Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters


 ,
,  , and
, and
Abstract
:1. Introduction
2. Material and Methods
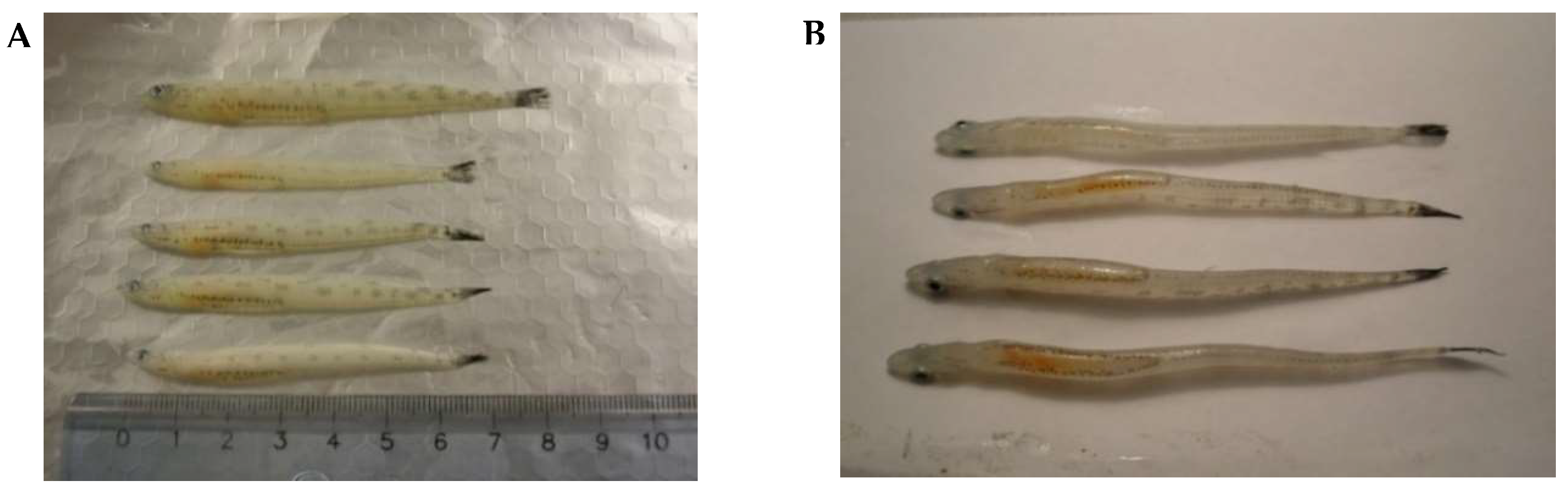
2.1. Sample Collection and Description
2.2. Lipid Extraction
2.3. Lipid Class Analysis
2.4. Fatty Acid Analysis
2.5. Statistical Analysis (Lipid Study)
2.6. Histological Analysis
2.7. Statistical Analysis (Histological Analysis)
3. Results
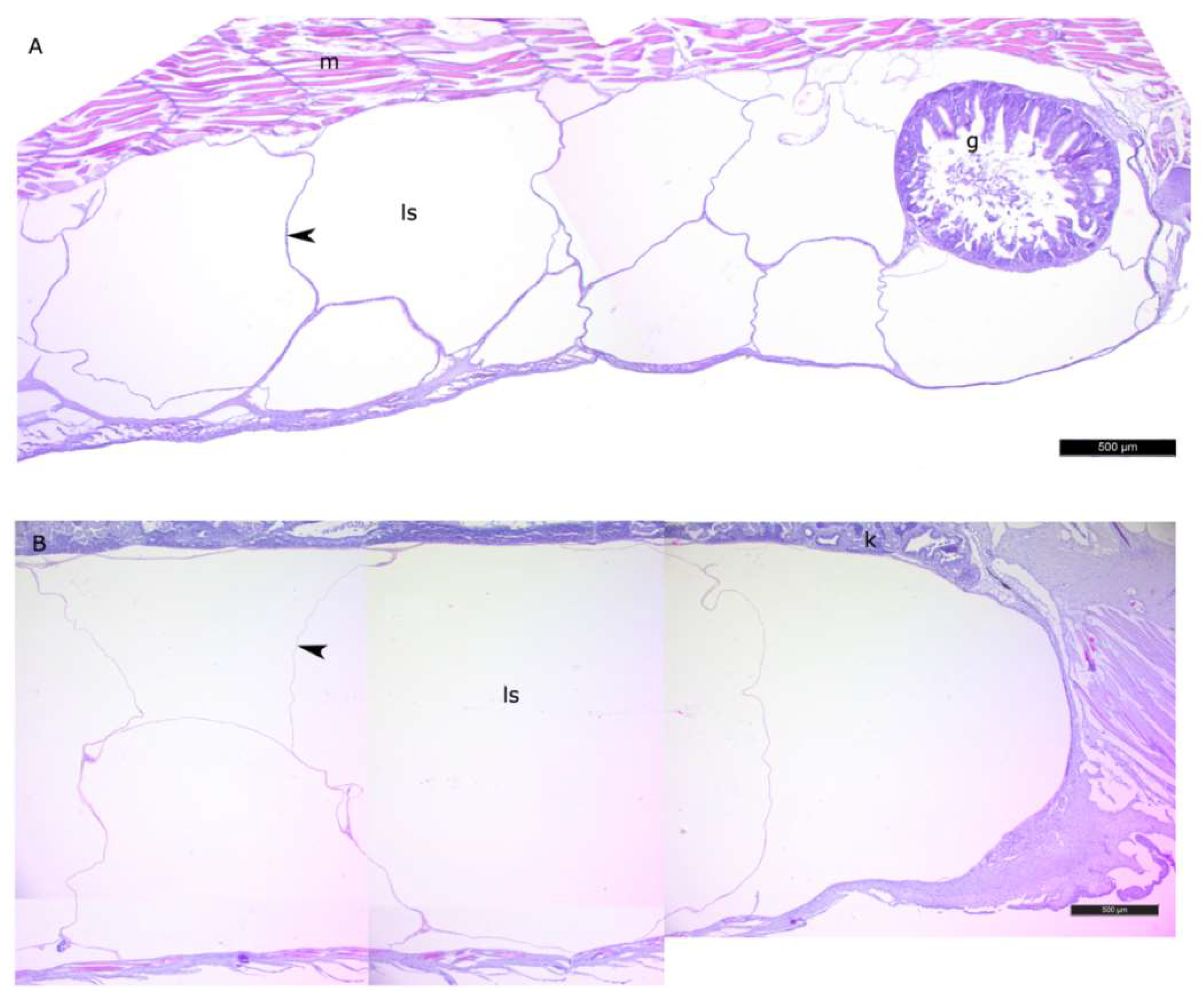
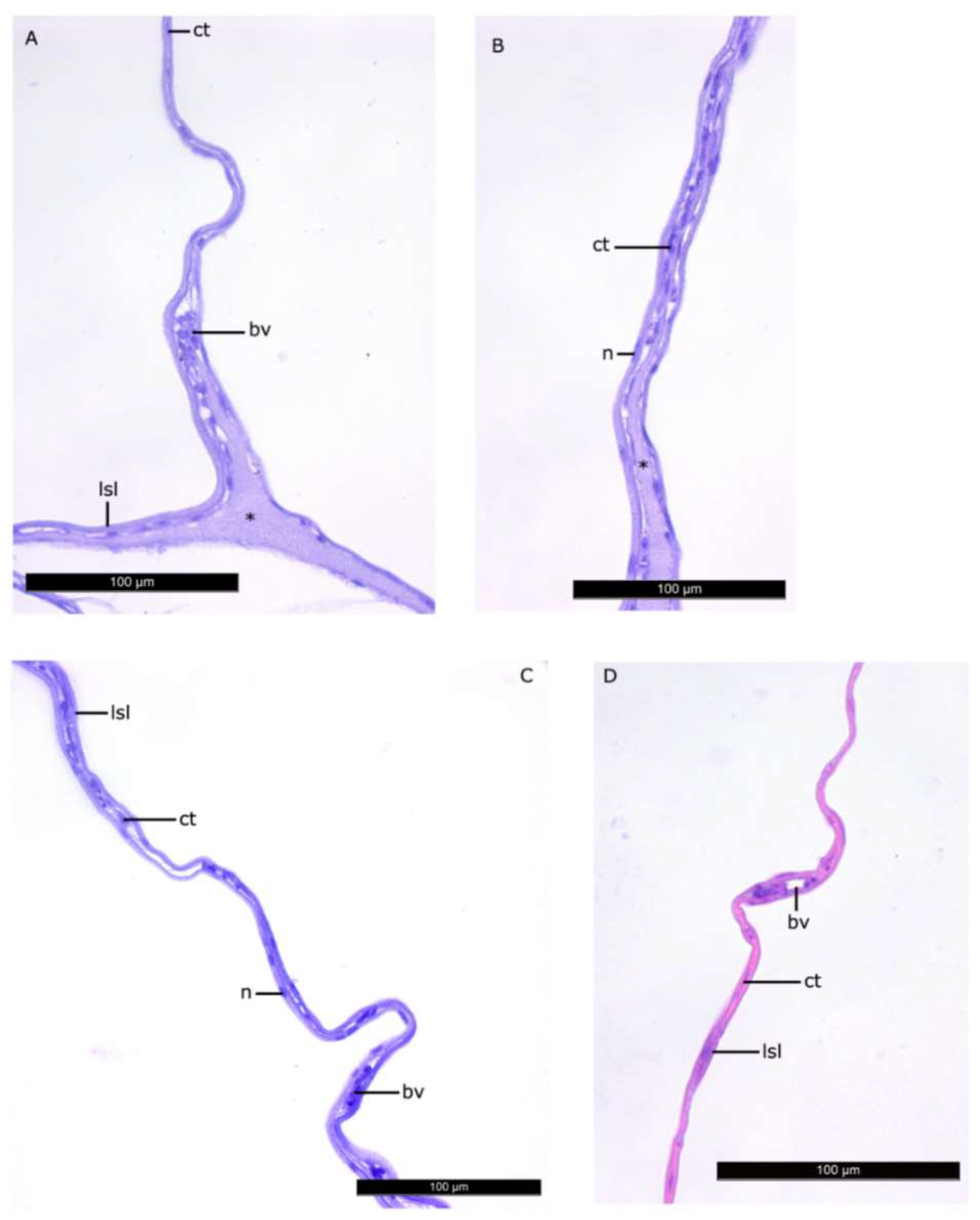
3.1. Histomorphology of the Lipid Sac of Postlarval L. Maculatus (L2–L5 Developmental Stages)
3.2. Seasonal Dynamics of Total Lipids in the Lipid Sac and Muscles of Pelagic Postlarval (L3 Developmental Stage) L. Maculatus
3.3. Seasonal Dynamics of Certain Lipid Classes in Lipid Sac and Muscles of Pelagic Postlarval L. Maculatus
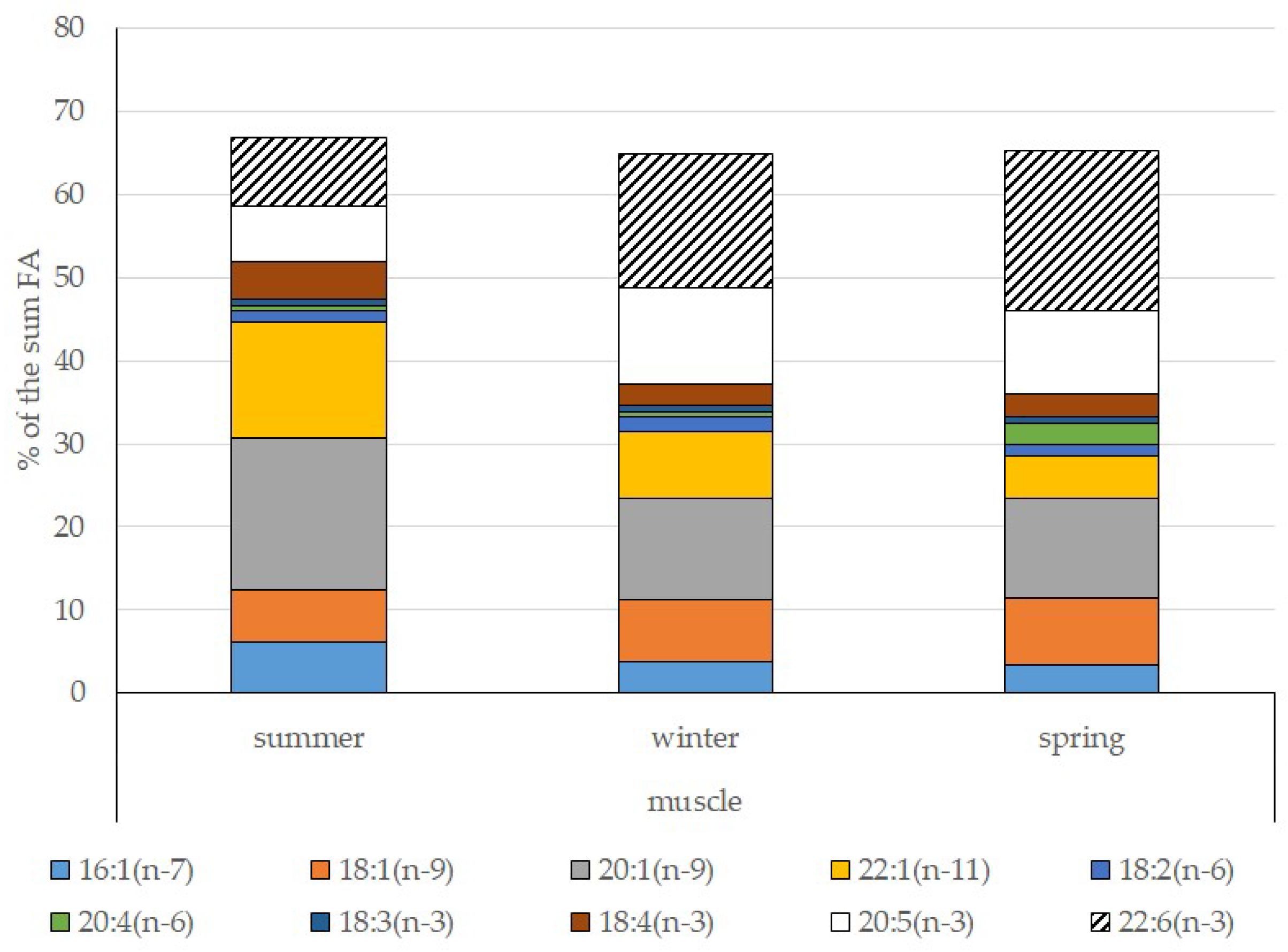
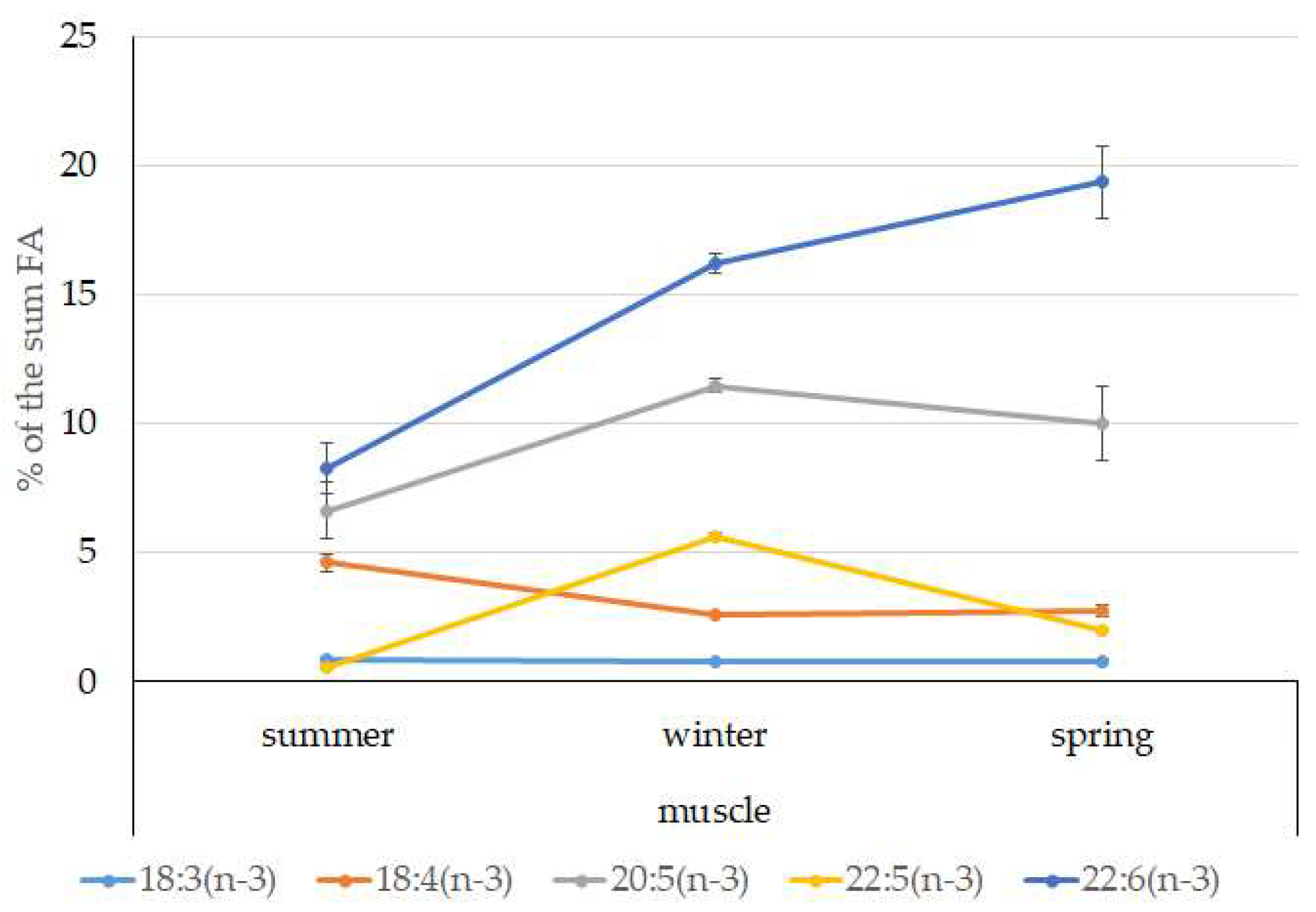
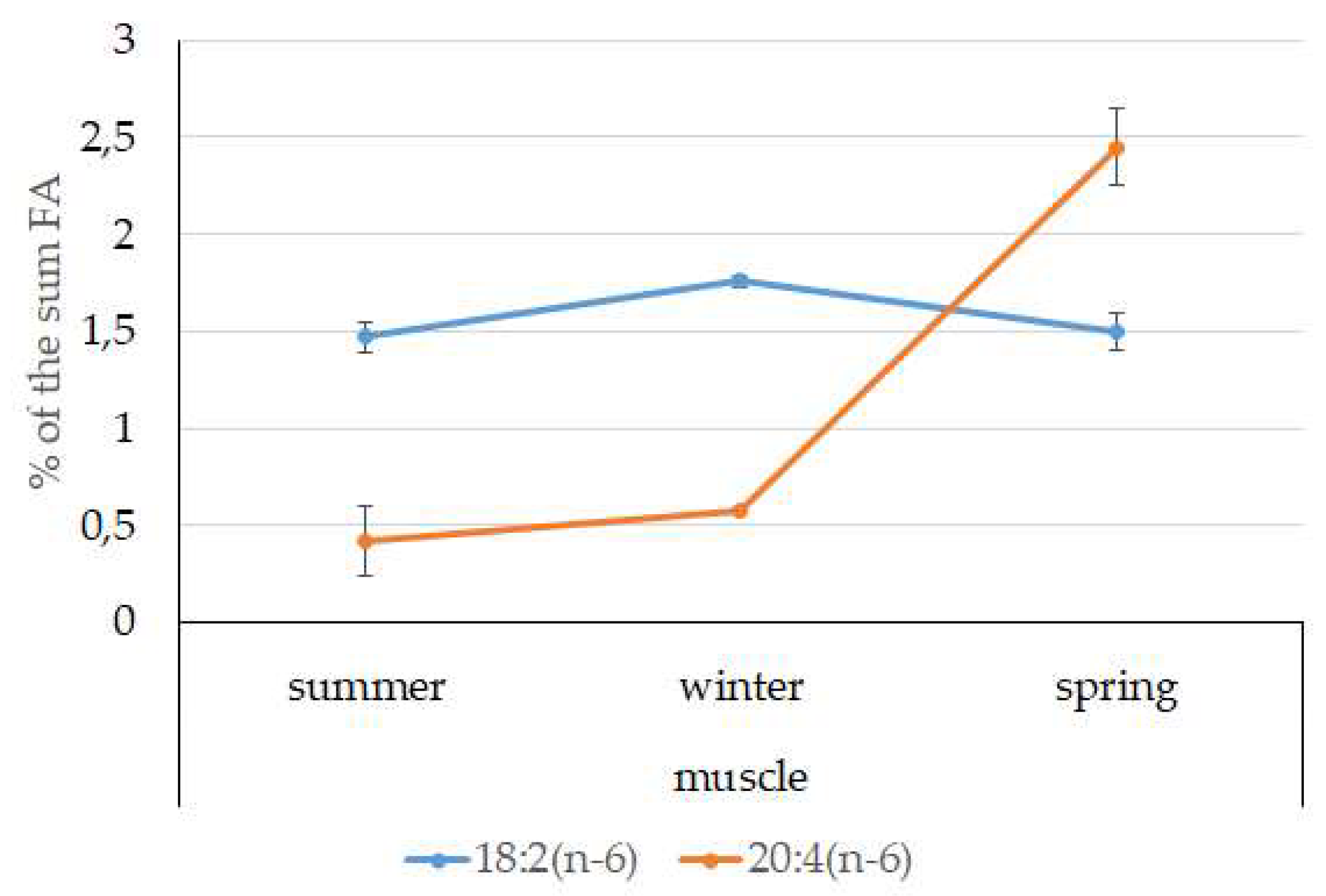
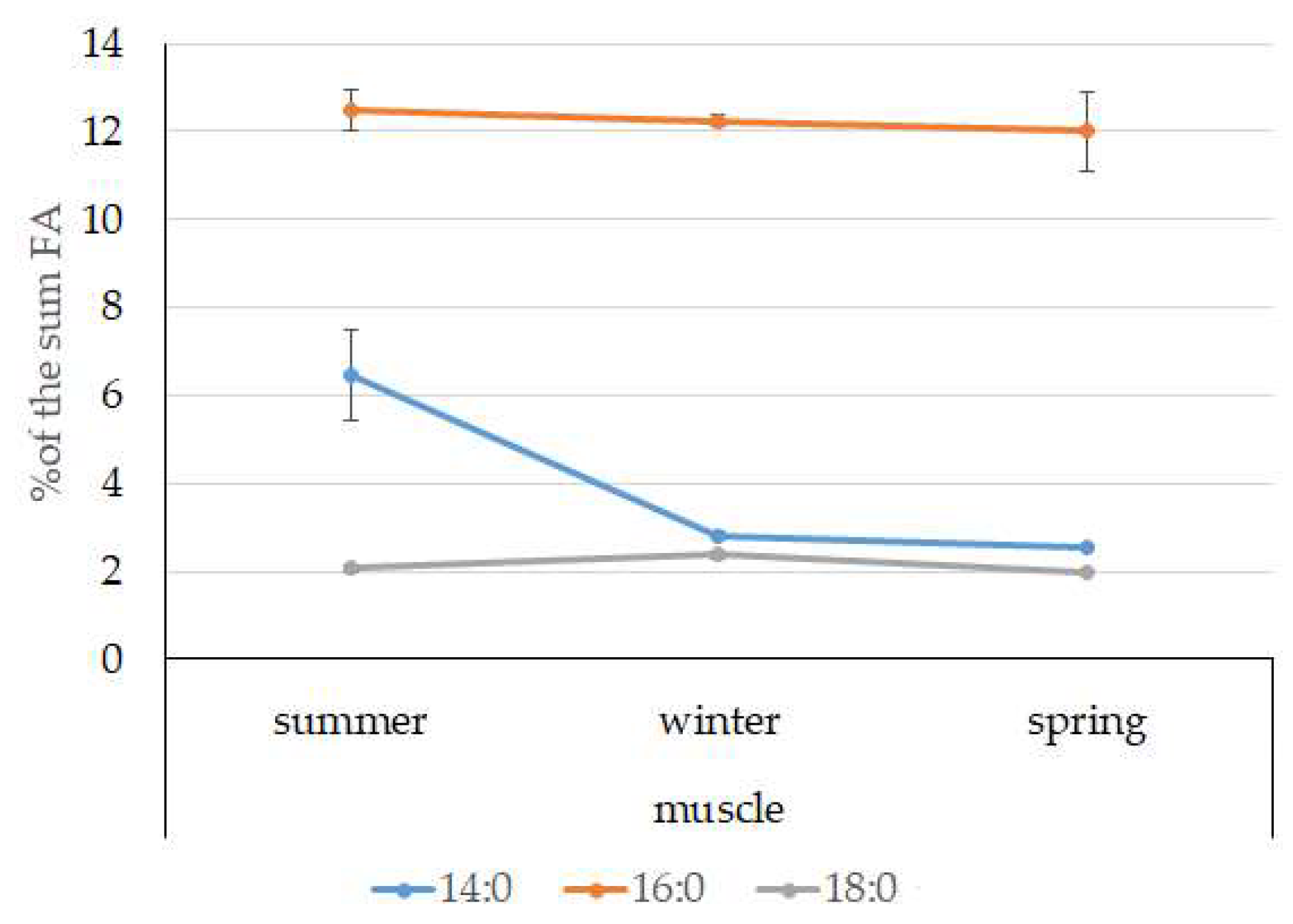
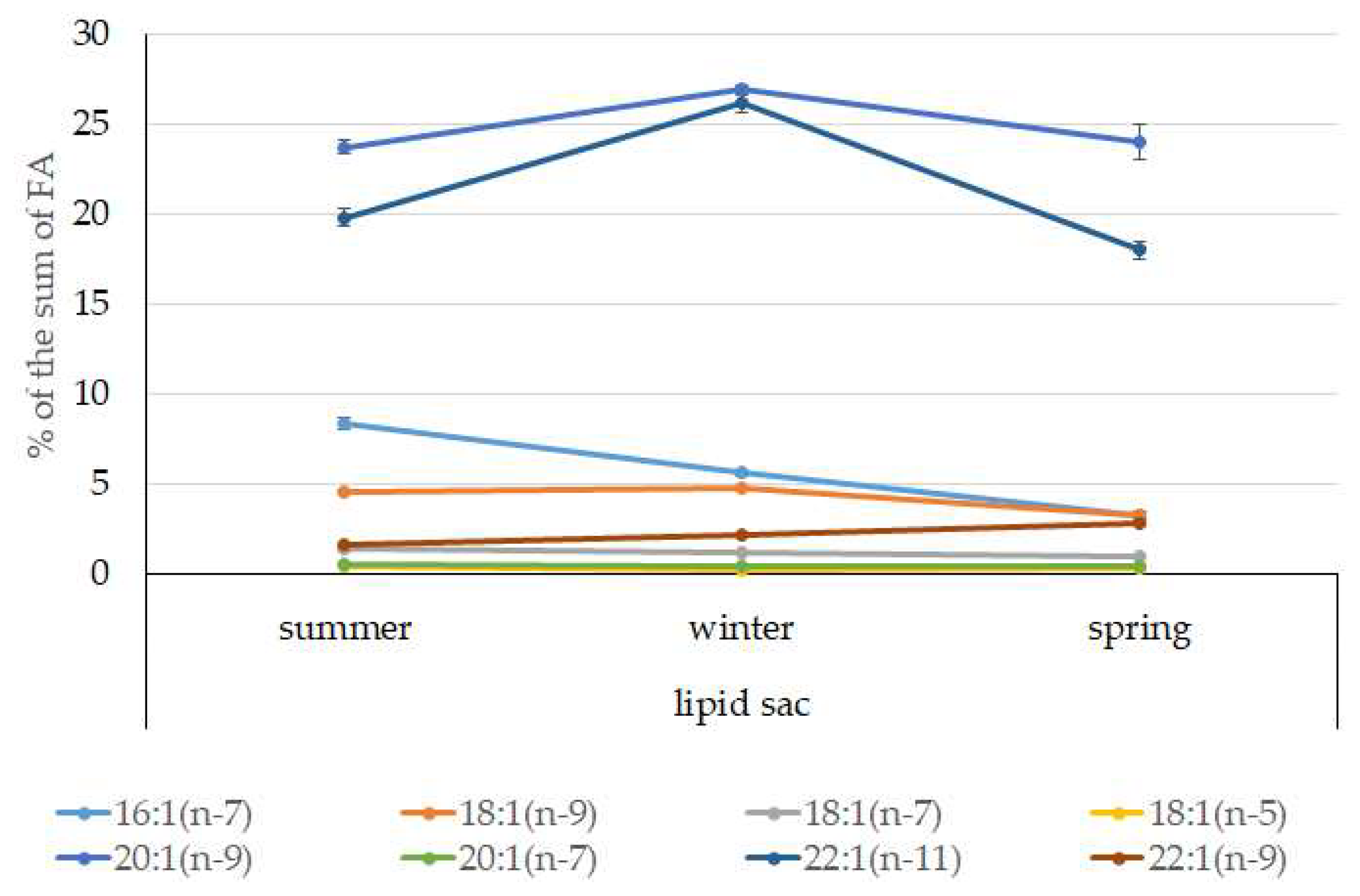
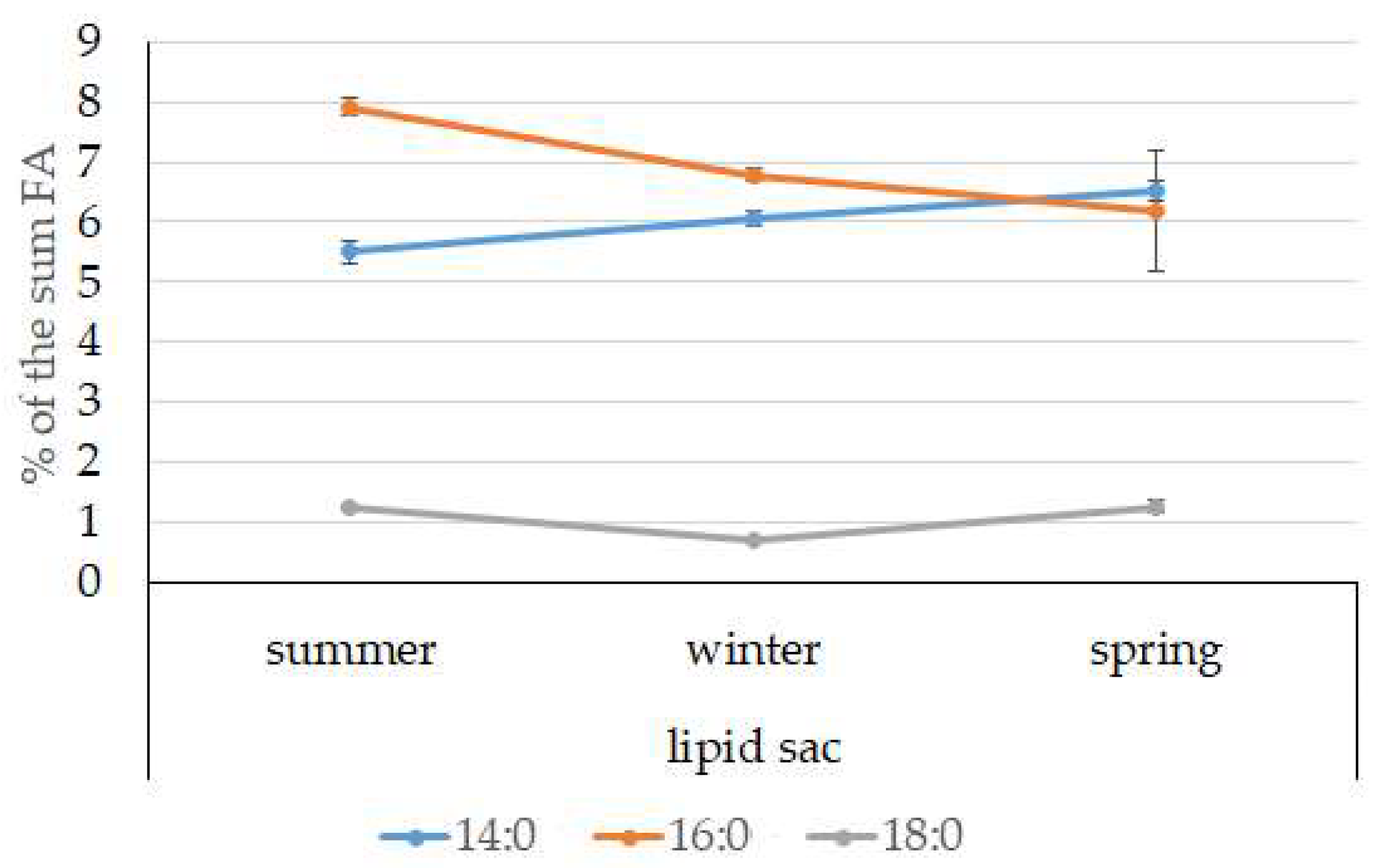
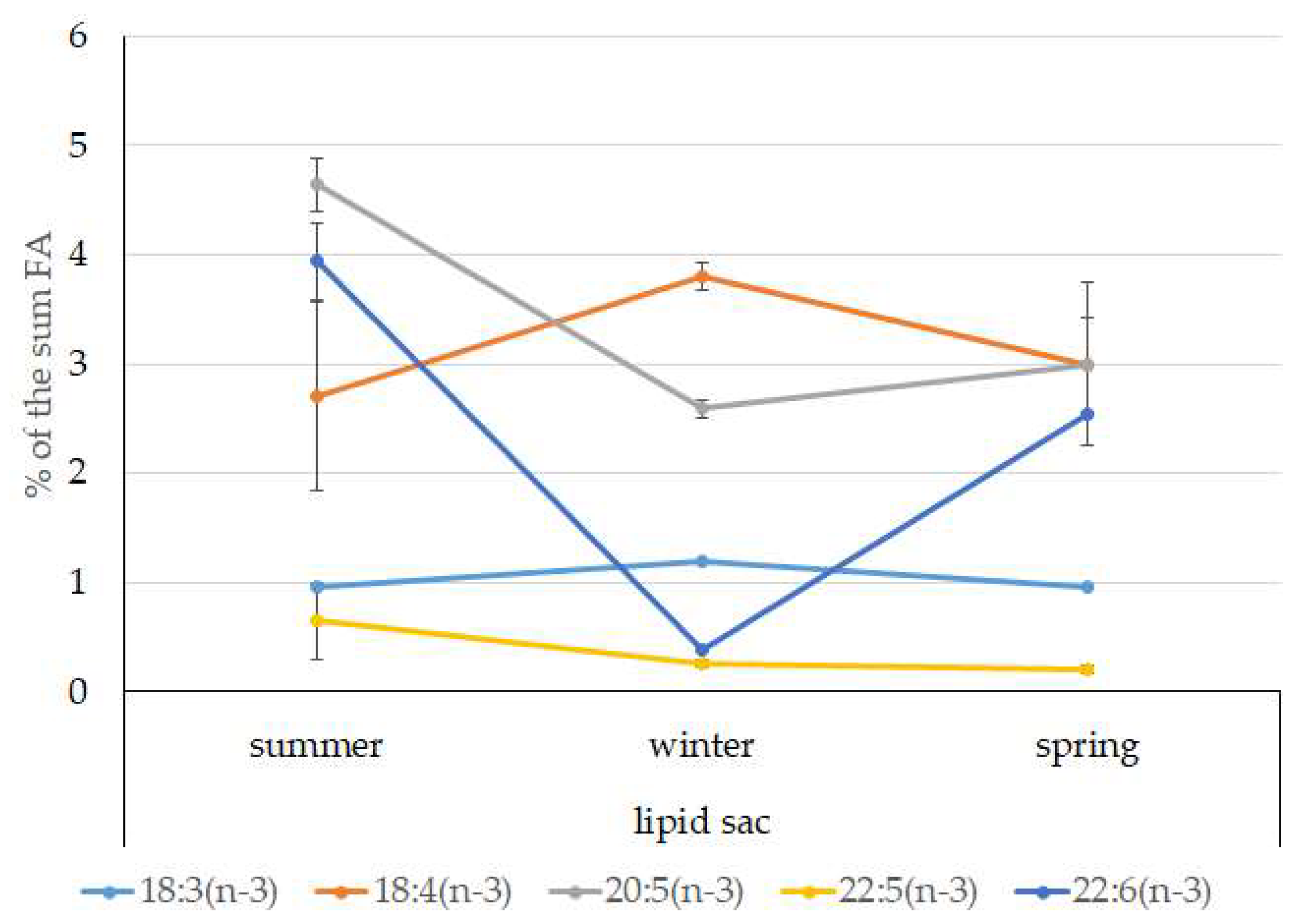
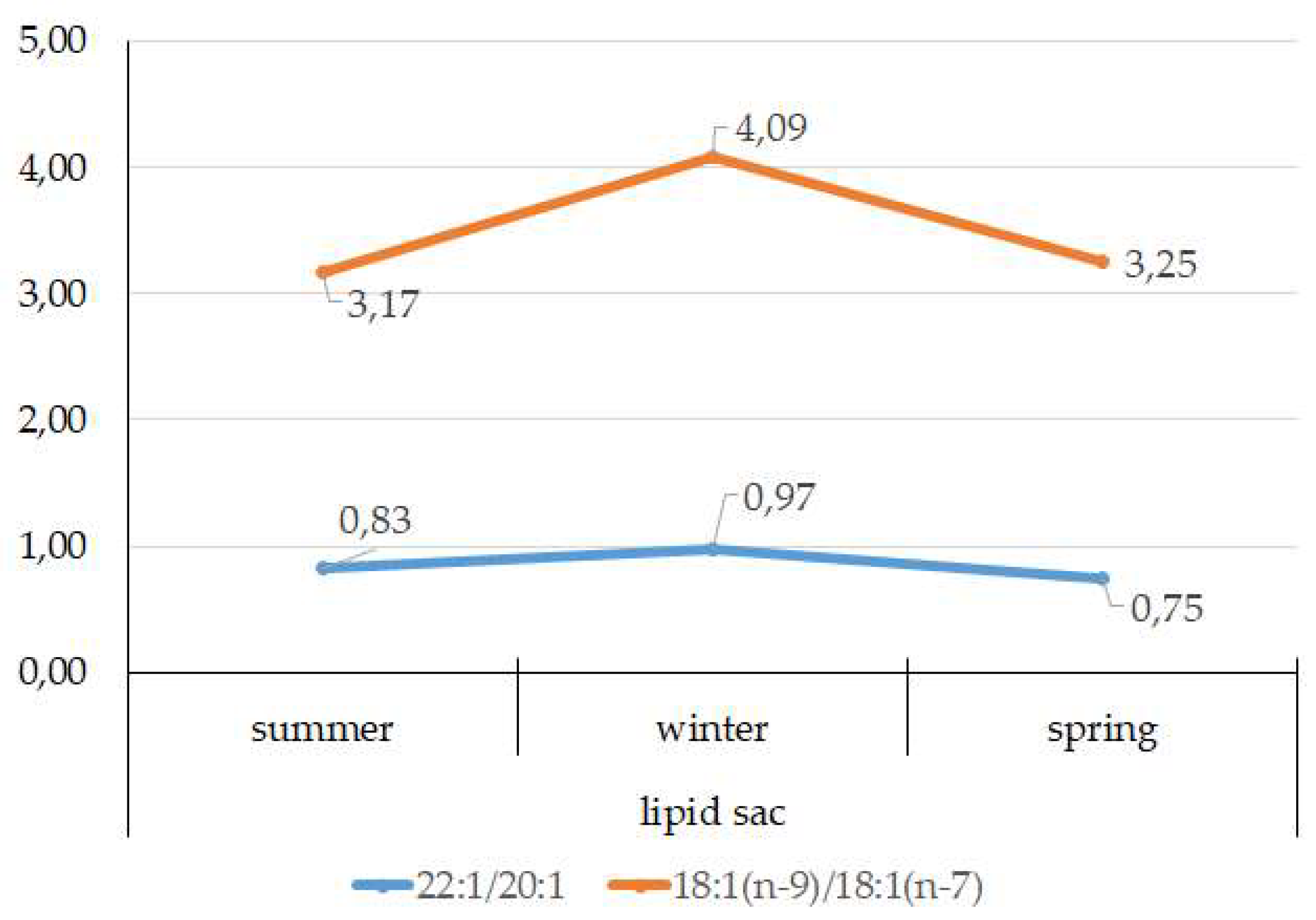
3.4. Seasonal Dynamics of Certain Fatty Acids in the Lipid Sac and Muscles of Pelagic Postlarval L. Maculatus
4. Discussion
4.1. Histomorphology of the Lipid Sac of Postlarval L. maculatus
4.2. Dynamics of Total Lipid and Certain Lipid Classes in the Lipid Sac and Muscles of Postlarval L. maculatus
4.3. Seasonal Dynamic of Certain Fatty Acids of Total Lipids in Postlarval L. maculatus
5. Conclusions
- The presence of a unique formation, in the form of the lipid sac, is a specific ecological and biochemical adaptation in the early development of daubed shanny. The lipid sac symplast of the juvenile daubed shanny is a structure that was discovered and described for the first time;
- The lipid sac accumulates a large amount of lipids, mainly TAGs and specific MUFAs of food origin, 20:1(n-11) and 22:1(n-9), indicating that Calanus spp., which form the basis of the Arctic zooplankton biomass, dominate in the diet of juveniles. The established seasonal dynamics and changes in the ratios of these FAs indicate different species composition and accessibility of copepods in the studied fjords (mainly Kongsfjord, Svalbard) for the pelagic juveniles of the daubed shanny. Tracking the movement of food-derived FAs determines the basis for the chemoindication of qualitative and quantitative relationships between zooplankton and zooplanktophages in Arctic marine food chains;
- A slight but significant increase in the level of derived from food FAs in the lipid sac in spring is synchronized with the beginning of the spring bloom of phytoplankton and the appearance of stage V–VI copepods in the pelagic water layers to start a reproductive period. Interestingly, the inclusion of long-chain 20:1(n-9) and 22:1(n-11) FAs in the structure of energy lipids (mainly TAGs) of the lipid sac of the daubed shanny maintains their energy value, which brings on the corresponding needs of juveniles, even at low temperatures during the long winter period in the Arctic;
- A minor class of energy lipids in the form of Chol esters and waxes is only found in the lipid sac of juveniles in the winter–spring period, which reflects the peculiarities of their nutrition and lipid accumulation.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kreps, E.M. Lipids of Cell Membranes, Evolution of Brain Lipids, Adaptive Function of Lipids; Science: St. Petersburg, Russia, 1981; p. 339. [Google Scholar]
- Boldyrev, A.A.; Prokop’eva, V.D. How the activity of membrane enzymes is regulated? Biol. Sci. 1985, 3, 5–13. [Google Scholar]
- Lehninger, A. Principles of Biochemistry; Mir: Moscow, Russia, 1985; p. 367. [Google Scholar]
- Gennis, R. Biomembranes. Molecular Structure and Function; Mir: Moscow, Russia, 1997; p. 624. [Google Scholar]
- Dyatlovitskaya, E.V.; Bezuglov, V.V. Lipids as bioeffectors. Introduction. Biochemistry 1998, 63, 3–6. [Google Scholar]
- Sergeeva, M.G.; Varfolomeeva, A.T. Arachidonic Acid Cascade; Narodnoe obrazovanie: Moscow, Russia, 2006; p. 256. [Google Scholar]
- Pavlov, D.S.; Nemova, N.N.; Kirillova, E.A.; Kirillov, P.I.; Nefedova, Z.A.; Murzina, S.A. Lipid content in the young-of-the-year sockeye salmon Oncorhynchus nerka during feeding migration (the Ozernaya River, Western Kamchatka). Dokl. Biol. Sci. 2012, 445, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R.; Fonseca-Madrigal, J.; Bell, J.G.; Dick, J.R.; Henderson, R.J.; Sargent, J.R. Effect of diets containing linseed oil on fatty acid desaturation and oxidation in hepatocytes and intestinal enterocytes in Atlantic salmon (Salmo salar). Fish. Physiol. Biochem. 2002, 26, 157–170. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford University Press: New York, NY, USA, 2002; p. 466. [Google Scholar]
- Radchenko, O.A.; Chereshnev, I.A.; Petrovskaya, A.V. Genetic differentiation of species and taxonomic structure of the superfamily Stichaeoidea (Perciformes: Zoarcoidei). Rus. J. Mar. Biol. 2014, 40, 481–492. [Google Scholar] [CrossRef]
- Radchenko, O.A. Molecular Systematics and Phylogeny of the Suborder Zoarcoidei (Pisces, Perciformes); A.N. Severtsov Institute of Ecology and Evolution: Moscow, Russia, 2015. [Google Scholar]
- Radchenko, O.A. Timeline of the evolution of eelpouts from the suborder Zoarcoidei (Perciformes) based on DNA variability. Vopr. Ikhtiol. 2016, 42, 359–367. [Google Scholar] [CrossRef]
- Byrkjedal, I.; Høynes, Å. Distribution of demersal fish in the south-western Barents Sea. Pol. Res. 2007, 26, 135–151. [Google Scholar] [CrossRef]
- Mecklenburg, C.W.; Møller, P.R.; Steinke, D. Biodiversity of Arctic marine fishes: Taxonomy and zoogeography. Mar. Biodivers. 2011, 41, 109–140. [Google Scholar] [CrossRef]
- Møller, P.R.; Gravlund, P. Phylogeny of the eelpout genus Lycodes (Pisces, Zoarcidae) as inferred from mitochondrial cytochrome b and 12S rDNA. Mol. Phylogenet. Evol. 2003, 26, 369–388. [Google Scholar] [CrossRef]
- Mecklenburg, C.W.; Sheiko, B.A. Family Stichaeidae Gill 1864-pricklebacks. Calif. Acad. Sci. Annot. Checkl. Fish. 2004, 35, 1–36. [Google Scholar]
- Dolgov, A.V. Ichthyofauna species composition and ichteocenose structure of the Barents Sea. Izv. TINRO 2004, 137, 177–195. [Google Scholar]
- Dolgov, A.V.; Benzik, A.N. Feeding rations and food consumption of Greenland halibut in the Barents Sea. Vestn. MSTU 2012, 15, 509–516. [Google Scholar]
- Muchina, N.V.; Dolgov, A.V. Composition and structure of the Barents Sea fish community at early ontogenesis stages according to data of Russian ichthyoplankton surveys (1959–1993). Vestn. MSTU 2012, 818–826. [Google Scholar]
- Ochiai, T.; Fuji, A. Energy Transformations by a Blenny (Opisthocentrus ocellatus) Population of Usu Bay, Southern Hokkaido. Bull. Fac. Fish. Hokkaido Univ. 1980, 31, 314–326. [Google Scholar]
- Miki, T.; Yoshida, H.; Amaoka, K. Rare stichaeid fish, Pseudoalectrias tarasovi (Popov), from Japan and its larvae and juveniles. Bull. Fac. Fish. Hokkaido Univ. 1987, 38, 1–13. [Google Scholar]
- Keats, D.W.; Steele, D.H.; Green, J.M.; Martel, G.M. Diet and population size of the Arctic shanny, Stichaeus punctatus (Pisces: Stichaeidae), at sites in eastern Newfoundland and the eastern Arctic. Environ. Biol. Fish. 1993, 37, 173–180. [Google Scholar] [CrossRef]
- Smith, C.; Wootton, R.J. Parental energy expenditure of the male three-spined stickleback. J. Fish Biol. 1999, 54, 1132–1136. [Google Scholar] [CrossRef]
- Kolpakov, N.V.; Klimkin, A.F. Specific features of biology of shannies Stichaeus grigorjewi and S. nozawae (Stichaeidae) in waters of Northern Primorye. J. Ichthyol. 2004, 44, 592–599. [Google Scholar]
- Rose, G.A. On distributional responses of North Atlantic fish to climate change. ICES J. Mar. Sci. 2005, 62, 1360–1374. [Google Scholar] [CrossRef] [Green Version]
- Murzina, S.A.; Falk-Petersen, S.; Nemova, N.N. Lipids in the Arctic fish, Leptoclinus maculatus larvae. Chem. Phys. Lipids 2008, 154, 26. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Falk-Petersen, I.-B.; Sargent, J.R. Structure and function of an unusal lipid storage organ in the Arctic fish Lumpenus maculatus Fries, 1838. Sarsia 1986, 71, 1–6. [Google Scholar] [CrossRef]
- Meyer Ottesen, C.A.; Haakon, H.; Schou Christiansen, J.; Falk-Petersen, S. Early life history of the daubed shanny (Teleostei: Lepticlinus maculatus) in Svalbard waters. Mar. Biodivers. 2011, 41, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Pekkoeva, S.N.; Murzina, S.A.; Ieshko, E.P.; Nefedova, Z.A.; Falk-Petersen, S.; Berge, J.; Lonne, O.; Nemova, N.N. Environmental groups arctic-boreal species Daubed shanny Leptoclinus maculatus (Fries, 1838) in process growth and early development. Rus. J. Ecol. 2018, 3, 225–233. [Google Scholar]
- Murzina, S.A.; Nefedova, Z.A.; Falk-Petersen, S.; Hop, H. Lipids in the daubed shanny (Teleostei: Leptoclinus maculatus) in Svalbard waters. Polar Biol. 2013, 36, 1619–1631. [Google Scholar] [CrossRef]
- Murzina, S.A.; Meyer Ottesen, C.A.; Falk-Petersen, S.; Hop, H.; Nemova, N.N.; Poluektova, O.G. Oogenesis and lipids in gonad and liver of daubed shanny (Leptoclinus maculatus) females from Svalbard waters. Fish Physiol. Biochem. 2012, 38, 1393–1407. [Google Scholar] [CrossRef]
- Pekkoeva, S.N.; Murzina, S.A.; Nefedova, Z.A.; Ripatti, P.O.; Falk-Petersen, S.; Berge, J.; Lonne, O.; Nemova, N.N. Ecology role lipids and fatty acids in early postembryonic development Daubed shanny Leptoclinus maculatus (Fries, 1838) from Kongsfjord (isl. West Svalbard) in winter period. Rus. J. Ecol. 2017, 3, 186–191. [Google Scholar] [CrossRef]
- Pekkoeva, S.N.; Murzina, S.A.; Nefedova, Z.A.; Ruokolaynen, T.R.; Falk-Petersen, S.; Berge, J.; Lønne, O.J.; Nemova, N.N. Role phospholipids in development young arctic-boreal species Leptoclinus maculatus (Stichaeidae). J. Ichthyol. 2017, 57, 467–471. [Google Scholar] [CrossRef]
- Nilssen, K.T.; Haug, T.; Potelov, V.; Timoshenko, Y.K. Food habits and food availability of harp seal (Phoca groenlandica) during early summer and autumn in the northern Barents Sea. Polar Biol. 1995, 15, 485–493. [Google Scholar] [CrossRef]
- Hanson, J.M.; Chouinard, G.A. Diet of Atlantic cod in the southern Gulf of St Lawrence as an index of ecosystem change, 1959–2000. J. Fish Biol. 2002, 60, 902–922. [Google Scholar]
- Dahl, T.M.; Falk-Petersen, S.; Gabrielsen, G.W. Lipids and stable isotopes in common eider, black-legged kittiwake and northern fulmar: A trophic study from an Arctic fjord. Mar. Ecol. Prog. Ser. 2003, 256, 257–269. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Leu, E.; Berge, J.; Kwasniewski, S.; Nygard, H.; Rostad, A.; Keskinen, E.; Thormar, J.; Von Quillfeldt, C.; Wold, A.; et al. Vertical migration in high Arctic waters during autumn 2004. Deep-Sea Res. Part II 2008, 55, 2275–2285. [Google Scholar] [CrossRef]
- Golet, G.H.; Kuletz, K.J.; Roby, D.D.; Irons, D.B. Adult prey choice affects chick growth and reproductive success in pigeon guillemots. Auk 2000, 117, 82–91. [Google Scholar] [CrossRef]
- Meyer Ottesen, C.A.; Haakon, H.; Falk-Petersen, S.; Christiansen, J.S. Growth of daubed shanny (Teleostei: Leptoclinus maculatus) in Svalbard waters. Polar Biol. 2014, 37, 809–815. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissue. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Sidorov, V.S.; Lisenko, E.I.; Bolgova, O.M.; Nefedova, Z.A. Lipids of fish. I. Methods of analyses. The tissues specificity of vebdace lipids (Coregonus albula L.). In Salmonidae of Karelia. Issue 1 Ecology. Parasites. Biochemistry; Potapova, O.I., Smirnov, Y.A., Eds.; Karelian Branch of the USSR Academy of Sciences, Institute of Biology: Petrozavodsk, Russia, 1972; pp. 150–161. [Google Scholar]
- Walsh, D.E.; Banasik, O.J.; Gilles, K.A. Thin-layer chromatographic separation and colorimetric analysis of barley or malt lipid classes and their fatty acids. J. Chromatogr. 1965, 17, 278–287. [Google Scholar] [CrossRef]
- Engelbrecht, F.M.; Mari, F.; Anderson, J.T. Cholesterol determination in serum. A rapid direction method. S. Afr. Med. J. 1974, 48, 250–356. [Google Scholar]
- Arduini, A. High performance liquid chromatography of long-chain acylcarnitine and phospholipids in fatty acid turnover studies. J. Lipid Res. 1996, 37, 684–689. [Google Scholar]
- Jamieson, G.R. GLC-identification techniques for longchain unsaturated fatty acids. J. Chromatogr. Sci. 1975, 13, 491–497. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Berdchenko, I.V. Mesozooplankton waters of West Svalbard. PhD Thesis, Murmask marine biological Institute KRC RAS (Higher Attestation Committee of the Russian Federation), Murmansk, Russia, 2009. [Google Scholar]
- Hop, H.; Wiencke, C. The Ecosystem of Kongsfjord, Svalbard; Springer International Publishing: New York, NY, USA, 2019; p. 562. [Google Scholar]
- Los, D.A. Structure, regulation of expression and functioning of fatty acid desaturases. Biol. Chem. Rev. 2001, 41, 163–198. [Google Scholar]
- Cossins, A.R.; Prosser, C.L. Evolutionary adaptation of membranes to temperature. Proc. Natl. Acad. Sci. USA 1978, 75, 2040–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidorov, V.S. Ecological Biochemistry of Fish; Science: Leningrad, Russia, 1983; p. 240. [Google Scholar]
- Arts, M.T.; Brett, M.T.; Kainz, M.J. Lipids in Aquatic Ecosystems; Springer: New York, NY, USA, 2009; p. 377. [Google Scholar]
- Kattner, G.; Hagen, W.; Lee, R.F.; Cambell, R.; Deibel, D.; Falk-Petersen, S.; Graeve, M.; Hansen, B.W.; Hirche, H.J.; Jónasdottir, S.H.; et al. Perspectives on marine zooplankton lipids. Can. J. Fish. Aquat. Sci. 2007, 64, 1628–1639. [Google Scholar] [CrossRef] [Green Version]
- Kostetsky, E.Y.; Velansky, P.V.; Sanina, N.M. Phospholipid and fatty acid composition of phosphatidylcholine and phosphatidylethanolamine in the black plaice Pleuronectes obscura during thermoadaptation. J. Evol. Biochem. Physiol. 2018, 54, 96–104. [Google Scholar] [CrossRef]
- Kostetsky, E.Y.; Velansky, P.V.; Sanina, N.M. Phase transitions of phospholipids as a criterion for assessing the capacity for thermal adaptation in fish. Rus. J. Mar. Biol. 2013, 39, 136–143. [Google Scholar] [CrossRef]
- Murzina, S.A. Lipids Role and the Fatty Acids Components in Biochemical Adaptation Daubed Shanny Leptoclinus maculatus F. Northwest Coast of Svalbard. Ph.D. Thesis, Karelian State Pedagogical Academy (Higher Attestation Committee of the Russian Federation), Petrozavodsk, Russia, 2010. [Google Scholar]
- Sargent, J.R.; Henderson, R.J. Biological Chemistry of Marine Copepod. In Lipids; Corner, E.D.S., O’Hara, S., Eds.; Oxford University Press: Oxford, UK, 1986; pp. 59–108. [Google Scholar]
- Pekkoeva, S.N. Changes in the Lipid Composition of Muscles and the Lipid Sac of a Representative of the Arctic Ichthyofauna Daubed Shanny Leptoclinus maculatus in the Postembryonic Development. Ph.D. Thesis, I.M. Sechenov Institute of evolutionary physiology and biochemistry (Higher Attestation Committee of the Russian Federation), St. Petersburg, Russia, 2018. [Google Scholar]
- Loeb, V.J.; Kellermann, A.; Koubbi, P.; North, A.W.; White, M. Antarctic larval fish assemblages: A review. Bull. Mar. Sci. 1993, 53, 416–449. [Google Scholar]
- Koubbi, P.; Duhamel, G.; Hecq, J.H.; Beans, C.; Loots, C.; Pruvost, P.; Tavenier, E.; Vacchi, M.; Vallet, C. Ichthyoplankton in the neritic and coastal zone of Antarctica and Subantarctic islands: A review. J. Mar. Syst. 2009, 78, 547–556. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Hopkins, C.E.; Sargent, J.R. Trophic relationships in the pelagic, arctic food web. Trophic relationships in the Marine Environment. In Proceedings of the 24th European Marine Biology Symposium, Aberdeen, Scotland; Aberdeen University Press: Aberdeen, Scotland, 1990; pp. 315–333. [Google Scholar]
- Mayzaud, P.; Chevallier, J.; Tavernier, E.; Moteki, M.; Koubbi, P. Lipid composition of the Antarctic fish Pleuragramma antarcticum. Influence of age class. Polar Sci. 2011, 5, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Moteki, M.; Koubbi, P.; Pruvost, P.; Tavernier, E.; Hulley, P.A. Spatial distribution of pelagic fish off Ade’lie and George V Land, East Antarctica in the austral summer 2008. Polar Sci. 2011, 5, 211–224. [Google Scholar] [CrossRef] [Green Version]
- Rabinovich, A.L.; Ripatti, P.O. Polyunsaturated hydrocarbon chains of lipids: Structure, functions and properties. Biol. Chem. Rev. 1994, 114, 581–594. [Google Scholar]
- Rabinovich, A.L.; Ripatti, P.O.; Balabaev, N.K.; Leermakers, F.A.M. Molecular dynamics simulations of hydrated unsaturated lipid bilayers in the liquid-crystal phase and comparison to self-consistent field modeling. Phys. Rev. E 2003, 67, 011909. [Google Scholar] [CrossRef] [Green Version]
- Tocher, D.R. Glycerophospholipid metabolism. In Metabolic and Adaptational Biochemistry. Biochemistry and Molecular Biology of Fishes; Hochachka, P.W., Mommsen, T.P., Eds.; Elsevier Press: Amsterdam, The Netherlands, 1995; pp. 119–157. [Google Scholar]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquacult. Res. 2008, 41, 717–732. [Google Scholar] [CrossRef]
- Piquet, A.M.-T.; Van de Poll, W.H.; Visser, R.J.W.; Wiencke, C.; Bolhuis, H.; Buma, A.G.J. Springtime phytoplankton dynamics in Arctic Krossfjorden and Kongsfjorden (Spitsbergen) as a function of glacier proximity. Biogeoscience 2014, 11, 2263–2279. [Google Scholar] [CrossRef] [Green Version]
- Piquet, A.M.-T.; Bolhuis, H.; Meredith, M.P.; Buma, A.G.J. Shifts in coastal Antarctic marine microbal communities during and after melt water-related surface stratification. FEMS Microbiol. Ecol. 2011, 76, 413–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegseth, E.N.; Tvelberg, V. Effect of Atlantic water inflow on timing of the phytoplankton spring bloom in a high Arctic fjord (Kongsfjorden, Svalbard). J. Mar. Syst. 2013, 113–114, 94–105. [Google Scholar] [CrossRef]
- Sandel, E.; Nixon, O.; Lutzky, S.; Ginsbourg, B.; Tandler, A.; Uni, Z.; Koven, W. The effect of dietary phosphatidylcholine/phosphatidylinositol ratio on malformation in larvae and juvenile gilthead sea bream (Sparus aurata). Aquaculture 2010, 304, 42–48. [Google Scholar] [CrossRef]
- Kondakova, E.A.; Efremov, V.I.; Bogdanova, V.A. Structure of the yolk syncytial layer in the larvae of whitefishes: A histological study. Rus. J. Dev. Biol. 2017, 48, 176–184. [Google Scholar] [CrossRef]
- Lyubartsev, A.P.; Rabinovich, A.P. Force field development for lipid membrane simulations. Biochem. Phys. Acta (BBA)-Biomemnranes 2016, 10, 2483–2497. [Google Scholar] [CrossRef]
- Rabinovich, A.l.; Lyubartsev, A.P. Computer simulation of lipid membranes: Methodology and achievements. Polym. Sci. Ser. C 2013, 55, 805–808. [Google Scholar] [CrossRef]
- Murzina, S.A.; Nefedova, Z.A.; Ripatti, P.O.; Ruokolainen, T.R.; Nemova, N.N.; Falk-Petersen, S.; Pekkoeva, S.N. Lipid status of the two high latitude fish species, Leptoclinus maculatus and Lumpenus fabricii. Int. J. Mol. Sci. 2013, 14, 7048–7060. [Google Scholar] [CrossRef] [Green Version]
- Zhurkin, D.V.; Rabinovich, A.L. Monte Carlo properties of the hydrocarbon chains of phospholipid molecules. Rus. J. Phys. Chem. 2015, 89, 242–249. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Fjord | Kongsfjord | ||
|---|---|---|---|
| Season/Parameter | T (°C) | Salinity (%) | Depth (M) |
| Summer (end of July-beginning of August) | 4.2 | 34.7 | 100 |
| Winter (mid. January) | 1 | 34.8 | 125 |
| Spring (mid. April) | 0.5 | 34.95 | 100 |
| Season | Summer | Winter * | Spring | |||
|---|---|---|---|---|---|---|
| Tissue/Organ | Muscle | Lipid Sac | Muscle | Lipid Sac | Muscle | Lipid Sac |
| n | 15 | 15 | 40 | 40 | 12 | 12 |
| TL | 27.9 ± 3.00b | 64.4 ± 2.1a | 13.9 ± 0.5a | 75.9 ± 2.8b | 13.4 ± 0.4a | 46.9 ± 2.9c |
| TAG | 12.7 ± 1.8a | 58.3 ± 3.1a | 2.4 ± 0.2b | 68.8 ± 2.5b | 6.1 ± 1.0c | 28.1 ± 1.3c |
| Chol esters+wax esters | 1.7 ± 0.4a | 0 | 0.9 ± 0.1b | 3.02. ± 0.4 | 1.7 ± 0.2c | 2.6 ± 1.0 |
| PL | 9.6 ± 1.4a | 6.1 ± 1.7a | 7.2 ± 0.3b | 0.8 ± 0.3b | 2.5 ± 0.3c | 6.3 ± 1.3c |
| PI | 2.3 ± 1.0a | 1.1 ± 0.1a | 3.8 ± 0.2a | 2.4 ± 0.6b | 3.0 ± 0.2a | 2.3 ± 0.5b |
| PS | 1.0 ± 0.1a | 2.0 ± 0.1a | 3.4 ± 0.2b | 3.3 ± 0.2b | 3.1 ± 0.2b | 1.2 ± 0.4c |
| PE | 20.0 ± 1.8a | 18.1 ± 1.3a | 26.7 ± 0.8b | 22.2 ± 1.0b | 24.6 ± 1.0c | 20.1 ± 1.4c |
| PC | 74.5 ± 2.7a | 75.2 ± 2.1a | 64.5 ± 1.1b | 68.2 ± 1.5b | 62.3 ± 2.0b | 66.2 ± 2.0c |
| LPC | 0.3 ± 0.1a | 0.8 ± 0.2a | 0.1 ± 0.0b | 0.9 ± 0.2b | 0.2 ± 0.1b | 1.0 ± 0.8b |
| SM | 0.4 ± 0.0a | 0.5 ± 0.1a | 1.5 ± 0.1b | 2.5 ± 0.5b | 1.0 ± 0.1c | 2.0 ± 0.2b |
| Chol | 3.9 ± 1.2a | 0a | 3.5 ± 0.2a | 3.4 ± 0.4b | 3.1 ± 0.1a | 10.0 ± 2.5c |
| Chol/PL | 0.4 ± 0.1a | 0.01a | 0.5 ± 0.2b | 4.1 ± 1.2b | 1.3 ± 0.1c | 1.6 ± 0.8c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murzina, S.A.; Pekkoeva, S.N.; Kondakova, E.A.; Nefedova, Z.A.; Filippova, K.A.; Nemova, N.N.; Orlov, A.M.; Berge, J.; Falk-Petersen, S. Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters. Biomolecules 2020, 10, 368. https://doi.org/10.3390/biom10030368
Murzina SA, Pekkoeva SN, Kondakova EA, Nefedova ZA, Filippova KA, Nemova NN, Orlov AM, Berge J, Falk-Petersen S. Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters. Biomolecules. 2020; 10(3):368. https://doi.org/10.3390/biom10030368
Chicago/Turabian StyleMurzina, Svetlana A., Svetlana N. Pekkoeva, Ekaterina A. Kondakova, Zinaida A. Nefedova, Kseniia A. Filippova, Nina N. Nemova, Alexei M. Orlov, Jorgen Berge, and Stig Falk-Petersen. 2020. "Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters" Biomolecules 10, no. 3: 368. https://doi.org/10.3390/biom10030368
APA StyleMurzina, S. A., Pekkoeva, S. N., Kondakova, E. A., Nefedova, Z. A., Filippova, K. A., Nemova, N. N., Orlov, A. M., Berge, J., & Falk-Petersen, S. (2020). Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters. Biomolecules, 10(3), 368. https://doi.org/10.3390/biom10030368