Tim17 Updates: A Comprehensive Review of an Ancient Mitochondrial Protein Translocator


Abstract
:1. Introduction
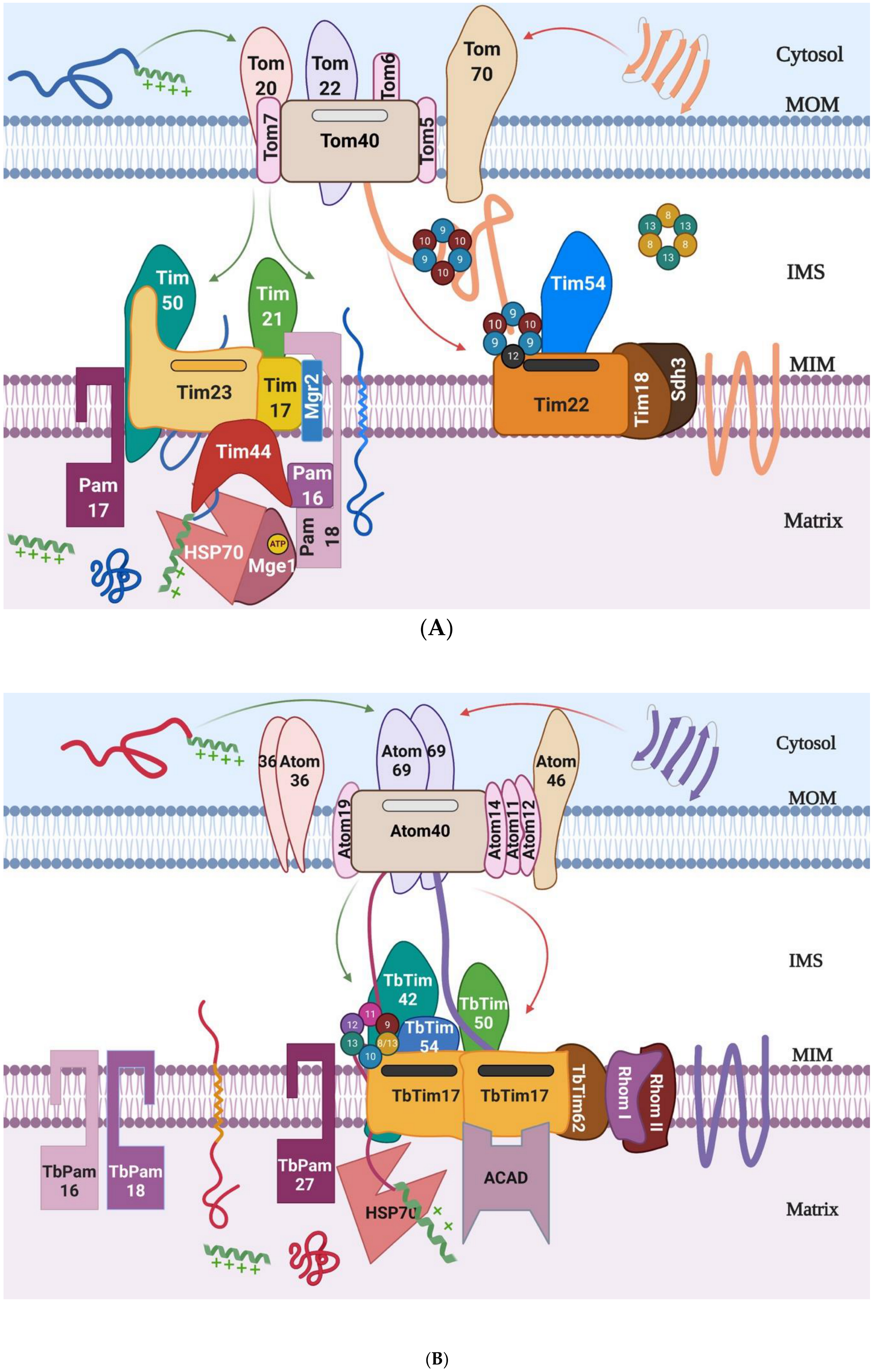
2. Mitochondrial Protein Import Machinery
3. Prevalence of Tim17 and Tim17 Homologues Among Eukaryotes
4. Tim17 and Tim17-Containing Protein Complexes in Different Eukaryotes
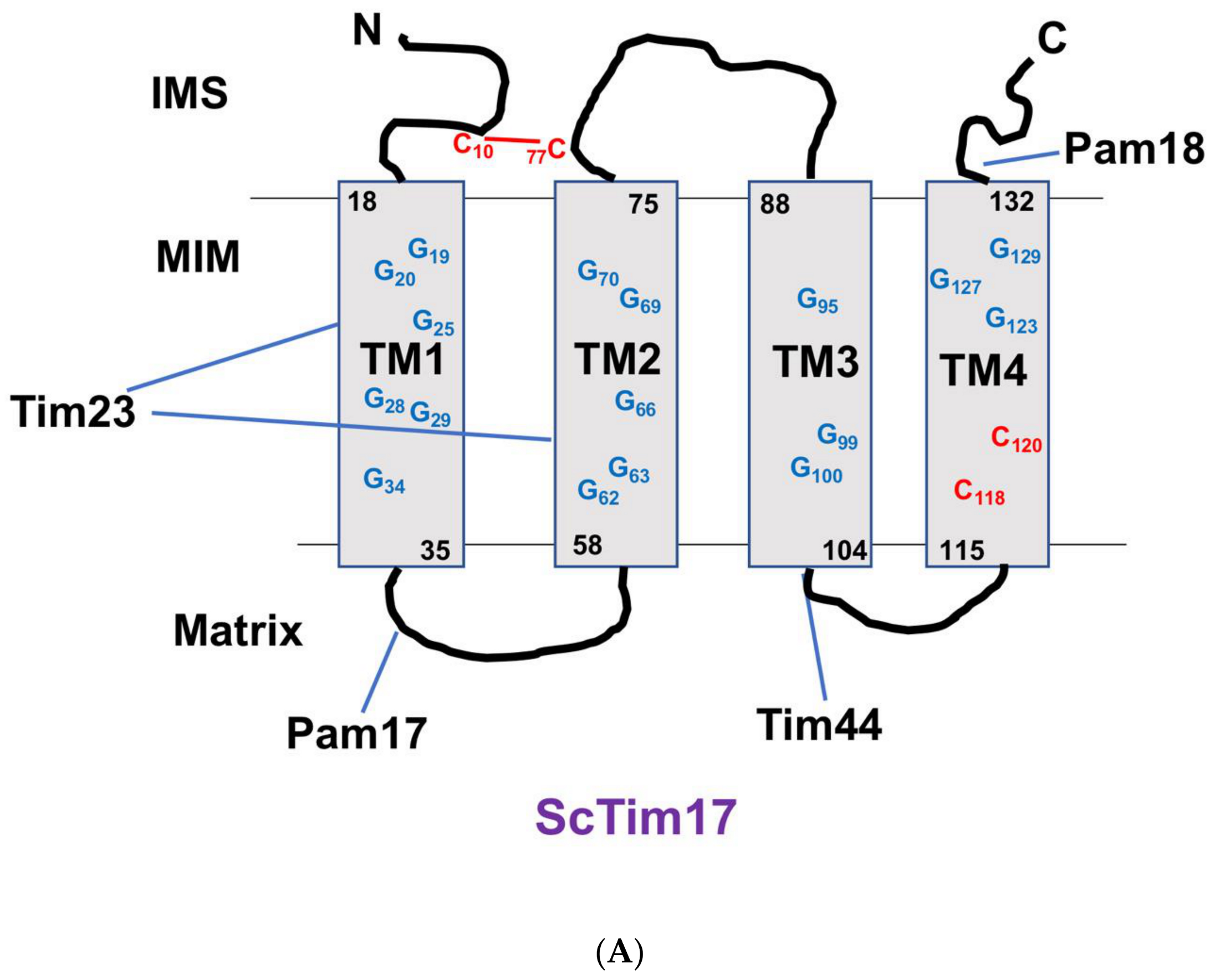
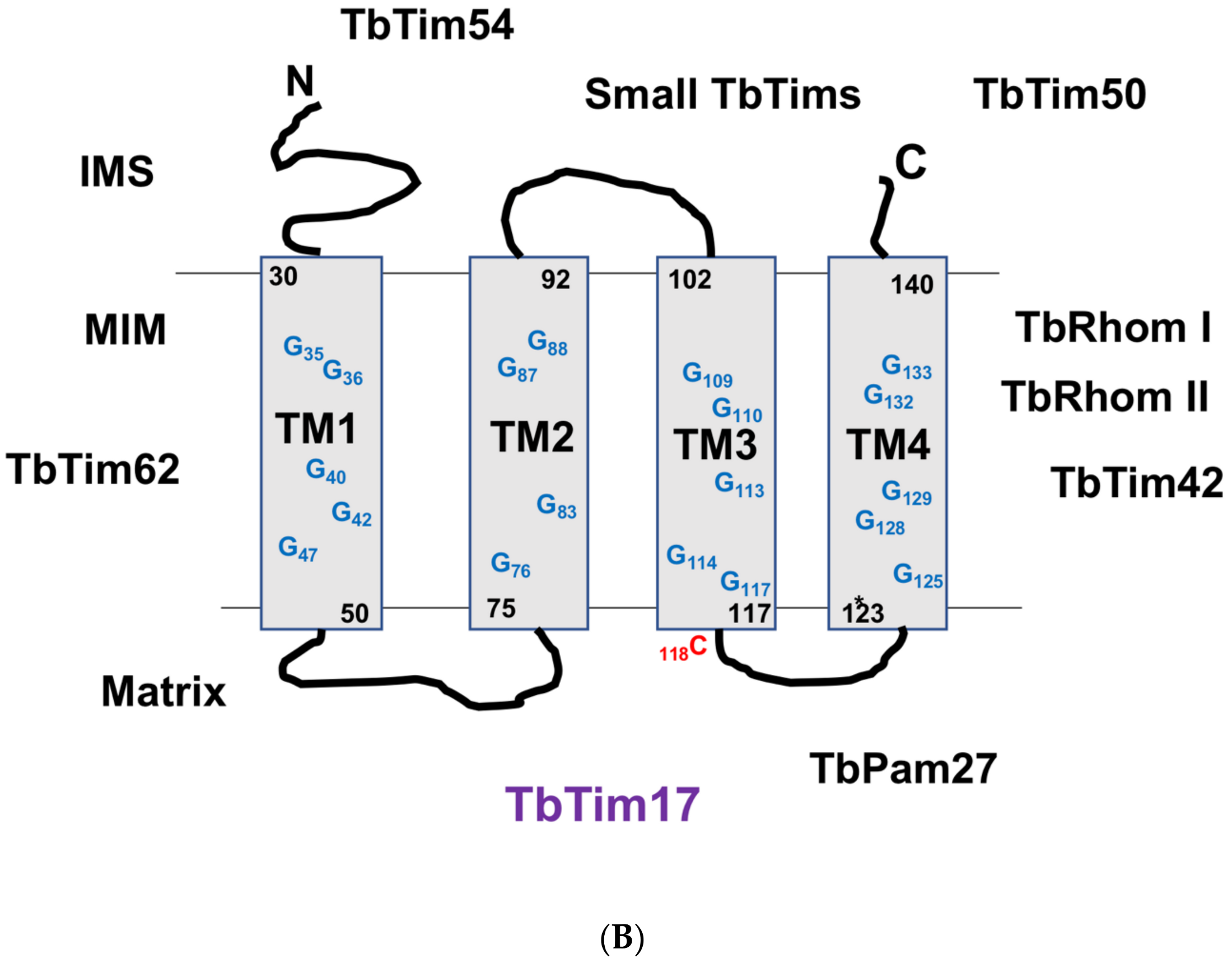
5. Tim17 Structure
6. Biogenesis of Tim17
7. Tim17 Interaction with Other Proteins and Its Function in Mitochondrial Protein Import
8. Non-Canonical Functions of Tim17
8.1. Role of Tim17 in Mitochondrial Stress Response
8.2. Connection of Tim17 Expression with Cancer
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mehta, M.M.; Weinberg, S.E.; Chandel, N.S. Mitochondrial control of immunity: Beyond ATP. Nat. Rev. Immunol. 2017, 17, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Eisner, V.; Picard, M.; Hajnóczky, G. Mitochondrial dynamics in adaptive and maladaptive cellular stress responses. Nat. Cell Biol. 2018, 20, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Rodriguez, M.; Bacskai, B.J. Mitochondria and calcium in Alzheimer’s disease: From cell signaling to neuronal cell death. Trends Neurosci. 2020. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Gincel, D. The voltage-dependent anion channel: Characterization, modulation, and role in cell life and death. Cell. Biochem. Biophys. 2003, 39, 279–292. [Google Scholar]
- Schleiff, E.; Becker, T. Common ground for protein translocation: Access control for mitochondria and chloroplasts. Nat. Rev. Mol. Cell Biol. 2011, 12, 48–59. [Google Scholar] [CrossRef]
- Senkler, J.; Senkler, M.; Braun, H.P. Structure and function of complex I in animals and plants—A comparative view. Physiol. Plant 2017, 161, 6–15. [Google Scholar] [CrossRef]
- Eramo, M.J.; Lisnyak, V.; Formosa, L.E.; Ryan, M.T. The ‘mitochondrial contact site and cristae organizing system’ (MICOS) in health and human disease. J. Biochem. 2020, 167, 243–255. [Google Scholar] [CrossRef]
- Truscott, K.N.; Brandner, K.; Pfanner, N. Mechanism of protein import into mitochondria. Curr. Biol. 2003, 13, R326–R337. [Google Scholar] [CrossRef] [Green Version]
- Hansen, K.G.; Herrmann, J.M. Transport of Proteins into mitochondria. Protein J. 2019, 38, 330–342. [Google Scholar] [CrossRef]
- Makiuchi, T.; Nozaki, T. Highly divergent mitochondrion-related organelles in anaerobic parasitic protozoa. Biochimie 2014, 100, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Neupert, W.; Herrmann, J.M. Translocation of proteins into mitochondria. Annu. Rev. Biochem. 2007, 76, 723–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, O.; Pfanner, N.; Meisinger, C. Mitochondrial protein import: From proteomics to functional mechanisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Drwesh, L.; Rapaport, D. Biogenesis pathways of a-helical mitochondrial outer membrane proteins. Biol. Chem. 2020, 401, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Fölsch, H.; Guiard, B.; Neupert, W.; Stuart, R.A. Internal targeting signal of the BCS1 protein: A novel mechanism of import into mitochondria. EMBO J. 1996, 15, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, V.; Singha, U.K.; Smith, J. T, Jr., Weems, E.; Chaudhuri, M. Trypanosome alternative oxidase possesses both an N-terminal and internal mitochondrial targeting signal. Eukaryot. Cell 2014, 13, 539–547. [Google Scholar]
- Bauer, M.F.; Gempel, K.; Reichert, A.S.; Rappold, G.A.; Lichtner, P.; Gerbitz, K.D.; Neupert, W.; Brunner, M.; Hofmann, S. Genetic and structural characterization of the human mitochondrial inner membrane translocase. J. Mol. Biol. 1999, 289, 69–82. [Google Scholar] [CrossRef]
- Lister, R.; Hulett, J.M.; Lithgow, T.; Whelan, J. Protein import into mitochondria: Origins and functions today (review). Mol. Membr. Biol. 2005, 22, 87–100. [Google Scholar] [CrossRef]
- Neupert, W.; Brunner, M. The protein import motor of mitochondria. Nat. Rev. Mol. Cell Biol. 2002, 3, 555–565. [Google Scholar] [CrossRef]
- Geissler, A.; Krimmer, T.; Bömer, U.; Guiard, B.; Rassow, J.; Pfanner, N. Membrane potential driven protein import into mitochondria. The sorting sequence of cytochrome b(2) modulates the dealtapsi-dependence of translocation of the matrix. Mol. Biol. Cell. 2000, 11, 3977–3991. [Google Scholar] [CrossRef]
- Glaser, S.M.; Miller, B.R.; Cumsky, M.G. Removal of a hydrophobic domain within the mature portion of a mitochondrial inner membrane protein causes its mislocalization to the matrix. Mol. Cell. Biol. 1990, 10, 1873–1881. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Baker, M.J.; Liem, M.; Louber, J.; McKenzie, M.; Atukorala, I.; Ang, C.S.; Keerthikumar, S.; Mathivanan, S.; Stojanovski, D. Tim29 is a novel subunit of the human TIM22 translocase and is involved in complex assembly and stability. Elife 2016, 5, e17463. [Google Scholar] [CrossRef] [PubMed]
- Kerscher, O.; Holder, J.; Srinivasan, M.; Leung, R.S.; Jensen, R.E. The Tim54p-Tim22p complex mediates insertion of proteins into the mitochondrial inner membrane. J. Cell Biol. 1997, 139, 1663–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, T.; Pfannschmidt, S.; Guiard, B.; Stojanovski, D.; Milenkovic, D.; Kutik, S.; Pfanner, N.; Meisinger, C.; Wiedemann, N. Biogenesis of the mitochondrial TOM complex: Mim1 promotes insertion and assembly of signal-anchored receptors. J. Biol. Chem. 2008, 283, 120–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, T.; Wagner, R. Mitochondrial Outer Membrane Channels: Emerging Diversity in Transport Processes. Bioessays 2018, 40, e1800013. [Google Scholar] [CrossRef]
- Dekker, P.J.T.; Ryan, M.T.; Brix, J.; Müller, H.; Hönlinger, A.; Pfanner, N. Preprotein translocase of the outer mitochondrial membrane: Molecular dissection and assembly of the general import pore complex. Mol. Cell. Biol. 1998, 18, 6515–6524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Künkele, K.-P.; Heins, S.; Dembowski, M.; Nargang, F.E.; Benz, R.; Thieffry, M.; Walz, J.; Lill, R.; Nussberger, S.; Neupert, W. The preprotein translocation channel of the outer membrane of mitochondria. Cell 1998, 93, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Araiso, Y.; Tsutsumi, A.; Qiu, J.; Imai, K.; Shiota, T.; Song, J.; Lindau, C.; Wenz, L.S.; Sakaue, H.; Yunoki, K.; et al. Structure of the mitochondrial import gate reveals. Distinct preprotein paths. Nature 2019, 575, 395–401. [Google Scholar] [CrossRef]
- Suzuki, H.; Okazawa, Y.; Komiya, T.; Saeki, K.; Mekada, E.; Kitada, S.; Ito, A.; Mihara, K. Characterization of rat TOM40, a central component of the preprotein translocase of the mitochondrial outer membrane. J. Biol. Chem. 2000, 275, 37930–37936. [Google Scholar] [CrossRef] [Green Version]
- Ghifari, A.S.; Gill-Hille, M.; Murcha, M.W. Plant mitochondrial protein import: The ins and outs. Biochem. J. 2018, 475, 2191–2208. [Google Scholar] [CrossRef]
- Panigrahi, R.; Whelan, J.; Vrielink, A. Exploring ligand recognition, selectivity and dynamics of TPR domains of chloroplast Toc64 and mitochondrial Om64 from Arabidopsis thaliana. J. Mol. Recognit. 2014, 27, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Kozjak, V. Wiedemann, N.; Milenkovic, D.; Lohaus, C.; Meyer, H.E.; Guiard, B.; Meisinger, C.; Pfanner, N. An essential role of Sam50 in the protein sorting and assembly machinery of the mitochondrial outer membrane. J. Biol. Chem. 2003, 278, 48520–48523. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, P.; Dagley, M.J.; Kono, M.; Wolynec, P.; Likić, V.A.; Foo, J.H.; Sedinová, M.; Tachezy, J.; Bachmann, A.; Bruchhaus, I.; et al. The essentials of protein import in the degggerate mitochondrion of Entamoeba histolytica. PLoS Pathog. 2010, 6, e1000812. [Google Scholar] [CrossRef] [Green Version]
- Dagley, M.J.; Dolezal, P.; Likic, V.A.; Smid, O.; Purcell, A.W.; Buchanan, S.K.; Tachezy, J.; Lithgow, T. The protein import channel in the outer mitosomal membrane of Giardia intestinalis. Mol. Biol. Evol. 2009, 26, 1941–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rada, P.; Doležal, P.; Jedelský, P.L.; Bursac, D.; Perry, A.J.; Šedinová, M.; Smíšková, K.; Novotný, M.; Beltrán, N.C.; Hrdý, I.; et al. The core components of organelle biogenesis and membrane transport in the hydrogenosomes of Trichomonas vaginalis. PLoS ONE 2011, 6, e24428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makki, A.; Rada, P.; Zarsky, V.; Kereiche, S.; Kovacik, L.; Novotny, M.; Jores, T.; Rapaport, D.; Tachezy, J. Triplet-pore structure of a highly divergent TOM complex of hydrogenosomes in Trichomonas vaginalis. PLoS Biol. 2001, 17, e3000098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomer, U.; Rassow, J.; Zufall, N.; Pfanner, N.; Meijer, M.; Maarse, A.C. The preprotein translocase of the inner mitochondrial membrane: Evolutionary conservation of targeting and assembly of Tim17. J. Mol. Biol. 1996, 262, 389–395. [Google Scholar] [CrossRef]
- Herrmann, J.M.; Neupert, W. Protein insertion into the inner membrane of mitochondria. IUBMB Life 2003, 55, 219–225. [Google Scholar] [CrossRef]
- Schendzielorz, A.B.; Bragoszewski, P.; Naumenko, N.; Gomkale, R.; Schulz, C.; Guiard, B.; Chacinska, A.; Rehling, P. Motor recruitment to the TIM23 channel’s lateral gate restricts polypeptide release into the inner membrane. Nat. Commun. 2018, 9, 4028. [Google Scholar] [CrossRef] [Green Version]
- Sirrenberg, C.; Bauer, M.F.; Guiard, B.; Neupert, W.; Brunner, M. Import of carrier proteins into the mitochondria inner membrane mediated by Tim22. Nature 1996, 384, 582–585. [Google Scholar] [CrossRef]
- Koehler, C.M.; Jarosch, E.; Tokatlidis, K.; Schmid, K.; Schweyen, R.J.; Schatz, G. Import of mitochondrial carriers mediated by essential proteins of the intermembrane space. Science 1998, 279, 369–373. [Google Scholar] [CrossRef]
- Kolli, R.; Soll, J.; Carrie, C. Plant Mitochondrial Inner Membrane Protein Insertion. Int. J. Mol. Sci. 2018, 19, 641. [Google Scholar] [CrossRef] [Green Version]
- Rassow, J.; Dekker, P.J.; van Wilpe, S.; Meijer, M.; Soll, J. The preprotein translocase of the mitochondrial inner membrane: Function and evolution. J. Mol. Biol. 1999, 286, 105–120. [Google Scholar] [CrossRef]
- Truscott, K.N.; Kovermann, P.; Geissler, A.; Merlin, A.; Meijer, M.; Driessen, A.J.; Rassow, J.; Pfanner, N.; Wagner, R. A presequence- and voltage-sensitive channel of the mitochondrial preprotein translocase formed by Tim23. Nat. Struct. Biol. 2001, 8, 1074–1082. [Google Scholar] [CrossRef] [Green Version]
- Rehling, P.; Model, K.; Brandner, K.; Kovermann, P.; Sickmann, A.; Meyer, H.E.; Kuhlbrandt, W.; Wagner, R.; Truscott, K.N.; Pfanner, N. Protein insertion into the mitochondrial inner membrane by a twin-pore translocase. Science 2003, 299, 1747–1751. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Caballero, S.; Grigoriev, S.M.; Herrmann, J.M. Tim17p regulates the twin pore structure and voltage gating of the mitochondrial protein import complex TIM23. J. Biol. Chem. 2007, 282, 3584–3593. [Google Scholar] [CrossRef]
- Mokranjac, D.; Paschen, S.A.; Kozany, C.; Prokisch, H.; Hoppins, S.C.; Nargang, F.E.; Neupert, W.; Hell, K. Tim50, a novel component of the TIM23 preprotein translocase of mitochondria. EMBO J. 2003, 22, 816–825. [Google Scholar] [CrossRef]
- Tamura, Y.; Harada, Y.; Shiota, T.; Yamano, K.; Watanabe, K.; Yokota, M.; Yamamoto, H.; Sesaki, H.; Endo, T. Tim23-Tim50 pair coordinates functions of translocators and motor proteins in mitochondrial protein import. J. Cell Biol. 2009, 184, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Matta, S.K.; Kumar, A.; D’Silva, P. Mgr2 regulates mitochondrial preprtein import by associating with channel-forming Tim23 subunit. Mol. Biol. Cell. 2020, 31, 1112–1123. [Google Scholar] [CrossRef]
- Callegari, S.; Richter, F.; Chojnacka, K.; Jans, D.C.; Lorenzi, I.; Pacheu-Grau, D.; Jakobs, S.; Lenz, C.; Urlaub, H.; Dudek, J.; et al. TIM29 is a subunit of the human carrier translocase required for protein transport. FEBS Lett. 2016, 590, 4147–4158. [Google Scholar] [CrossRef]
- Kerscher, O.; Sepuri, N.B.; Jensen, R.E. Tim18p is a new component of the Tim54p-Tim22p translocon in the mitochondrial inner membrane. Mol. Biol. Cell 2000, 11, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Mårtensson, C.U.; Becker, T. Mitochondrial protein transport meets lipid biosynthesis. Trends. Cell Biol. 2017, 27, 700–702. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Wang, Q.; Guan, Z.; Wu, Y.; Shen, C.; Hong, S.; Cao, J.; Zhang, X.; Yan, C.; Yin, P. Cryo-EM structure of the human mitochondrial translocase TIM22 complex. Cell Res. 2020. [Google Scholar] [CrossRef]
- Zhang, Y.; Ou, X.; Wang, X.; Sun, D.; Zhou, X.; Wu, X.; Li, Q.; Li, L. Structure of the mitochondrial TIM22 complex from yeast. Cell Res. 2020. [Google Scholar] [CrossRef]
- Bömer, U.; Meijer, M.; Maarse, A.C.; Hönlinger, A.; Dekker, P.J.; Pfanner, N.; Rassow, J. Multiple interactions of components mediating preprotein translocation across the inner mitochondrial membrane. EMBO J. 1997, 16, 2205–2216. [Google Scholar] [CrossRef] [Green Version]
- Ting, S.Y.; Schilke, B.A.; Hayashi, M. Architecture of the TIM23 inner mitochondrial translocon and interaction with the matrix import motor. J. Biol Chem. 2014, 289, 28689–28696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilke, B.A.; Hayashi, M.; Craig, E.A. Genetic analysis. Of complex interactions among. Components of the mitochondrial import motor and translocon in Saccharomyces cerevisiae. Genetics 2012, 190, 1341–1353. [Google Scholar] [CrossRef] [Green Version]
- Sinha, D.; Srivastava, S.; D’Silva, P. Functional diversity of human mitochondrial J-protein is independent of their association with the inner membrane presequence translocase. J. Biol. Chem. 2016, 291, 17345–17359. [Google Scholar] [CrossRef] [Green Version]
- Wiedemann, N.; Pfanner, N.; Chacinska, A. Chaperoning through the mitochondrial intermembrane space. Mol. Cell 2006, 21, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.J.; Webb, C.T.; Stroud, D.A.; Palmer, C.S.; Frazier, A.E.; Guiard, B.; Chacinska, A.; Gulbis, J.M.; Ryan, M.T. Structural and functional requirements for activity of the Tim9-Tim10. Mol. Biol. Cell 2009, 20, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Weinhäupl, K.; Lindau, C.; Hessel, A.; Wang, Y.; Schütze, C.; Jores, T.; Melchionda, L.; Schönfisch, B.; Kalbacher, H.; Bersch, B.; et al. Structural basis of membrane protein chaoeroning through the mitochondrial intermembrane space. Cell 2018, 175, 1365–1378.e25. [Google Scholar] [CrossRef] [Green Version]
- Gebert, N.; Chacinska, A.; Wagner, K.; Guiard, B.; Koehler, C.M.; Rehling, P.; Pfanner, N.; Wiedemann, N. Assembly of the three small Tim proteins precedes docking to the mitochondrial carrier translocase. EMBO Rep. 2008, 9, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Hoppins, S.C.; Nargang, F.E. The Tim8–13 complex of Neurospora crassa functions in the assembly of proteins into both mitochondrial membranes. J. Biol. Chem. 2004, 279, 11245–12396. [Google Scholar] [CrossRef] [Green Version]
- Ceh-Pavia, E.; Spiller, M.P.; Lu, H. Folding and biogenesis of mitochondrial small Tim proteins. Int. J. Mol Sci. 2013, 14, 16685–16705. [Google Scholar] [CrossRef] [Green Version]
- Mühlenbein, N.; Hofmann, S.; Rothbauer, U.; Bauer, M.F. Organization and function of the small Tim complexes acting along the import pathway of metabolite carriers into mammalian mitochondria. J. Biol. Chem. 2004, 279, 13540–13546. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A. Mitochondrial protein import in trypanosomatids: Variations on a theme or fundamentally different? PLoS Pathog. 2018, 14, e1007351. [Google Scholar] [CrossRef]
- Eckers, E.; Cyrklaff, M.; Simpson, L.; Deponte, M. Mitochondrial protein import pathways are functionally conserved among eukaryotes despite compositional diversity of the import machineries. Biol. Chem. 2012, 393, 513–524. [Google Scholar] [CrossRef]
- Pusnik, M.; Schmidt, O.; Perry, A.J.; Oeljeklaus, S.; Niemann, M.; Warscheid, B.; Lithgow, T.; Meisinger, C.; Schneider, A. Mitochondrial preprotein translocase of trypanosomatids has a bacterial origin. Curr. Biol. 2011, 21, 1738–1743. [Google Scholar] [CrossRef] [Green Version]
- Harsman, A.; Niemann, M.; Pusnik, M.; Schmidt, O.; Burmann, B.M.; Hiller, S.; Meisinger, C.; Schneider, A.; Wagner, R. Bacterial origin of a mitochondrial outer membrane protein translocase: New paerspectives from comparative single channel electrophysiology. J. Biol. Chem. 2012, 287, 31437–31445. [Google Scholar] [CrossRef] [Green Version]
- Mani, J.; Meisinger, C.; Schneider, A. Peeping at TOMs-diverse entry gates to mitochondria provide insights into the evolution of Eukaryotes. Mol. Biol. Evol. 2016, 33, 3337–3351. [Google Scholar] [CrossRef] [Green Version]
- Mani, J.; Rout, S.; Desy, S.; Schneider, A. Mitochondrial protein import – Functional analysis of the highly diverged Tom22 orthologue of Trypanosoma brucei. Sci. Rep. 2017, 7, 40738. [Google Scholar] [CrossRef] [Green Version]
- Singha, U.K.; Peprah, E.; Williams, S.; Walker, R.; Saha, L.; Chaudhuri, M. Characterization of the mitochondrial inner membrane protein translocator Tim17 from Trypanosoma brucei. Mol. Biochem. Parasitol. 2008, 159, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singha, U.K.; Hamilton, V.; Duncan, M.R.; Weems, E.; Tripathi, M.K.; Chaudhuri, M. Protein translocase of mitochondrial inner membrane in Trypanosoma brucei. J. Biol. Chem. 2012, 287, 14480–14493. [Google Scholar] [CrossRef] [Green Version]
- Weems, E.; Singha, U.K.; Hamilton, V.; Smith, J.T.; Waegemann, K.; Mokranjac, D.; Chaudhuri, M. Functional complementation analyses reveal that the single PRAT family protein of Trypanosoma brucei is a divergent homolog of Tim17 in Saccharomyces cerevisiae. Eukaryot. Cell 2015, 14, 286–296. [Google Scholar] [CrossRef] [Green Version]
- Duncan, M.R.; Fullerton, M.; Chaudhuri, M. Tim50 in Trypanosoma brucei possesses a dual specificity phosphatase activity and is critical for mitochondrial protein import. J. Biol. Chem. 2013, 288, 3184–3197. [Google Scholar] [CrossRef] [Green Version]
- Harsman, A.; Oeljeklaus, S.; Wenger, C.; Huot, J.L.; Warscheid, B.; Schneider, A. The non-canonical mitochondrial inner membrane presequence translocase of trypanosomatids contains two essential rhomboid-like proteins. Nat. Commun. 2016, 7, 13707. [Google Scholar] [CrossRef] [Green Version]
- Týč, J.; Klingbeil, M.M.; Lukeš, J. Mitochondrial heat shock protein machinery hsp70/hsp40 is indispensable for proper mitochondrial DNA maintenance and replication. mBio. 2015, 6, e02425-14. [Google Scholar]
- Mensa-Wilmot, K.; Hoffman, B.; Wiedeman, J.; Sullenberger, C.; Sharma, A. Kinetoplast division. Factors in a trypanosome. Trends Parasitol. 2019, 35, 119–128. [Google Scholar] [CrossRef]
- Von Käne, L.C.; Muñoz-Gómez, S.A.; Oeljeklaus, S.; Wenger, C.; Warscheid, B.; Wideman, J.G.; Harsman, A.; Schneider, A. Homologue replacement in the import motor of the mitochondrial inner membrane of trypanosomes. Elife 2020, 9, e52560. [Google Scholar]
- Singha, U.K.; Hamilton, V.; Chaudhuri, M. Tim62, a Novel Mitochondrial Protein in Trypanosoma brucei, Is Essential for Assembly and Stability of the TbTim17 Protein Complex. J. Biol. Chem. 2015, 290, 23226–23239. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A. Mitochondrial tRNA import and its consequences for mitochondrial translation. Annu. Rev. Biochem. 2011, 80, 1033–1053. [Google Scholar] [CrossRef] [Green Version]
- Niemann, M.; Harsman, A.; Mani, J.; Peikert, C.D.; Oeljeklaus, S.; Warscheid, B.; Wagner, R.; Schneider, A. tRNAs and proteins use the same import channel for translocation across the mitochondrial outer membrane of trypanosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E7679–E7687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barozai, M.Y.K.; Chaudhuri, M. Role of the translocase of the mitochondrial inner membrane in the import of tRNAs into mitochondria in Trypanosoma brucei. Gene 2020, 748, 144705. [Google Scholar] [CrossRef] [PubMed]
- Zikova, A.; Verner, Z.; Nenarokova, A.; Michels, P.A.M.; Lukes, J. A paradigm shift: The mitoproteomes of procyclic and bloodstream Trypanosoma brucei are comparably complex. PLoS Pathog. 2017, 13, e1006679. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, M.; Nargang, F.E. Import and assembly of Neurospora crassa Tom40 into mitochondria of Trypanosoma brucei in vivo. Curr. Genet. 2003, 44, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Tschopp, F.; Charrière, F.; Schneider, A. In vivo study in Trypanosoma brucei links mitochondrial transfer RNA import to mitochondrial protein import. EMBO Rep. 2011, 12, 825–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žárský, V.; Doležal, P. Evolution of the Tim17 protein family. Biol. Direct. 2016, 11, 54. [Google Scholar]
- Rossig, C.; Reinbothe, C.; Gray, J.; Valdes, O.; von Wettstein, D.; Reinbothe, S. Three proteins mediate import of transit sequence-less precursors into the inner envelope of chloroplasts in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2013, 110, 19962–19967. [Google Scholar] [CrossRef] [Green Version]
- Reguenga, C.; Oliveira, M.E.; Gouveia, A.M.; Eckerskorn, C.; Sá-Miranda, C.; Azevedo, J.E. Identification of a 24 kDa intrinsic membrane protein from mammalian peroxisomes. Biochim. Biophys. Acta 1999, 1445, 337–341. [Google Scholar] [CrossRef]
- Guarani, V.; Paulo, J.; Zhai, B.; Huttlin, E.L.; Gygi, S.P.; Harper, J.W. TIMMDC1/C3orf1 functions as a membrane-embedded mitochondrial complex I assembly factor through association with the MCIA complex. Mol. Cell. Biol. 2014, 34, 847–861. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Carrie, C.; Giraud, E.; Elhafez, D.; Narsai, R.; Duncan, O.; Whelan, J.; Murcha, M.W. location of the mitochondrial preprotein transporters B14.7 and Tim23–2 in complex I and the TIM17:23 complex in Arabidopsis links mitochondrial activity and biogenesis. Plant Cell 2012, 24, 2675–2695. [Google Scholar] [CrossRef] [Green Version]
- Gentle, I.E.; Perry, A.J.; Alcock, F.H.; Likić, V.A.; Dolezal, P.; Ng, E.T.; Purcell, A.W.; McConnville, M.; Naderer, T.; Chanez, A.-L.; et al. Conserved motifs reveal details of ancestry and structure in the small TIM chaperones of the mitochondrial intermembrane space. Mol. Biol. Evol. 2007, 24, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Pyrihova, E.; Motyckova, A.; Voleman, L.; Wandyszewska, N.; Fiser, R.; Seydlova, G.; Roger, A.; Kolisko, M.; Dolezal, P. A Single Tim Translocase in the Mitosomes of Giardia intestinalis Illustrates Convergence of Protein Import Machines in Anaerobic Eukaryotes. Genome Biol. Evol. 2018, 10, 2813–2822. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, V.L.; Heinz, E.; Shingu-Vazquez, M.; Qu, Y.; Jelicic, B.; Lo, T.L.; Beilharz, T.H.; Dumsda, Y.G.; Gabriel, K.; Traven, A.; et al. A model system for mitochondrial biogenesis reveals evolutionary rewiring of protein import and membrane assembly pathways. Proc. Natl. Acad. Sci. USA 2012, 109, E3356–E3366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, D.; Srivastava, S.; Krishna, L.; D’Silva, P. Unraveling the intricate organization of mammalian mitochondrial presequence translocase: Existence of multiple translocases for maintenance of mitochondrial function. Mol. Cell Biol. 2014, 34, 1757–1775. [Google Scholar] [CrossRef] [Green Version]
- Murcha, M.W.; Lister, R.; Ho, A.Y.; Whelan, J. Identification, expression, and import of components 17 and 23 of the inner mitochondrial membrane translocase from Arabidopsis. Plant Physiol. 2003, 131, 1737–1747. [Google Scholar] [CrossRef] [Green Version]
- Murcha, M.W.; Elhafez, D.; Lister, R.; Tonti-Filippini, J.; Baumgartner, M.; Philippar, K.; Carrie, C.; Mokranjac, D.; Soll, J.; Whelan, J. Characterization of the preprotein and amino acid transporter gene family in Arabidopsis. Plant Physiol. 2007, 143, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Heinz, E.; Lithgow, T. Back to basics: A revealing secondary reduction of the mitochondrial protein import pathway in diverse intracellular parasites. Biochim. Biophys. Acta 2013, 1833, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Maarse, A.C.; Blom, J.; Keil, P.; Pfanner, N.; Meijer, M. Identification of the essential yeast protein MIM17, an integral mitochondrial inner membrane protein involved in protein import. FEBS Lett. 1994, 349, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Kübrich, M.; Keil, P.; Rassow, J.; Dekker, P.J.; Blom, J.; Meijer, M.; Pfanner, N. The polytopic mitochondrial inner membrane protein MIM17 and MIM23 operates at the same preprotein import site. FEBS Lett. 1994, 349, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Berthold, J.; Bauer, M.F.; Schneider, H.C.; Klaus, C.; Dietmeier, K.; Neupert, W.; Brunner, M. The MIM complex mediates preprotein translocation across the mitochondrial inner membrane and couples it to the mt-Hsp70/ATP driving system. Cell 1995, 81, 1085–1093. [Google Scholar] [CrossRef] [Green Version]
- Dekker, P.J.; Martin, F.; Maarse, A.C.; Bömer, U.; Müller, H.; Guiard, B.; Meijer, M.; Rassow, J.; Pfanner, N. The Tim core complex defines the number of mitochondrial translocation contact sites and can hold arrested preproteins in the absence of matrix Hsp70-Tim44. EMBO J. 1997, 16, 5409–5419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Laan, M.; Hutu, D.P.; Rehling, P. On the mechanism of preprotein import by the mitochondrial presequence translocase. Biochim. Biophys. Acta 2010, 1803, 732–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacinska, A.; Lind, M.; Frazier, A.E.; Dudek, J.; Meisinger, C.; Geissler, A.; Sickmann, A.; Meyer, H.E.; Truscott, K.N.; Guiard, B.; et al. Mitochondrial presequence translocase: Switching between TOM tethering and motor recruitement involves Tim21 and Tim17. Cell 2005, 120, 817–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singha, U.K.; Tripathi, A.; Smith, J.T.; Quinones, L.; Saha, A.; Singha, T.; Chaudhuri, M. Tim54, a novel IM-associated protein, is involved in the import of the internal signal-containing mitochondrial protein in Trypanosoma brucei. Biol. Cell 2020. [Google Scholar] [CrossRef] [PubMed]
- Wenger, C.; Oeljeklaus, S.; Warscheid, B.; Schneider, A.; Harsman, A. A trypanosomal orthologue of an intermembrane space chaperone has a non-canonical function in biogenesis of the single mitochondrial inner membrane protein translocase. PLoS Pathog. 2017, 13, e1006550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.T., Jr.; Singha, U.K.; Misra, S.; Chaudhuri, M. Divergent Small Tim Homologues Are Associated with TbTim17 and Critical for the Biogenesis of TbTim17 Protein Complexes in Trypanosoma brucei. mSphere 2018, 3, e00204. [Google Scholar] [CrossRef] [Green Version]
- Donzeau, M.; Káldi, K.; Adam, A.; Paschen, S.; Wanner, G.; Guiard, B.; Bauer, M.F.; Neupert, W.; Brunner, M. links the inner and outer mitochondrial membrane. Cell 2000, 101, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.F.; Sirrenberg, C.; Neupert, W.; Brunner, M. Role of Tim23 as voltage sensor and presequence receptor in protein import into mitochondria. Cell 1996, 87, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Geissler, A.; Chacinska, A.; Truscott, K.N.; Wiedemann, N.; Brandner, K.; Sickmann, A.; Meyer, H.E.; Meisinger, C.; Pfanner, N.; Rehling, P. The mitochondrial presequence translocase: An essential role of Tim50 in directing preproteins to the import channel. Cell 2002, 111, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Waegemann, K.; Popov-Čeleketić, D.; Neupert, W.; Azem, A.; Mokranjac, D. Cooperation of TOM and TIM23 complexes during translocation of proteins into mitochondria. J. Mol. Biol. 2015, 427, 1075–1084. [Google Scholar] [CrossRef]
- Popov-Celeketić, D.; Mapa, K.; Neupert, W.; Mokranjac, D. Active remodeling of the TIM23 complex during translocation of preproteins into mitochondria. EMBO J. 2008, 27, 1469–1480. [Google Scholar] [PubMed]
- Günsel, U.; Paz, E.; Gupta, R.; Mathes, I.; Azem, A.; Mokranjac, D. In vivo dissection of the Intrinsically disordered receptor domain of Tim23. J. Mol. Biol. 2020, 432, 3326–3337. [Google Scholar] [CrossRef] [PubMed]
- Káldi, K.; Bauer, M.F.; Sirrenberg, C.; Neupert, W.; Brunner, M. Biogenesis of Tim23 and Tim17, integral components of the TIM machinery for matrix-targeted preproteins. EMBO J. 1998, 17, 1569–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothbauer, U.; Hofmann, S.; Mühlenbein, N.; Paschen, S.A.; Gerbitz, K.D.; Neupert, W.; Brunner, M.; Bauer, M.F. Role of the deafness dystonia peptide 1 (DDP1) in import of humanTim23 into the inner membrane of mitochondria. J. Biol. Chem. 2001, 2766, 37327–37334. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.J.; Alder, N.N.; Jensen, R.E.; Johnson, A.E. The Tim9p/Tim10p and Tim8p/Tim13p complexes bind to specific sites on Tim23 during mitochondrial protein import. Mol. Biol. Cell 2007, 18, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Beverly, K.N.; Sawaya, M.R.; Schmid, E.; Koehler, C.M. The Tim8-Tim13 complex has multiple substrate binding sites and binds cooperatively to Tim23. J. Mol. Biol. 2008, 382, 1144–1156. [Google Scholar] [CrossRef] [Green Version]
- Meier, S.; Neupert, W.; Herrmann, J.M. Conserved N-terminal negative charges in the Tim17 subunit of the TIM23 translocase play a critical role in the import of preproteins into mitochondria. J. Biol. Chem. 2005, 280, 7777–7785. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, A.; Peleh, V.; Martinez-Caballero, S.; Wollweber, F.; Sommer, F.; van der Laan, M.; Schroda, M.; Alexander, R.T.; Campo, M.L.; Herrmann, J.M. A disulfide bond in the TIM23 complex is crucial for voltage gating and mitochondrial protein import. J. Cell Biol. 2016, 214, 417–431. [Google Scholar] [CrossRef] [Green Version]
- Wrobel, L.; Sokol, A.M.; Chojnacka, M.; Chacinska, A. The presence of disulfide bonds reveals an evolutionary conserved mechanism involved in mitochondrial protein translocase assembly. Sci. Rep. 2016, 6, 27484. [Google Scholar] [CrossRef] [Green Version]
- Weems, E.; Singha, U.K.; Smith, J.T.; Chaudhuri, M. The divergent N-terminal domain of Tim17 is critical for its assembly in the TIM complex in Trypanosoma brucei. Mol. Biochem. Parasitol. 2017, 218, 4–15. [Google Scholar] [CrossRef]
- Matta, S.K.; Pareek, G.; Bankapalli, K.; Oblesha, A.; D’Silva, P. Role of Tim17 transmembrane regions in regulating the Architecture of presequence translocase and mitochondrial DNA stability. Mol. Cell. Biol. 2017, 37, e00491-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demishtein-Zohary, K.; Günsel, U.; Marom, M.; Banerjee, R.; Neupert, W.; Azem, A.; Mokranjac, D. Role of Tim17 in coupling the import motor to the translocation channel. Of the mitochondrial presequence translocase. Elife 2017, 6, e22696. [Google Scholar] [CrossRef] [PubMed]
- Murcha, M.W.; Wang, Y.; Whelan, J. A molecular link between mitochondrial preprotein transporters and respiratory chain complex. Plant Signal Behav. 2012, 7, 1594–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokranjac, D.; Popov-Celeketić, D.; Hell, K.; Neupert, W. Role of Tim21 in mitochondrial translocation contact sites. J. Biol. Chem. 2005, 280, 23437–23440. [Google Scholar] [CrossRef] [Green Version]
- van der Laan, M.; Wiedemann, N.; Mick, D.U.; Guiard, B.; Rehling, P.; Pfanner, N. A role for Tim21 in membrane-potential-dependent preporting sorting in mitochondria. Curr. Biol. 2006, 6, 2271–2276. [Google Scholar] [CrossRef]
- Rainbolt, T.K.; Atanassova, N.; Genereux, J.C.; Wiseman, R.L. Stress-regulated translational attenuation adapts mitochondrial protein import through Tim17A degradation. Cell Metab. 2013, 18, 908–919. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, O.; Harbauer, A.B.; Rao, S.; Eyrich, B.; Zahedi, R.P.; Stojanovski, D.; Schönfisch, B.; Guiard, B.; Sickmann, A.; Pfanner, N.; et al. Regulation of mitochondrial protein import by cytosolic kinases. Cell 2011, 144, 227–239. [Google Scholar] [CrossRef]
- Rao, S.; Schmidt, O.; Harbauer, A.B.; Schönfisch, B.; Guiard, B.; Pfanner, N.; Meisinger, C. Biogenesis of the preprotein translocase of the outer mitochondrial membrane: Protein kinase A phosphorylates the precursor Tom40 and impairs its import. Mol. Biol. Cell. 2012, 23, 118–1627. [Google Scholar] [CrossRef]
- Pellegrino, M.W.; Nargund, A.M.; Haynes, C.M. Signaling the mitochondrial unfolded protein response. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2013, 1833, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.C.; Van Aken, O. Mitochondrial unfolded protein-related response across kingdoms: Similar problems, different regulators. Mitochondrion 2020, 53, 166–177. [Google Scholar] [CrossRef]
- Nargund, A.M.; Pellegrino, M.W.; Fiorese, C.J.; Baker, B.M.; Haynes, C.M. Mitochondrial import efficiency of ATFS-1 regulates mitochondrial UPR activation. Science 2012, 337, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Nargund, A.M.; Fiorese, C.J.; Pellegrino, M.W.; Deng, P.; Haynes, C.M. Mitochondrial and nuclear accumulation of the transcription factor ATFS-1 promotes OXPHOS recovery during the UPR(mt). Mol. Cell. 2015, 58, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zöller, E.; Laborenz, J.; Krämer, L.; Boos, F.; Räschle, M.; Alexander, R.T.; Herrmann, J.M. The intermembrane space protein Mix23 is a novel stress-induced mitochondrial import factor. J. Biol. Chem. 2020, 295, 14686–14697. [Google Scholar] [CrossRef] [PubMed]
- Marada, A.; Allu, P.K.; Murari, A.; PullaReddy, B.; Tammineni, P.; Thiriveedi, V.R.; Danduprolu, J.; Sepuri, N.B. Mge1, a nucleotide exchange factor of Hsp70 acts as an oxidative sensor to regulate mitochondrial Hsp70 function. Mol. Biol. Cell 2013, 24, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Sinha, D.; Saha, P.P.; Marthala, H.; D’Silva, P. Maggmas functions as a ROS reglator and provides cytoprotection against oxidative stress-mediated damage. Cell Death Dis. 2014, 5, e1394. [Google Scholar] [CrossRef] [Green Version]
- Wojityla, L.; Kosmala, A.; Garnczarska, M. Lupine embryo axes under salinity stress. II. Mitochondrial proteome response. Acta Physiol. Plant 2013, 35, 2383–2392. [Google Scholar] [CrossRef] [Green Version]
- Fullerton, M.; Singha, U.K.; Duncan, M.; Chaudhuri, M. Down regulation of Tim50 in Trypanosoma brucei increases tolerance to oxidative stress. Mol. Biochem. Parasitol. 2015, 199, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Singh, U.K.; Paromov, V.; Hill, S.; Pratap, S.; Rose, K.; Chaudhuri, M. The crosstalk between TbTim50 and PIP39, two aspartate-based protein phosphatases, maintain cellular homeostasis in Trypanosoma brucei. mSphere 2019, 4, e00353–19. [Google Scholar]
- Avagliano, A.; Ruocco, M.R.; Aliotta, F.; Belviso, I.; Accurso, A.; Masone, S.; Montagnani, S.; Arcucci, A. Mitochondrial Flexibility of Breast Cancers: A growth advantage and a therapeutic opportunity. Cell 2019, 8, 401. [Google Scholar] [CrossRef] [Green Version]
- Fulda, S.; Galluzzi, L.; Kroemer, G. Targeting mitochondria for cancer therapy. Nat. Rev. Drug Discov. 2010, 9, 447–464. [Google Scholar] [CrossRef]
- Yang, X.; Si, Y.; Tao, T.; Martin, T.A.; Cheng, S.; Yu, H.; Li, J.; He, J.; Jiang, W.G. The Impact of TIMM17A on Aggressiveness of human breast cancer cells. Anticancer Res. 2016, 366, 1237–1241. [Google Scholar]
- Salhab, M.; Patani, N.; Jiang, W.; Mokbel, K. High TIMM17A expression is associated with adverse pathological and clinical outcomes in human breast cancer. Breast Cancer 2012, 19, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Deng, S.; Pang, X.; Song, Y.; Luo, S.; Jin, L.; Pan, Y. LncRNA NEAT1 silenced miR-133b promotes migration and invasion of breast cancer cells. Int. J. Mol. Sci. 2019, 20, 33616. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Type | Destination | Examples | Reference |
|---|---|---|---|
| N-terminal cleavable 1 | Matrix | Most of the matrix proteins 2 | [11,18] |
| Stop-transfer 3 | MIM, IMS | Cox5a, Mia40, Tim50, Yme1, Cytb2 | [18,19,20] |
| Internal 4 | MIM | Mitochondrial carrier proteins (SLC25) family proteins 5 | [21,22] |
| Signal/tail-anchored 6 | MOM | Tom70, Tom20, hFis1, Bak, Bax | [13,23] |
| Internal presequence type 7 | MIM | BCS1, TAO 8 | [14,15] |
| Complex | Yeast/Fungi | Human | Plant | Trypanosomatids |
|---|---|---|---|---|
| TIM23 | Tim23 Tim17 Tim50 Tim21 Mgr2 | TIMM23 TIMM17 TIMM50 | Tim23 Tim17 Tim50 Tim21 | |
| PAM | Pam18 Pam16 Pam17 Tim44 mHSP70 Mge-1 | DnaJC19 a DnaJC15 a Magma b Mortalin/HSPA9 c | Pam18 Pam16 Tim14 Tim44 Hsp70 Mge1 | Pam27 mHSP70 Mge1 Pam16 Pam18 |
| TIM22 | Tim22 Tim54 Tim18 Sdh3 Tim12 | TIMM22 TIMM29 AGK | Tim22 | |
| Small Tims | Tim9 Tim10 Tim8 Tim13 Tim12 | Tim9 Tim10a Tim10b Tim8 Tim13 | Tim9 Tim10 Tim8 Tim13 | Tim9 Tim10 Tim8/13 Tim11 Tim12 Tim13 |
| TbTIM d | Tim17 Tim62 Tim42 Tim50 Tim54 Rhomboid I Rhomboid II ACAD Small Tims |
| Species | Tim17 | Tim22 | Tim23 | References |
|---|---|---|---|---|
| Saccharomyces cerevisiae | 1 | 1 | 1 | [11,18,42] |
| Candida Albicans | 1 | 1 | 1 | [93] |
| Homo sapien | 2 | 1 | 2 | [16,94] |
| Drosophila melanogaster | 3 | >1 | >1 | [86] |
| Plants (Arabidopsis thaliana) | 3 | 1 | 2 | [95,96] |
| Euglenozoa | 1 | 1 | 1 | [86] |
| Trypanosoma cruzi | 1 | [72,91] | ||
| Trypanosoma brucei | 1 | [72,73] | ||
| Leishmania species | 1 | [91] | ||
| Plasmodium species | 1 | 1 | 1 | [93,97] |
| Giardia Intestinalis | 1 | [86,92] | ||
| Acanthameoba castellanii | 1 | 1 | 1 | [86] |
| Trichomonas vaginalis | 4 to 5 a | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhuri, M.; Darden, C.; Soto Gonzalez, F.; Singha, U.K.; Quinones, L.; Tripathi, A. Tim17 Updates: A Comprehensive Review of an Ancient Mitochondrial Protein Translocator. Biomolecules 2020, 10, 1643. https://doi.org/10.3390/biom10121643
Chaudhuri M, Darden C, Soto Gonzalez F, Singha UK, Quinones L, Tripathi A. Tim17 Updates: A Comprehensive Review of an Ancient Mitochondrial Protein Translocator. Biomolecules. 2020; 10(12):1643. https://doi.org/10.3390/biom10121643
Chicago/Turabian StyleChaudhuri, Minu, Chauncey Darden, Fidel Soto Gonzalez, Ujjal K. Singha, Linda Quinones, and Anuj Tripathi. 2020. "Tim17 Updates: A Comprehensive Review of an Ancient Mitochondrial Protein Translocator" Biomolecules 10, no. 12: 1643. https://doi.org/10.3390/biom10121643
APA StyleChaudhuri, M., Darden, C., Soto Gonzalez, F., Singha, U. K., Quinones, L., & Tripathi, A. (2020). Tim17 Updates: A Comprehensive Review of an Ancient Mitochondrial Protein Translocator. Biomolecules, 10(12), 1643. https://doi.org/10.3390/biom10121643