 AJ133130.1 AK081698.1 BC157965.1 BC151071.1 |
| Mm01179833_m1 | FAM | 5′-AAGGAGGTGCTCGTAGCCGTGATCG-3′ | Sdc3 | syndecan 3 | Mus musculus | NM_011520.3 BC054795.1 AK129153.1 BC062093.1 AK150120.1 AK155127.1 |
| Mm99999915_g1 | FAM | 5′-GGTGTGAACGGATTTGGCCGTATTG-3′ | Gapdh | glyceraldehyde-3-phosphate dehydrogenase | Mus musculus | NM_008084.2 M32599.1 BC082592.1 BC083079.1 BC083080.1 BC083065.1 BC083149.1 BC085315.1 BC085274.1 BC085275.1 AK191938.1 AK192040.1 AK201736.1 BC091768.1 BC092252.1 BC092264.1 BC092294.1 BC093508.1 BU5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadanaka, S.; Miyata, S.; Yaqiang, B.; Tamura, J.-i.; Habuchi, O.; Kitagawa, H. Reconsideration of the Semaphorin-3A Binding Motif Found in Chondroitin Sulfate Using Galnac4s-6st-Knockout Mice. Biomolecules 2020, 10, 1499. https://doi.org/10.3390/biom10111499
Nadanaka S, Miyata S, Yaqiang B, Tamura J-i, Habuchi O, Kitagawa H. Reconsideration of the Semaphorin-3A Binding Motif Found in Chondroitin Sulfate Using Galnac4s-6st-Knockout Mice. Biomolecules. 2020; 10(11):1499. https://doi.org/10.3390/biom10111499
Chicago/Turabian StyleNadanaka, Satomi, Shinji Miyata, Bai Yaqiang, Jun-ichi Tamura, Osami Habuchi, and Hiroshi Kitagawa. 2020. "Reconsideration of the Semaphorin-3A Binding Motif Found in Chondroitin Sulfate Using Galnac4s-6st-Knockout Mice" Biomolecules 10, no. 11: 1499. https://doi.org/10.3390/biom10111499
APA StyleNadanaka, S., Miyata, S., Yaqiang, B., Tamura, J. -i., Habuchi, O., & Kitagawa, H. (2020). Reconsideration of the Semaphorin-3A Binding Motif Found in Chondroitin Sulfate Using Galnac4s-6st-Knockout Mice. Biomolecules, 10(11), 1499. https://doi.org/10.3390/biom10111499