Rheumatoid Arthritis: Applicability of Ready-to-Use Human Cartilaginous Cells for Screening of Compounds with TNF-Alpha Inhibitory Activity



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation and Culturing of Immature Cartilaginous Cells
2.3. TNF-α Stimulation
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. MTT Viability Test
2.6. Cryopreservation
2.7. Thawing of Cryopreserved Ready-to-Use Cells
2.8. Statistical Analysis
3. Results
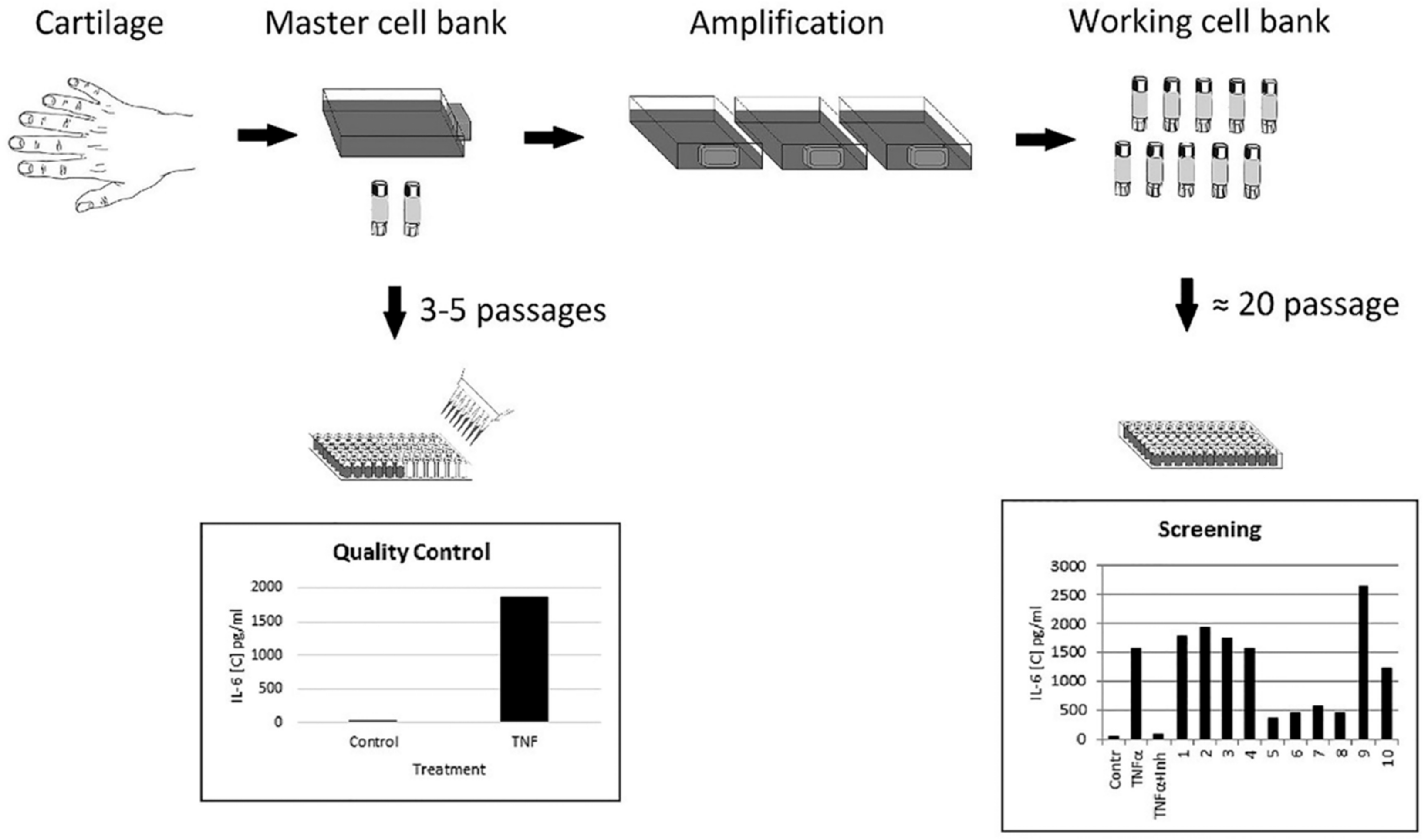
3.1. Cartilage Tissue Specimen Obtained from Amputated Supernumerary Digits of Juvenile Donors Is an Attractive Source of Cartilaginous Cells for In Vitro Assays
3.2. Response of Immature Cartilaginous Cells to TNF-α Stimulation
3.3. TNF-α Inhibitor Etanercept Inhibits IL-6 Expression in a Dose-Dependent Manner
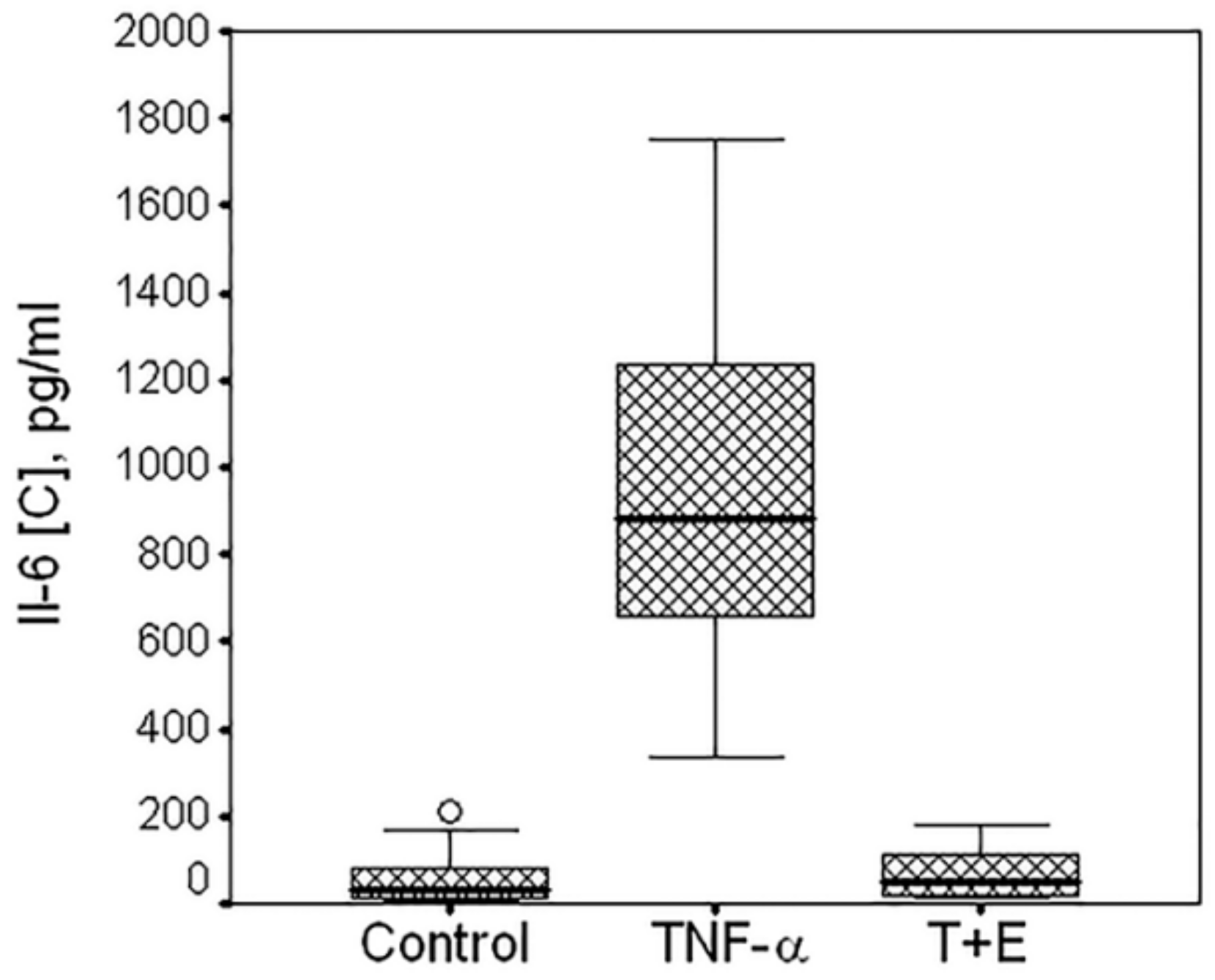
3.4. Evaluation of TNF-α-Mediated Effects on the Cells Obtained from 10 Donors
3.5. Optimization of Assay
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Willrich, M.A.V.; Murray, D.L.; Snyder, M.R. Tumor necrosis factor inhibitors: Clinical utility in autoimmune diseases. Transl. Res. 2015, 165, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Sedger, L.M.; McDermott, M.F. TNF and TNF-receptors: From mediators of cell death and inflammation to therapeutic giants–past, present and future. Cytokine Growth Factor Rev. 2014, 25, 453–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melagraki, G.; Ntougkos, E.; Rinotas, V.; Papaneophytou, C.; Leonis, G.; Mavromoustakos, T.; Kontopidis, G.; Douni, E.; Afantitis, A.; Kollias, G. Cheminformatics-aided discovery of small-molecule Protein-Protein Interaction (PPI) dual inhibitors of Tumor Necrosis Factor (TNF) and Receptor Activator of NF-κB Ligand (RANKL). PLoS Comput. Biol. 2017, 13, e1005372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Tweedie, D.; Beedie, S.L.; Vargesson, N.; Figg, W.D.; Greig, N.H.; Scerba, M.T. Design, synthesis and biological assessment of N-adamantyl, substituted adamantyl and noradamantyl phthalimidines for nitrite, TNF-α and angiogenesis inhibitory activities. Bioorganic Med. Chem. 2018, 26, 1547–1559. [Google Scholar] [CrossRef]
- Eglen, R.; Reisine, T. Primary Cells and Stem Cells in Drug Discovery: Emerging Tools for High-Throughput Screening. Assay Drug Dev. Technol. 2010, 9, 108–124. [Google Scholar] [CrossRef]
- Podgurskaya, A.D.; Tsvelaya, V.A.; Slotvitsky, M.M.; Dementyeva, E.V.; Valetdinova, K.R.; Agladze, K.I. The Use of iPSC-Derived Cardiomyocytes and Optical Mapping for Erythromycin Arrhythmogenicity Testing. Cardiovasc. Toxicol. 2019, 19, 518–528. [Google Scholar] [CrossRef]
- Horvath, P.; Aulner, N.; Bickle, M.; Davies, A.M.; Del Nery, E.; Ebner, D.; Montoya, M.C.; Östling, P.; Pietiäinen, V.; Price, L.S.; et al. Screening out irrelevant cell-based models of disease. Nat. Rev. Drug Discov. 2016, 15, 751–769. [Google Scholar] [CrossRef] [Green Version]
- Goldbard, S. Bringing primary cells to mainstream drug development and drug testing. Curr. Opin. Drug Discov. Dev. 2006, 9, 110–116. [Google Scholar]
- Flick, D.A.; Gifford, G.E. Comparison of in vitro cell cytotoxic assays for tumor necrosis factor. J. Immunol. Methods 1984, 68, 167–175. [Google Scholar] [CrossRef]
- Humphreys, D.T.; Wilson, M.R. Modes of L929 cell death induced by TNF-α and other cytotoxic agents. Cytokine 1999, 11, 773–782. [Google Scholar] [CrossRef]
- Khabar, K.S.A.; Siddiqui, S.; Armstrong, J.A. WEHI-13VAR: A stable and sensitive variant of WEHI 164 clone 13 fibrosarcoma for tumor necrosis factor bioassay. Immunol. Lett. 1995, 46, 107–110. [Google Scholar] [CrossRef]
- Espevik, T.; Nissen-Meyer, J. A highly sensitive cell line, WEHI 164 clone 13, for measuring cytotoxic factor/tumor necrosis factor from human monocytes. J. Immunol. Methods 1986, 95, 99–105. [Google Scholar] [CrossRef]
- Shealy, D.J.; Cai, A.; Staquet, K.; Baker, A.; Lacy, E.R.; Johns, L.; Vafa, O.; Gunn, G.; Tam, S.; Sague, S.; et al. Characterization of golimumab, a human monoclonal antibody specific for human tumor necrosis factor α. MAbs 2010, 2, 428–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallemand, C.; Kavrochorianou, N.; Steenholdt, C.; Bendtzen, K.; Ainsworth, M.A.; Meritet, J.-F.; Blanchard, B.; Lebon, P.; Taylor, P.; Charles, P.; et al. Reporter gene assay for the quantification of the activity and neutralizing antibody response to TNFα antagonists. J. Immunol. Methods 2011, 373, 229–239. [Google Scholar] [CrossRef]
- Siegel, S.A.; Shealy, D.J.; Nakada, M.T.; Le, J.; Woulfe, D.S.; Probert, L.; Kollias, G.; Ghrayeb, J.; Vilcek, J.; Daddona, P.E. The mouse/human chimeric monoclonal antibody ca2 neutralizes tnf in vitro and protects transgenic mice from cachexia and tnf lethality in vivo. Cytokine 1995, 7, 15–25. [Google Scholar] [CrossRef]
- Barlič, A.; Žigon, S.; Blejec, A.; Velikonja, N.K. Gene expression of cultured human chondrocytes as a model for assessing neutralization efficacy of soluble TNFα by TNFα antagonists. Biologicals 2015, 43, 171–180. [Google Scholar] [CrossRef]
- Žigon-Branc, S.; Jeras, M.; Blejec, A.; Barlič, A. Applicability of human osteoarthritic chondrocytes for in vitro efficacy testing of anti-TNFα drugs. Biologicals 2017, 45, 96–101. [Google Scholar] [CrossRef]
- Lories, R.J.U.; Derese, I.; De Bari, C.; Luyten, F.P. Evidence for uncoupling of inflammation and joint remodeling in a mouse model of spondylarthritis. Arthritis Rheum. 2007, 56, 489–497. [Google Scholar] [CrossRef]
- Asquith, D.L.; Miller, A.M.; McInnes, I.B.; Liew, F.Y. Animal models of rheumatoid arthritis. Eur. J. Immunol. 2009, 39, 2040–2044. [Google Scholar] [CrossRef]
- Karlsen, T.A.; Shahdadfar, A.; Brinchmann, J.E. Human Primary Articular Chondrocytes, Chondroblasts-Like Cells, and Dedifferentiated Chondrocytes: Differences in Gene, MicroRNA, and Protein Expression and Phenotype. Tissue Eng. Part C Methods 2010, 17, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B. Chondrogenesis, chondrocyte differentiation, and articular cartilage metabolism in health and osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Somoza, R.A.; Welter, J.F.; Correa, D.; Caplan, A.I. Chondrogenic Differentiation of Mesenchymal Stem Cells: Challenges and Unfulfilled Expectations. Tissue Eng. Part B Rev. 2014, 20, 596–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grogan, S.P.; Duffy, S.F.; Pauli, C.; Koziol, J.A.; Su, A.I.; D’Lima, D.D.; Lotz, M. Zone-specific gene expression patterns in articular cartilage. Arthritis Rheum. 2013, 65, 418–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLise, A.M.; Fischer, L.; Tuan, R.S. Cellular interactions and signaling in cartilage development. Osteoarthr. Cartil. 2000, 8, 309–334. [Google Scholar] [CrossRef] [Green Version]
- Kozhemyakina, E.; Lassar, A.B.; Zelzer, E. A pathway to bone: Signaling molecules and transcription factors involved in chondrocyte development and maturation. Development 2015, 142, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Santoro, A.; Conde, J.; Scotece, M.; Abella, V.; Lopez, V.; Pino, J.; Gomez, R.; Gómez-Reino, J.J.; Gualillo, O. Choosing the right chondrocyte cell line: Focus on nitric oxide. J. Orthop. Res. 2015, 33, 1784–1788. [Google Scholar] [CrossRef]
- Darling, E.M.; Athanasiou, K.A. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. J. Orthop. Res. 2005, 23, 425–432. [Google Scholar] [CrossRef]
- Hamada, T.; Sakai, T.; Hiraiwa, H.; Nakashima, M.; Ono, Y.; Mitsuyama, H.; Ishiguro, N. Surface markers and gene expression to characterize the differentiation of monolayer expanded human articular chondrocytes. Nagoya J. Med. Sci. 2013, 75, 101–111. [Google Scholar]
- Mortazavi, F.; Shafaei, H.; Soleimani Rad, J.; Rushangar, L.; Montaceri, A.; Jamshidi, M. High Quality of Infant Chondrocytes in Comparison with Adult Chondrocytes for Cartilage Tissue Engineering. World J. Plast. Surg. 2017, 6, 183–189. [Google Scholar]
- Lin, Z.; Fitzgerald, J.B.; Xu, J.; Willers, C.; Wood, D.; Grodzinsky, A.J.; Zheng, M. Gene expression profiles of human chondrocytes during passaged monolayer cultivation. J. Orthop. Res. 2008, 26, 1230–1237. [Google Scholar] [CrossRef]
- Zaman, G.J.R.; DeRoos, J.; Blomenrohr, M.; Vankoppen, C.; Oosterom, J. Cryopreserved cells facilitate cell-based drug discovery. Drug Discov. Today 2007, 12, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Cawkill, D.; Eaglestone, S.S. Evolution of cell-based reagent provision. Drug Discov. Today 2007, 12, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Harris, D. Biobanking and Regenerative Medicine: An Overview. J. Clin. Med. 2018, 7, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donors | Control 1 | TNF-α 2 | TNF-α+E 3 | pANOVA | p1-2 | p1-3 | p2-3 |
|---|---|---|---|---|---|---|---|
| D1 | 154.29 ± 4.32 | 371.70 ± 26.91 | 158.15 ± 17.14 | <0.001 | 0.001 | 0.935 | <0.001 |
| D2 | 47.20 ± 31.48 | 649.21 ± 11.28 | 70.70 ± 25.06 | <0.001 | <0.001 | 0.750 | 0.012 |
| D3 | 49.85 ± 23.68 | 714.87 ± 84.26 | 85.84 ± 95.79 | 0.005 | 0.074 | 0.929 | 0.043 |
| D4 | 15.24 ± 0.11 | 772.67 ± 94.69 | 31.70 ± 10.06 | <0.001 | 0.011 | 0.212 | 0.011 |
| D5 | 8.24 ± 0.23 | 867.10 ± 214.78 | 18.77 ± 7.14 | 0.001 | 0.042 | 0.251 | 0.043 |
| D6 | 14.55 ± 1.20 | 861.39 ± 124.85 | 14.78 ± 3.78 | <0.001 | 0.015 | 0.999 | 0.015 |
| D7 | 75.77 ± 9.91 | 1117.99 ± 9.42 | 88.33 ± 33.93 | <0.001 | 0.045 | 0.933 | 0.020 |
| D8 | 189.68 ± 32.68 | 1320.40 ± 139.69 | 53.09 ± 8.54 | 0.001 | 0.077 | 0.154 | 0.081 |
| D9 | 38.10 ± 6.07 | 1323.68 ± 81.07 | 57.36 ± 27.39 | <0.001 | 0.003 | 0.633 | 0.001 |
| D10 | 4.11 ± 0.07 | 1483.77 ± 250.76 | 14.84 ± 1.86 | <0.001 | 0.020 | 0.020 | 0.020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volova, L.T.; Pugachev, E.I.; Rossinskaya, V.V.; Boltovskaya, V.V.; Dolgushkin, D.A.; Ossina, N. Rheumatoid Arthritis: Applicability of Ready-to-Use Human Cartilaginous Cells for Screening of Compounds with TNF-Alpha Inhibitory Activity. Biomolecules 2020, 10, 1563. https://doi.org/10.3390/biom10111563
Volova LT, Pugachev EI, Rossinskaya VV, Boltovskaya VV, Dolgushkin DA, Ossina N. Rheumatoid Arthritis: Applicability of Ready-to-Use Human Cartilaginous Cells for Screening of Compounds with TNF-Alpha Inhibitory Activity. Biomolecules. 2020; 10(11):1563. https://doi.org/10.3390/biom10111563
Chicago/Turabian StyleVolova, Larissa T., Evgeniy I. Pugachev, Victoria V. Rossinskaya, Violetta V. Boltovskaya, Dmitry A. Dolgushkin, and Natalya Ossina. 2020. "Rheumatoid Arthritis: Applicability of Ready-to-Use Human Cartilaginous Cells for Screening of Compounds with TNF-Alpha Inhibitory Activity" Biomolecules 10, no. 11: 1563. https://doi.org/10.3390/biom10111563
APA StyleVolova, L. T., Pugachev, E. I., Rossinskaya, V. V., Boltovskaya, V. V., Dolgushkin, D. A., & Ossina, N. (2020). Rheumatoid Arthritis: Applicability of Ready-to-Use Human Cartilaginous Cells for Screening of Compounds with TNF-Alpha Inhibitory Activity. Biomolecules, 10(11), 1563. https://doi.org/10.3390/biom10111563