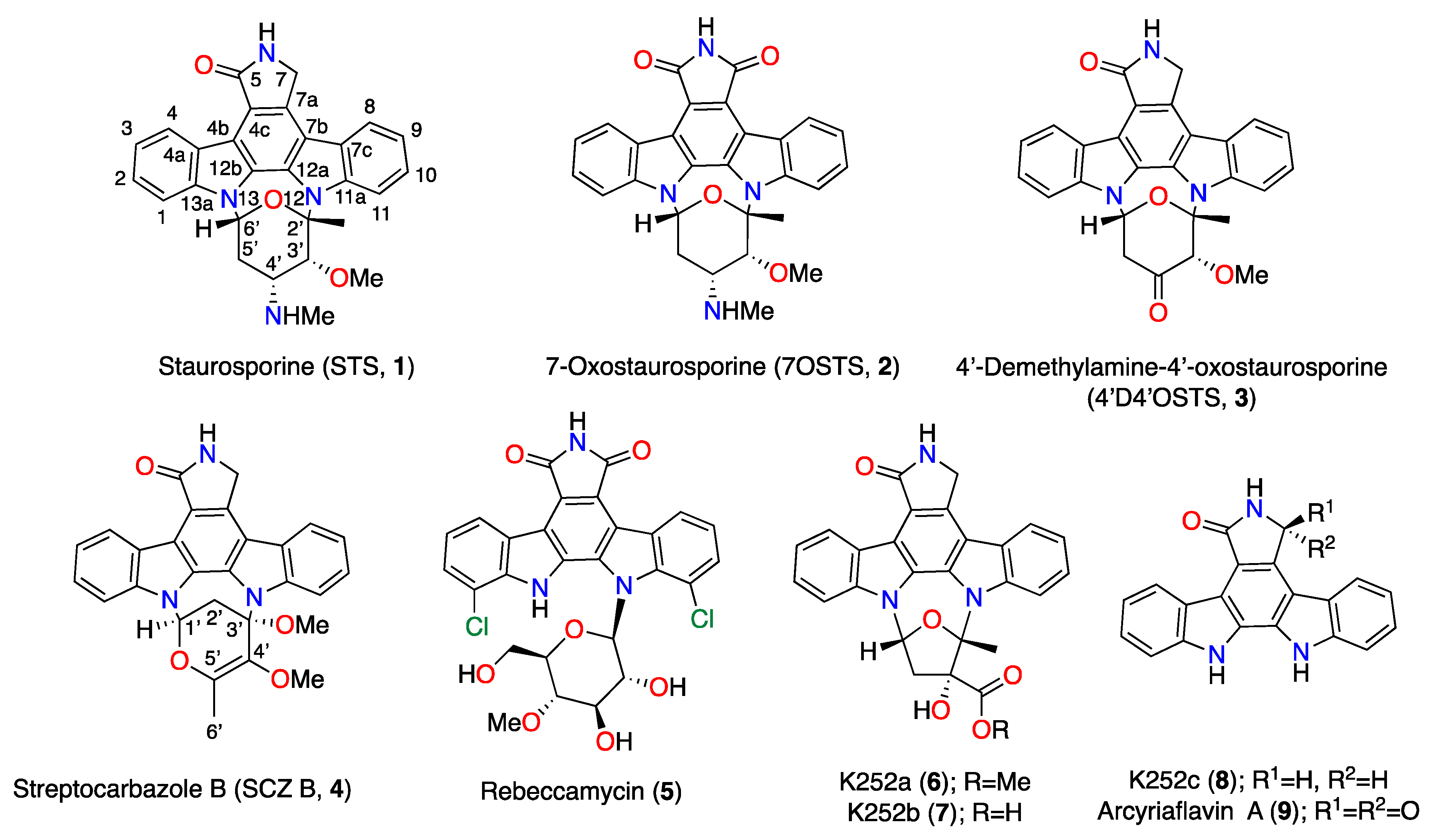
Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis

 ,
, 


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Methods
2.2. Biological Material, Culture and Bioassay-Guided Isolation of Natural ICZ Metabolites 1‒4
2.3. Commercial ICZ Analogs 5–9
2.4. Parasite Strain
2.5. Evaluation of Leishmanicidal, Trypanocidal and Cytotoxic Activities
2.5.1. Leishmanicidal Capacity Assay
2.5.2. Trypanocidal Capacity Assay
2.5.3. Cytotoxicity Assay
2.6. Mechanisms of Cell Death
2.6.1. Plasma Membrane Permeability
2.6.2. Analysis of Mitochondrial Membrane Potential
2.6.3. Measurement of ATP
2.7. Statistical Analysis
3. Results
3.1. ICZ Metabolites: Natural Source and Bioassay-Guided Isolation
3.2. Antiparasitic Assays
Antikinetoplastid Activities
3.3. Mechanisms of Cell Death
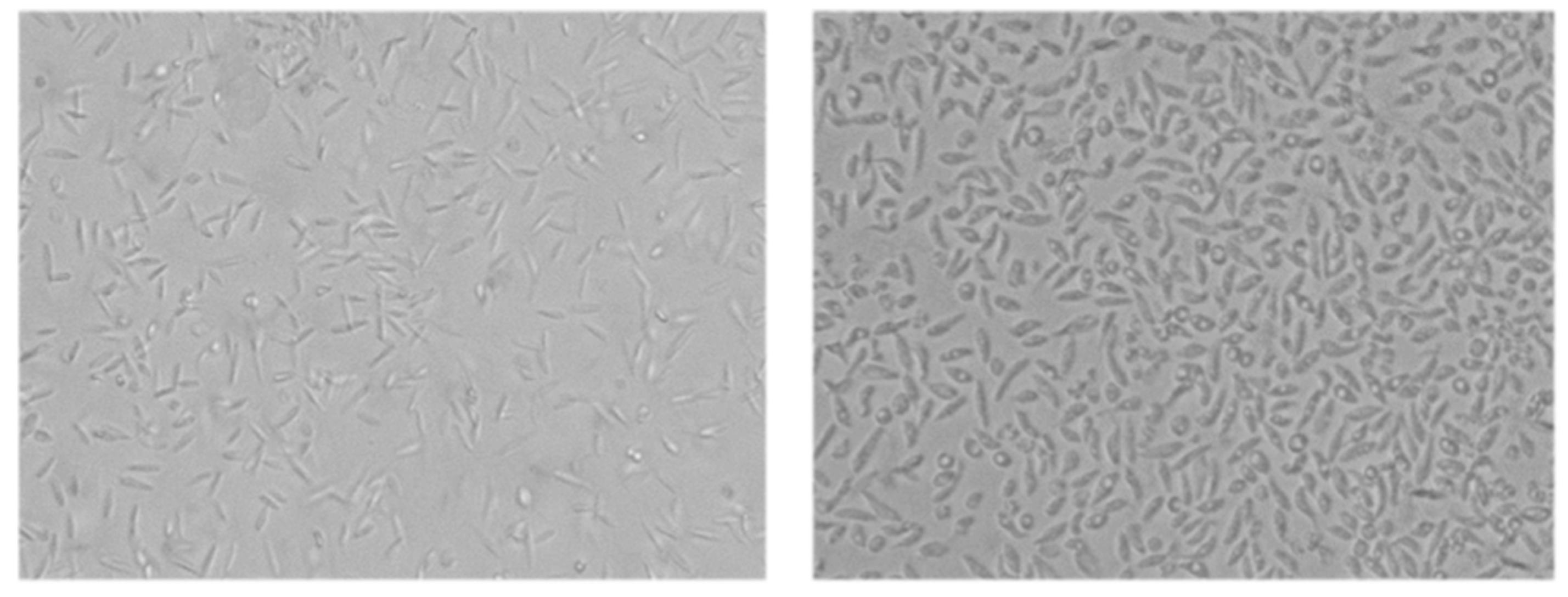
3.3.1. Mitochondrial Damage in Leishmania amazonensis Induced by 7-oxostaurosporine (2)
3.3.2. Cytoplasmic Membrane Permeability in Leishmania donovani and Trypanosoma cruzi Induced by 7-oxostaurosporine (2)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stolk, W.A.; Kulik, M.C.; le Rutte, E.A.; Jacobson, J.; Richardus, J.H. Between-Country Inequalities in the Neglected Tropical Disease Burden in 1990 and 2010, with Projections for 2020. PLoS Negl. Trop. Dis. 2016, 10, e0004560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackey, T.K.; Liang, B.A.; Cuomo, R.; Hafen, R.; Brower, K.C.; Lee, D.E. Emerging and Reemerging Neglected Tropical Diseases: A Review of Key Characteristics, Risk Factors, and the Policy and Innovation Environment. Clin. Microbiol. Rev. 2014, 27, 949–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, K.; Brun, R.; Croft, S.; Fairlamb, A.; Gürtler, R.E.; McKerrow, J.; Reed, S.; Tarleton, R. Kinetoplastids: Related protozoan pathogens, different diseases. J. Clin. Investig. 2008, 118, 1301–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filardy, A.A.; Guimarães-Pinto, K.; Nunes, M.P.; Zukeram, K.; Fliess, L.; Pereira, L.; Oliveira, D.; Conde, L.; Morrot, A. Human Kinetoplastid Protozoan Infections: Where Are We Going Next? Front. Immunol. 2018, 9, 1493. [Google Scholar] [CrossRef]
- Pereira Nunes, M.C.; Dones, W.; Morillo, C.A.; Encina, J.J.; Ribeiro, A.L. Chagas Disease: An Overview of Clinical and Epidemiological Aspects. J. Am. Coll. Cardiol. 2013, 62, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization. Guidelines for the Diagnosis and Treatment of Chagas Disease; PAHO: Washington, DC, USA, 2019; Available online: http://iris.paho.org/xmlui/bitstream/handle/123456789/49653/9789275120439_eng.pdf?sequence=6&isAllowed=y (accessed on 11 February 2020).
- Murillo, C.A.; Waskin, H.; Sosa-Estani, S.; Bangher, M.; Cuneo, C.; Milesi, R.; Mallagray, M.; Apt, W.; Beloscar, J.; Gascon, J.; et al. Benznidazole and posaconazole in eliminating parasites in asymptomatic T. cruzi carriers: The STOP-CHAGAS Trial. J. Am. Coll. Cardiol. 2017, 69, 939–947. [Google Scholar] [CrossRef]
- Merritt, C.; Silva, L.E.; Tanner, A.L.; Stuart, K.; Pollastri, M.P. Kinases as Druggable Targets in Trypanosomatid Protozoan Parasites. Chem. Rev. 2014, 114, 11280–11304. [Google Scholar] [CrossRef] [Green Version]
- Braga, M.V.; deSouza, W. Effects of protein kinase and phosphatidylinositol-3 kinase inhibitors on growth and ultrastructure of Trypanosoma cruzi. FEMS Microbiol. Lett. 2006, 256, 209–216. [Google Scholar] [CrossRef]
- Naula, C.; Parsons, M.; Mottram, J.C. Protein kinases as drug targets in trypanosomes and Leishmania. Biochim. Biophys. Acta 2005, 1754, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Hassan, P.; Fergusson, D.; Grant, K.M.; Motram, J.C. The CRK3 protein kinase is essential for cell cycle progression of Leishmania mexicana. Mol. Biochem. Parasitol. 2001, 113, 189–198. [Google Scholar] [CrossRef]
- Willey, S.; Thomas, M.; Patterson, S.; Crouch, S.; De Rycker, M.; Lowe, R.; Gresham, S.; Urbaniak, M.; Otto, T.; Stojanovski, L.; et al. Cyclin-dependent kinase 12 is a drug target for visceral leishmaniasis. Nature 2018, 560, 192–197. [Google Scholar]
- Grant, K.M.; Dunion, M.H.; Yardley, V.; Skaltsounis, A.-L.; Marko, D.; Eisenbrand, G.; Croft, S.L.; Meijer, L.; Mottram, J.C. Inhibitors of Leishmania mexicana CRK3 Cyclin-Dependent Kinase: Chemical Library Screen and Antileishmanial Activity. Antimicrob. Agents Chemoter. 2004, 48, 3033–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña, I.; Manzano, M.P.; Cantizani, J.; Kessler, A.; Alonso-Padilla, J.; Bardera, A.I.; Alvarez, E.; Colmenarejo, G.; Cotillo, I.; Roquero, I.; et al. New Compound Sets Identified from High Throughput Phenotypic Screening Against Three Kinetoplastid Parasites: An Open Resource. Sci. Rep. 2015, 5, 8771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welburn, S.C.; Macleod, E.; Figarella, K.; Duzensko, M. Programmed cell death in African trypanosomes. Parasitology 2006, 132, S7–S18. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Howe, J.; Tan, K. Staurosporine-induced programmed cell death in blastocystis occurs independently of caspases and cathepsins and is augmented by calpain inhibition. Microbiology 2010, 156, 1284–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruges, G.; Betancourt, M.; March, M.; Sanchez, E.; Mijares, A. Apoptotic-like activity of staurosporine in axenic cultures of Trypanosoma evansi. Rev. do Inst. de Med. Trop. de São Paulo 2012, 54, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Barth, T.; Bruges, G.; Meiwes, A.; Mogk, S.; Mudogo, C.N.; Duszenko, M. Staurosporine-Induced Cell Death in Trypanosoma brucei and the role of Endonuclease G during Apoptosis. Open J. Apoptosis 2013, 3, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Foucher, A.L.; Rachidi, N.; Gharbi, S.; Blisnik, T.; Bastin, P.; Pemberton, I.K.; Späth, G. Apoptotic Marker Expression in the Absence of Cell Death in Staurosporine-Treated Leishmania donovani. Antimicrob. Agents Chemother. 2013, 57, 1252–1261. [Google Scholar] [CrossRef] [Green Version]
- Osada, H.; Koshino, H.; Kudo, T.; Onose, R.; Isono, K. A new inhibitor of protein kinase C, RK-1409 (7-oxostaurosporine) I. Taxonomy and biological activity. J. Antibiot. 1992, 45, 189–194. [Google Scholar] [CrossRef]
- Fabre, S.; Prudhomme, M. Protein Kinase C Inhibitors; Structure-Activity Relationships in K252c-Related Compounds. Bioorg. Med. Chem. 1993, 1, 193–196. [Google Scholar] [CrossRef]
- Fabre, S.; Prudhomme, M.; Sancelme, M.; Rapp, M. Indolocarbazole Protein Kinase C Inhibitors from Rebeccamycin. Bioorg. Med. Chem. 1994, 2, 73–77. [Google Scholar] [CrossRef]
- Cartuche, L.; Sifaoui, I.; Cruz, D.; Reyes-Batlle, M.; Lopez-Arencibia, A.; Fernández, J.J.; Diaz-Marrero, A.R.; Piñero, J.E.; Lorenzo-Morales, J. Staurosporine from Streptomyces sanyensis activates Programmed Cell Death in Acanthamoeba via the mitochondrial pathway and presents low cytotoxicity levels to vertebrate cells. Sci. Rep. 2019, 9, 11651. [Google Scholar] [CrossRef]
- Cartuche, L.; Reyes-Batlle, M.; Sifaoui, I.; Arberas-Jiménez, I.; Piñero, J.E.; Fernández, J.J.; Lorenzo-Morales, J.; Díaz-Marrero, A.R. Antiamoebic Activities of Indolocarbazole Metabolites Isolated from Streptomyces sanyensis Cultures. Mar. Drugs 2019, 17, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sifaoui, I.; López-Arencibia, A.; Martín-Navarro, C.M.; Chammem, N.; Reyes-Battle, M.; Mejri, M.; Lorenzo-Morales, J.; Abderabba, M.; Piñero, J.E. Activity of olive leaf extracts against the promastigote stage of Leishmania species and their correlation with the antioxidant activity. Exp. Parasitol. 2014, 141, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Sahu, R.; Walker, L.A.; Tekwani, B.L. A parasite rescue and transformation assay for antileishmanial screening against intracellular Leishmania donovani amastigotes in THP1 human acute monocytic leukemia cell line. J. Vis. Exp. 2012, 70, 4054. [Google Scholar] [CrossRef] [Green Version]
- Omura, S.; Iwai, Y.; Hirano, A.; Nakagawa, A.; Awaya, J.; Tsuchiya, H.; Takahashi, Y.; Masuma, R. A new alkaloid AM-2282 of Streptomyces origin. Taxonomy, fermentation, isolation and preliminary characterization. J. Antibiot. 1977, 30, 275–282. [Google Scholar] [CrossRef]
- Cai, Y.; Fredenhagen, A.; Hug, P.; Meyer, T. Further minor metabolites of Staurosporine produced by a Streptomyces longisporoflavus strain. J. Antibiot. 1996, 49, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.; Yang, C.; Wang, Y.; Liu, P.; Ma, Y.; Xu, L.; Su, M.; Hong, K.; Zhu, W. Streptocarbazoles A and B, Two Novel Indolocarbazoles from the Marine-Derived Actinomycete Strain Streptomyces sp. FMA. Org. Lett. 2012, 14, 2422–2425. [Google Scholar] [CrossRef]
- Becker, S.; Jaffe, C. Effect of protein kinase inhibitors on the growth, morphology, and infectivity of Leishmania promastigotes. Parasitol. Res. 1997, 83, 273–280. [Google Scholar] [CrossRef]
- Menna-Barreto, R.F.S. Cell death pathways in pathogenic trypanosomatids: Lessons of (over)kill. Cell Death Dis. 2019, 10, 93. [Google Scholar] [CrossRef]
- Yanosik, T.; Rannug, A.; Rannug, U.; Whalström, N.; Slätt, J. Chemistry and properties of indolocarbazoles. Chem. Rev. 2018, 118, 9058–9128. [Google Scholar] [CrossRef] [PubMed]
- Gani, O.A.S.M.; Engh, R.A. Protein kinase inhibition of clinically important staurosporine analogues. Nat. Prod. Rep. 2010, 27, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, A.M.; Noble, M.E.M.; Tunnah, P.; Brown, N.R.; Johnson, L.N.; Endicott, J.A. Protein Kinase Inhibition by Staurosporine Revealed in Details of the Molecular Interaction with CDK2. Nat. Struct. Biol. 1997, 4, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Tanramluk, D. On the Origins of Enzyme Inhibitor Selectivity and Promiscuity: A Case of Study of Protein Kinase Binding to Staurosporine. Doctoral Dissertation, University of Cambridge, Cambridge, UK, 2009. [Google Scholar]
- Pereira, E.R.; Belin, L.; Sancelme, M.; Prudhomme, M.; Ollier, M.; Rapp, M.; Sevère, D.; Riou, F.F.; Fabbro, D.; Meyer, T. Structure-activity relationships in a series of substituted indolocarbazoles: Topoisomerase I and protein kinase C inhibition and antitumoral and antimicrobial properties. J. Med. Chem. 1996, 39, 4471–4477. [Google Scholar] [CrossRef]
- Sunter, J.D.; Yanase, R.; Wang, Z.; Catta-Pretta, C.M.C.; Moreira-Leite, F.; Myskova, J.; Pruzinova, K.; Volf, P.; Mottram, J.C.; Gull, K. Leishmania flagellum attachment zone is critical for flagellar pocket shape, development in the sandfly, and pathogenicity in the host. PNAS 2019, 116, 6351–6360. [Google Scholar] [CrossRef] [Green Version]
- Katta, S.S.; Tammana, T.V.; Sahasrabuddhe, A.A.; Bajpai, V.K.; Gupta, C.M. Trafficking activity of myosin XXI is required in assembly of Leishmania flagellum. J. Cell Sci. 2010, 123, 2035–2044. [Google Scholar] [CrossRef] [Green Version]
- Yasuzawa, T.; Iida, T.; Yoshida, M.; Hirayama, N.; Takahashi, M.; Shirahata, K.; Sano, H. The structures of the novel protein kinase C inhibitors K252a, b, c and d. J. Antibiot. 1986, 39, 1072–1078. [Google Scholar] [CrossRef]
- Elliott, L.H.; Wilkinson, S.E.; Sedgwick, A.D.; Hill, C.H.; Lawton, G.; Davis, P.D.; Nixon, J.S. K252a is a potent and selective inhibitor of phosphorylase kinase. Biochem. Biophys. Res. Commun. 1990, 171, 148–154. [Google Scholar] [CrossRef]
- Davis, P.D.; Hill, C.H.; Lawton, G.; Nixon, J.S.; Wilkinson, S.E.; Hutst, S.A.; Keech, E.; Turner, S.E. Inhibitors of protein kinase C. 1. 2,3-Bisarylmaleimides. J. Med. Chem. 1992, 35, 177–184. [Google Scholar] [CrossRef]
- Teshima, R.; Saito, Y.; Ikebuchi, H.; Rajiva De Silva, N.; Morita, Y.; Nakanishi, M.; Sawada, J.; Kitani, S. Effect of an ectokinase inhibitor, K252b, on degranulation and Ca2+ signals of RBL-2H3 cells and human basophils. J. Immunol. 1997, 159, 964–969. [Google Scholar]
- Lautrette, C.; Loum-Ribot, E.; Petit, D.; Vermot-Desroches, C.; Wijdenes, J.; Jauberteau, M.O. Increase of Fas-induced apoptosis by inhibition of extracellular phosphorylation of Fas receptor in Jurkat cell line. Apoptosis 2006, 11, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, K.; Nakanishi, S.; Matsuda, Y. Inhibition of nerve growth factor-induced neurite outgrowth of PC12 cells by a protein kinase inhibitor which does not permeate the cell membrane. FEBS Lett. 1991, 293, 119–123. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | L. amazonensis IC50 (µM) | L. donovani IC50 (µM) | T. cruzi IC50 (µM) |
|---|---|---|---|
| 1 STS | 0.08 ± 0.02 | 2.07 ± 0.14 | 3.63 ± 0.77 |
| 2 7OSTS | 3.58 ± 1.10 | 0.56 ± 0.06 | 1.58 ± 0.52 |
| 3 4′D4′OSTS | 17.10 ± 4.78 | > 40 | 17.10 ± 1.64 |
| 4 SCZ B | 10.44 ± 0.21 | > 40 | 12.50 ± 2.06 |
| 5 Rebeccamycin | > 40 | > 40 | > 40 |
| 6 K252a | 5.90 ± 0.96 | 8.09 ± 1.12 | 4.00 ± 0.24 |
| 7 K252b | 20.62 ± 4.50 | 4.45 ± 0.71 | 7.41 ± 0.93 |
| 8 K252c | > 40 | > 40 | > 40 |
| 9 Arcyriaflavin A | > 40 | > 40 | > 40 |
| Miltefosine * | 6.48 ± 0.24 | 3.32 ± 0.27 | - |
| Benznidazole * | - | - | 6.94 ± 1.94 |
| Compounds | Macrophage J774A.1 CC50 (µM) |
|---|---|
| 1 STS | 8.74 ± 0.72 |
| 2 7OSTS | 5.20 ± 1.75 |
| 3 4′D4′OSTS | > 40 |
| 4 SCZ B | > 40 |
| 5 Rebeccamycin | 1.42 ± 0.19 |
| 6 K252a | 1.07 ± 0.21 |
| 7 K252b | > 40 |
| 8 K252c | 35.4 ± 2.47 |
| 9 Arcyriaflavin A | > 40 |
| Miltefosine * | 72.19 ± 3.06 |
| Benznidazole * | 400.00 ± 4.00 |
| Compounds | L. Amazonensis Amastigotes IC50 (µM) | Selectivity Index (CC50/IC50) |
|---|---|---|
| 1 STS | 10 * | -- |
| 2 7OSTS | 0.10 ± 0.00 | 52 |
| 3 4′D4′OSTS | 2.03 ± 0.27 | 20 |
| 4 SCZ B | 2.47 ± 0.09 | 16 |
| Miltefosine ** | 3.12 ± 0,30 | 23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cartuche, L.; Sifaoui, I.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Lorenzo-Morales, J.; Piñero, J.E.; Díaz-Marrero, A.R.; Fernández, J.J. Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis. Biomolecules 2020, 10, 657. https://doi.org/10.3390/biom10040657
Cartuche L, Sifaoui I, López-Arencibia A, Bethencourt-Estrella CJ, San Nicolás-Hernández D, Lorenzo-Morales J, Piñero JE, Díaz-Marrero AR, Fernández JJ. Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis. Biomolecules. 2020; 10(4):657. https://doi.org/10.3390/biom10040657
Chicago/Turabian StyleCartuche, Luis, Ines Sifaoui, Atteneri López-Arencibia, Carlos J. Bethencourt-Estrella, Desirée San Nicolás-Hernández, Jacob Lorenzo-Morales, José E. Piñero, Ana R. Díaz-Marrero, and José J. Fernández. 2020. "Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis" Biomolecules 10, no. 4: 657. https://doi.org/10.3390/biom10040657
APA StyleCartuche, L., Sifaoui, I., López-Arencibia, A., Bethencourt-Estrella, C. J., San Nicolás-Hernández, D., Lorenzo-Morales, J., Piñero, J. E., Díaz-Marrero, A. R., & Fernández, J. J. (2020). Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis. Biomolecules, 10(4), 657. https://doi.org/10.3390/biom10040657