Living in a Foster Home: The Single Subpolar Flagellum Fla1 of Rhodobacter sphaeroides


Abstract
:1. Introduction
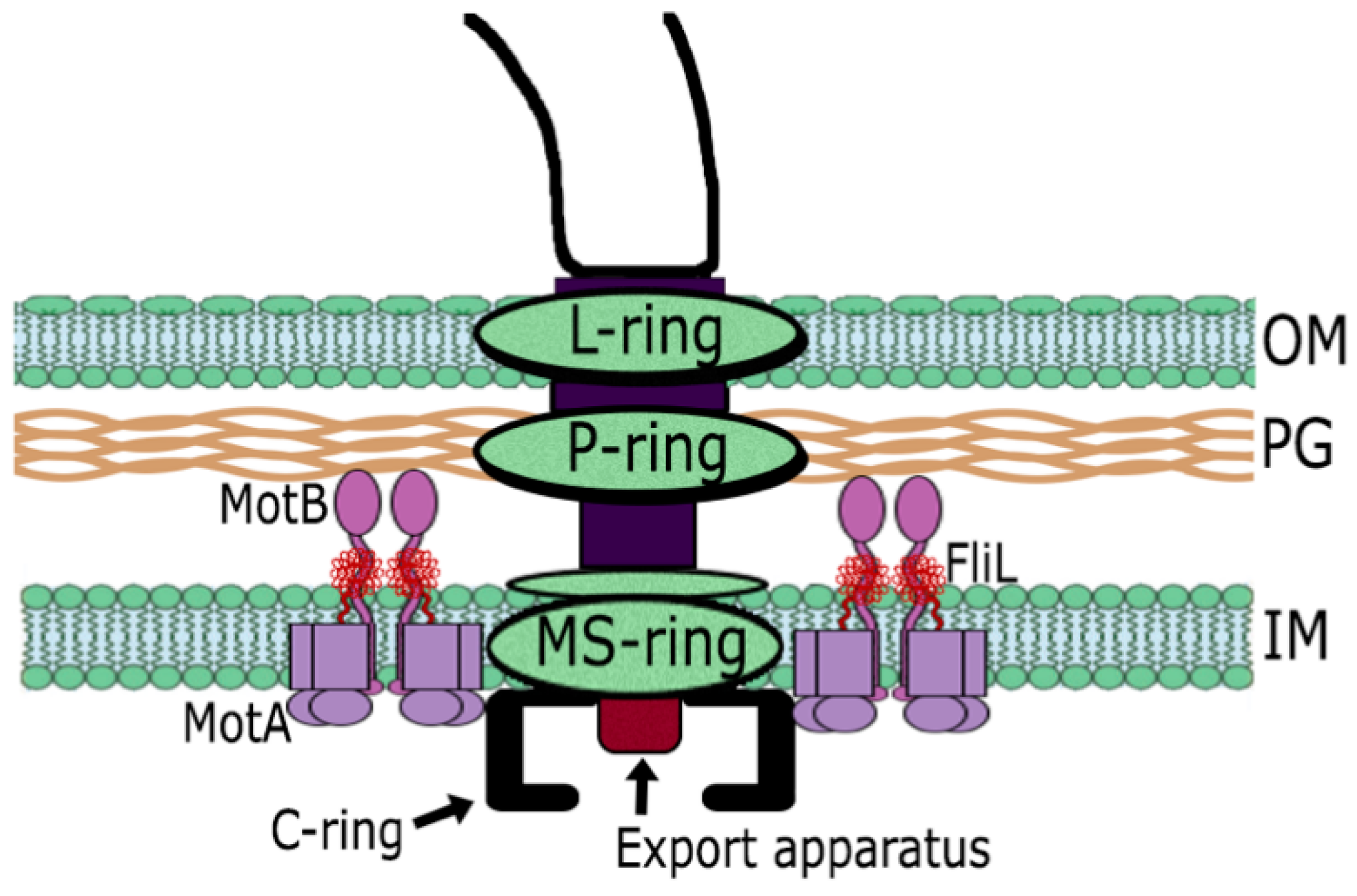
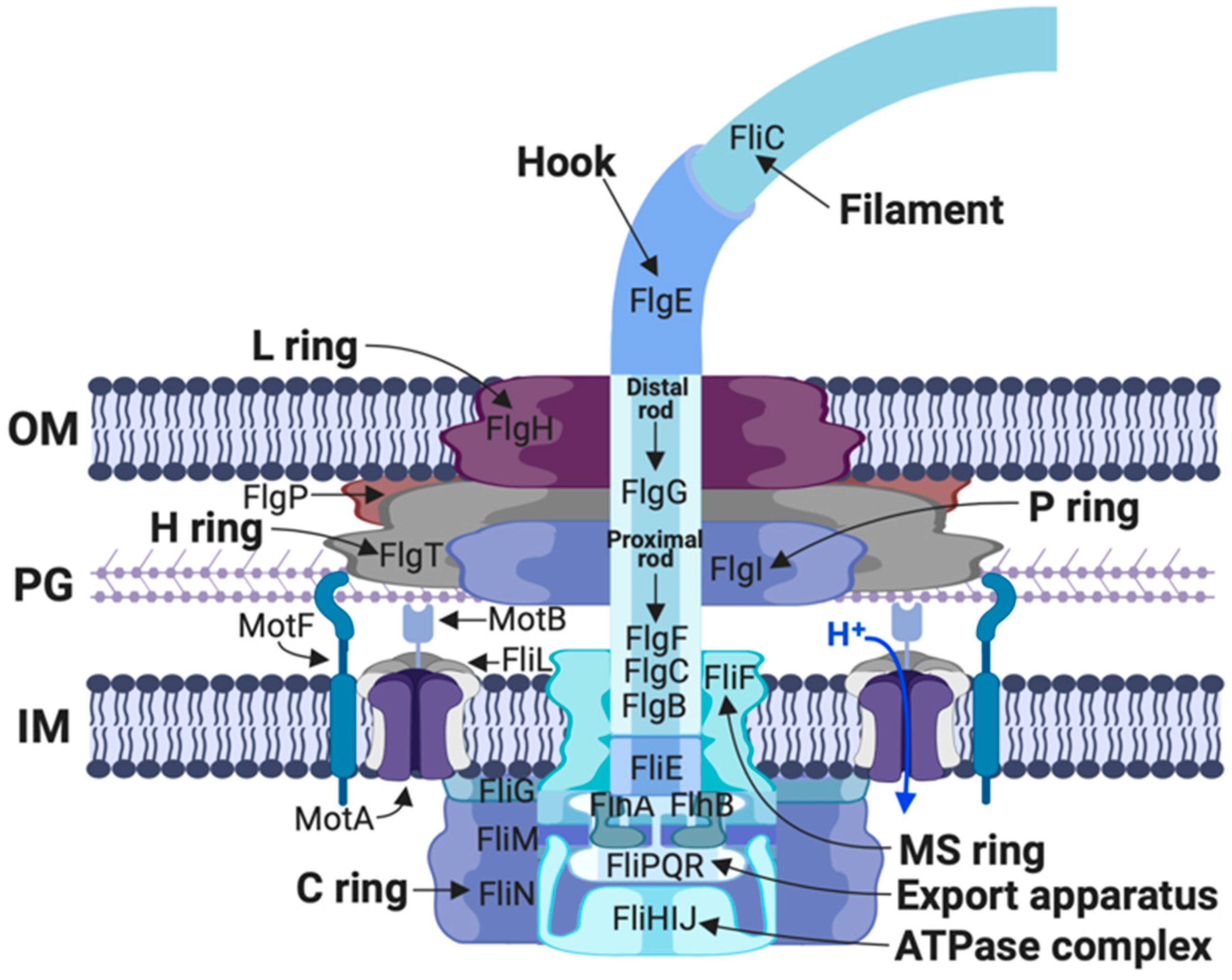
1.1. The Flagellar Structure
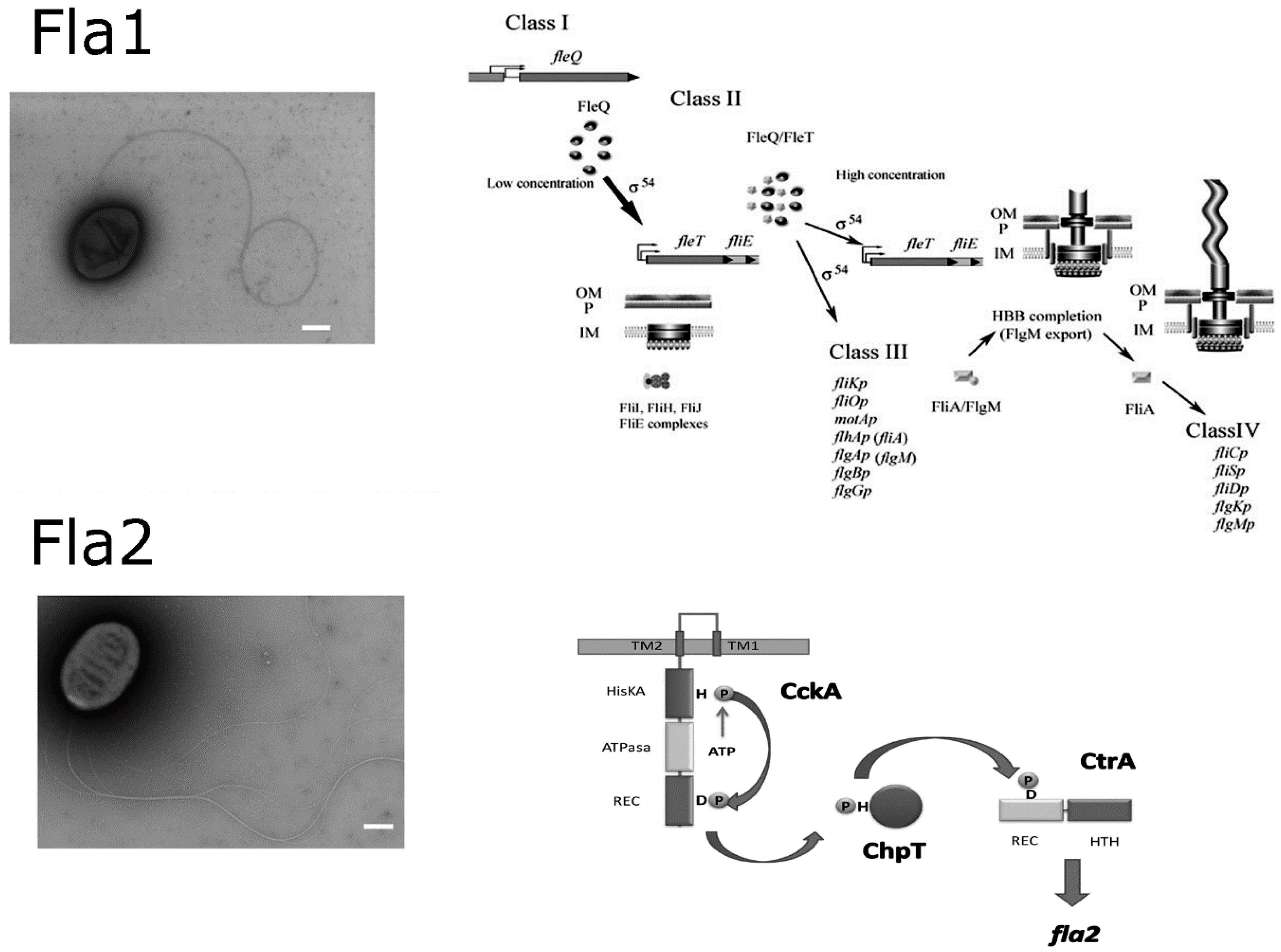
1.2. The Two Flagellar Systems of Rhodobacter sphaeroides
2. Overview of the Flagellar Genetic System in R. sphaeroides
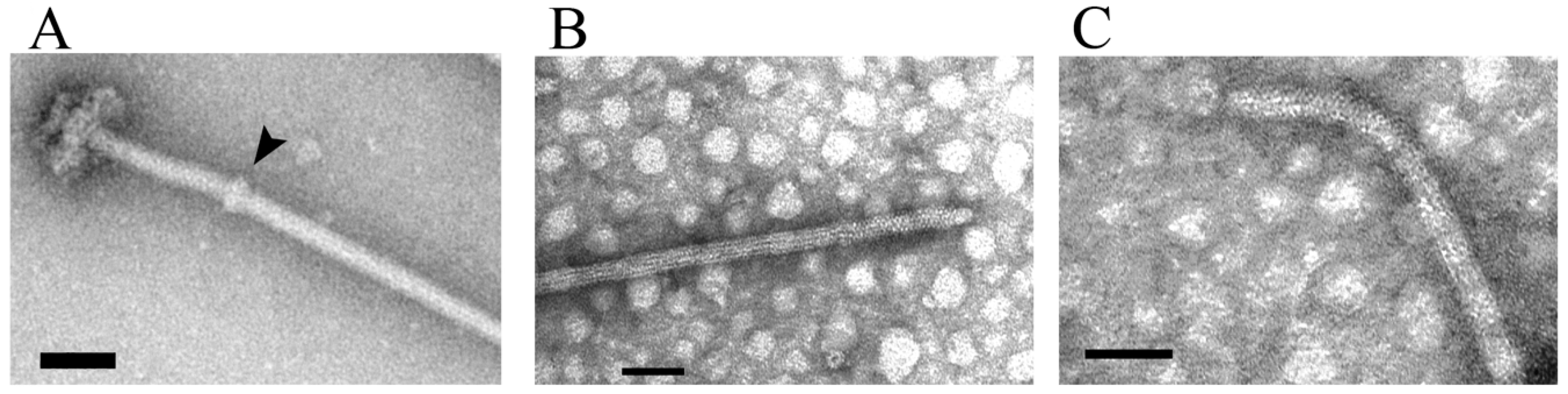
3. The Hook and Basal Body
4. Rod Assembly and Opening of the Peptidoglycan Barrier
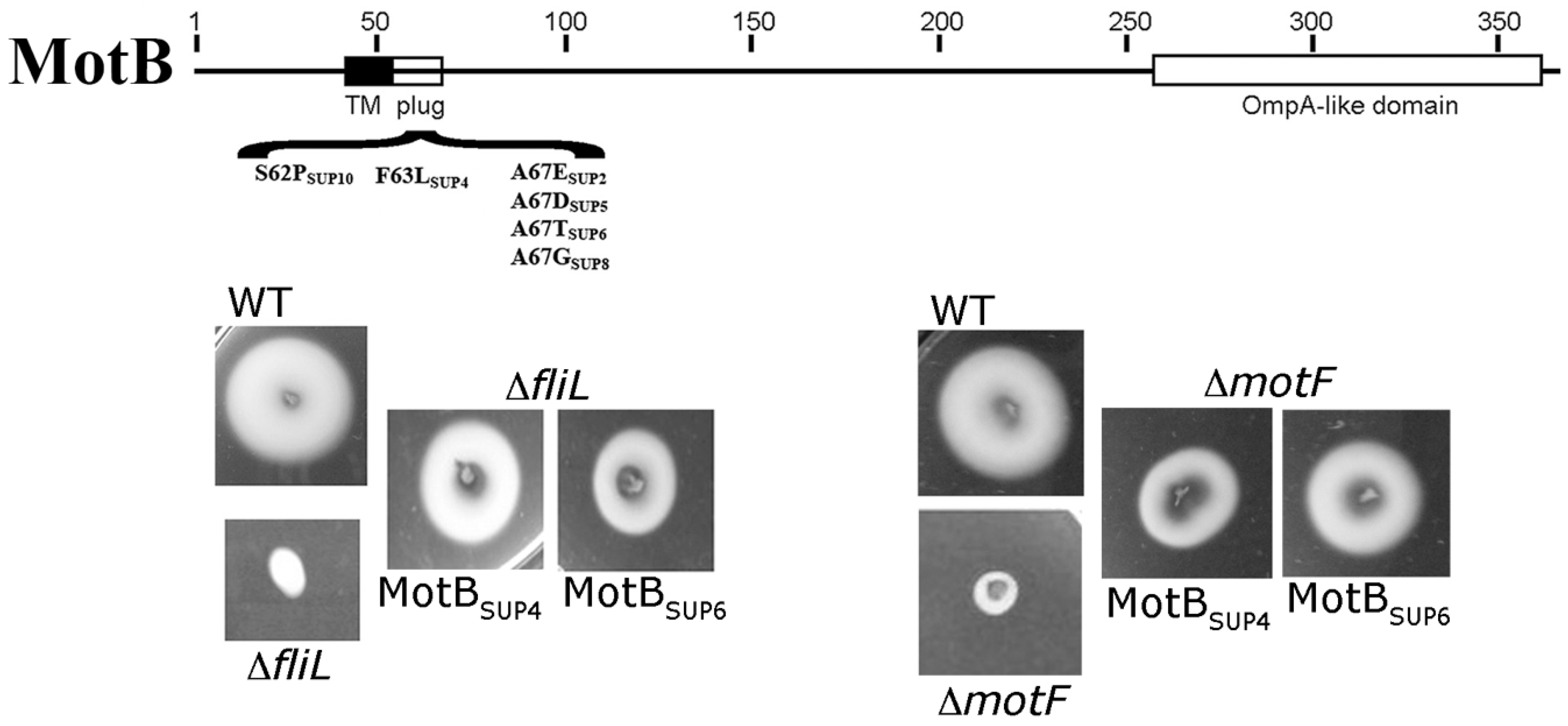
5. The Flagellar Motor of R. sphaeroides
6. Dominance of Fla1 over Fla2
7. Future Directions
Funding
Acknowledgments
Conflicts of Interest
References
- Nakamura, S.; Minamino, T. Flagella-Driven Motility of Bacteria. Biomolecules 2019, 9, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macnab, R.M. Type III flagellar protein export and flagellar assembly. Biochim. Biophys. Acta (BBA) Bioenerg. 2004, 1694, 207–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhlen, L.; Abrusci, P.; Johnson, S.; Gault, J.; Deme, J.; Caesar, J.; Dietsche, T.; Mebrhatu, M.T.; Ganief, T.; Macek, B.; et al. Structure of the Core of the Type Three Secretion System Export Apparatus. Nat. Struct. Mol. Biol. 2018, 25, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, T.; Makino, F.; Dietsche, T.; Kinoshita, M.; Kato, T.; Wagner, S.; Namba, K.; Imada, K.; Minamino, T. Assembly and stoichiometry of the core structure of the bacterial flagellar type III export gate complex. PLoS Biol. 2017, 15, e2002281. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Norris, S.J.; Liu, J. Molecular Architecture of the Bacterial Flagellar Motor in Cells. Biochemistry 2014, 53, 4323–4333. [Google Scholar] [CrossRef] [Green Version]
- Homma, M.; DeRosier, D.J.; Macnab, R.M. Flagellar hook and hook-associated proteins of Salmonella typhimurium and their relationship to other axial components of the flagellum. J. Mol. Biol. 1990, 213, 819–832. [Google Scholar] [CrossRef]
- Jones, C.J.; Homma, M.; Macnab, R.M. Identification of proteins of the outer (L and P) rings of the flagellar basal body of Escherichia coli. J. Bacteriol. 1987, 169, 1489–1492. [Google Scholar] [CrossRef] [Green Version]
- Akiba, T.; Yoshimura, H.; Namba, K. Monolayer crystallization of flagellar L-P rings by sequential addition and depletion of lipid. Science 1991, 252, 1544–1546. [Google Scholar] [CrossRef]
- Hirano, T.; Minamino, T.; Macnab, R.M. The role in flagellar rod assembly of the N-terminal domain of Salmonella FlgJ, a flagellum-specific muramidase. J. Mol. Biol. 2001, 312, 359–369. [Google Scholar] [CrossRef]
- Herlihey, F.A.; Moynihan, P.; Clarke, A.J. The Essential Protein for Bacterial Flagella Formation FlgJ Functions as a β-N-Acetylglucosaminidase. J. Biol. Chem. 2014, 289, 31029–31042. [Google Scholar] [CrossRef] [Green Version]
- Zaloba, P.; Bailey-Elkin, B.A.; Derksen, M.; Mark, B.L. Structural and Biochemical Insights into the Peptidoglycan Hydrolase Domain of FlgJ from Salmonella typhimurium. PLoS ONE 2016, 11, e0149204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nambu, T.; Minamino, T.; Macnab, R.M.; Kutsukake, K. Peptidoglycan-Hydrolyzing Activity of the FlgJ Protein, Essential for Flagellar Rod Formation in Salmonella typhimurium. J. Bacteriol. 1999, 181, 1555–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsunami, H.; Barker, C.S.; Yoon, Y.-H.; Wolf, M.; Samatey, F. Complete structure of the bacterial flagellar hook reveals extensive set of stabilizing interactions. Nat. Commun. 2016, 7, 13425. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Matsunami, H.; Aizawa, S.-I.; Wolf, M. Torque transmission mechanism of the curved bacterial flagellar hook revealed by cryo-EM. Nat. Struct. Mol. Biol. 2019, 26, 941–945. [Google Scholar] [CrossRef]
- Samatey, F.; Matsunami, H.; Imada, K.; Nagashima, S.; Shaikh, T.R.; Thomas, D.R.; Chen, J.Z.; DeRosier, D.J.; Kitao, A.; Namba, K. Structure of the bacterial flagellar hook and implication for the molecular universal joint mechanism. Nature 2004, 431, 1062–1068. [Google Scholar] [CrossRef]
- Kato, T.; Makino, F.; Miyata, T.; Horváth, P.; Namba, K. Structure of the native supercoiled flagellar hook as a universal joint. Nat. Commun. 2019, 10, 5295–5298. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.J.; Kim, T.H.; Song, W.S.; Ko, H.-J.; Lee, G.-S.; Kang, S.G.; Kim, P.-H.; Yoon, S.-I. Crystal structure of FlgL and its implications for flagellar assembly. Sci. Rep. 2018, 8, 14307. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Imada, K. The bacterial flagellar motor and its structural diversity. Trends Microbiol. 2015, 23, 267–274. [Google Scholar] [CrossRef]
- Minamino, T.; Terahara, N.; Kojima, S.; Namba, K. Autonomous control mechanism of stator assembly in the bacterial flagellar motor in response to changes in the environment. Mol. Microbiol. 2018, 109, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Gonzalez-Bonet, G.; Bilwes, A.M.; Crane, B.R.; Blair, D.F. Architecture of the flagellar rotor. EMBO J. 2011, 30, 2962–2971. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, Y.V.; Minamino, T. Structure and Function of the Bi-Directional Bacterial Flagellar Motor. Biomolecules 2014, 4, 217–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Lloyd, S.A.; Blair, D.F. Electrostatic interactions between rotor and stator in the bacterial flagellar motor. Proc. Natl. Acad. Sci. USA 1998, 95, 6436–6441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, Y.V.; Nakamura, S.; Kami-Ike, N.; Namba, K.; Minamino, T. Charged residues in the cytoplasmic loop of MotA are required for stator assembly into the bacterial flagellar motor. Mol. Microbiol. 2010, 78, 1117–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, Y.V.; Nakamura, S.; Hiraoka, K.D.; Namba, K.; Minamino, T. Distinct Roles of Highly Conserved Charged Residues at the MotA-FliG Interface in Bacterial Flagellar Motor Rotation. J. Bacteriol. 2012, 195, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Terashima, H.; Fukuoka, H.; Yakushi, T.; Kojima, S.; Homma, M. The Vibrio motor proteins, MotX and MotY, are associated with the basal body of Na+-driven flagella and required for stator formation. Mol. Microbiol. 2006, 62, 1170–1180. [Google Scholar] [CrossRef]
- Zhu, S.; Nishikino, T.; Takekawa, N.; Terashima, H.; Kojima, S.; Imada, K.; Homma, M.; Liu, J. In Situ Structure of the Vibrio Polar Flagellum Reveals a Distinct Outer Membrane Complex and Its Specific Interaction with the Stator. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef]
- Kojima, S.; Takao, M.; Almira, G.; Kawahara, I.; Sakuma, M.; Homma, M.; Kojima, C.; Imada, K. The Helix Rearrangement in the Periplasmic Domain of the Flagellar Stator B Subunit Activates Peptidoglycan Binding and Ion Influx. Structure 2018, 26, 590–598.e5. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Imada, K.; Sakuma, M.; Sudo, Y.; Kojima, C.; Minamino, T.; Homma, M.; Namba, K. Stator assembly and activation mechanism of the flagellar motor by the periplasmic region of MotB. Mol. Microbiol. 2009, 73, 710–718. [Google Scholar] [CrossRef]
- Zhu, S.; Takao, M.; Li, N.; Sakuma, M.; Nishino, Y.; Homma, M.; Kojima, S.; Imada, K. Conformational change in the periplamic region of the flagellar stator coupled with the assembly around the rotor. Proc. Natl. Acad. Sci. USA 2014, 111, 13523–13528. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J.; Xie, M.; Hijnen, M.; Roujeinikova, A. Role of the MotB linker in the assembly and activation of the bacterial flagellar motor. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 1009–1016. [Google Scholar] [CrossRef]
- Reboul, C.F.; Andrews, D.A.; Nahar, M.F.; Buckle, A.M.; Roujeinikova, A. Crystallographic and Molecular Dynamics Analysis of Loop Motions Unmasking the Peptidoglycan-Binding Site in Stator Protein MotB of Flagellar Motor. PLoS ONE 2011, 6, e18981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, D.A.; Nesmelov, Y.E.; Wilce, J.A.; Roujeinikova, A. Structural analysis of variant of Helicobacter pylori MotB in its activated form, engineered as chimera of MotB and leucine zipper. Sci. Rep. 2017, 7, 13435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, J.D.; Nieto, V.; Harshey, R.M. A New Player at the Flagellar Motor: FliL Controls both Motor Output and Bias. mBio 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suaste-Olmos, F.; Domenzain, C.; Mireles-Rodríguez, J.C.; Poggio, S.; Osorio, A.; Dreyfus, G.; Camarena, L. The Flagellar Protein FliL Is Essential for Swimming in Rhodobacter sphaeroides. J. Bacteriol. 2010, 192, 6230–6239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Kumar, A.; Kojima, S.; Homma, M. FliL associates with the stator to support torque generation of the sodium-driven polar flagellar motor of Vibrio. Mol. Microbiol. 2015, 98, 101–110. [Google Scholar] [CrossRef]
- Subramanian, S.; Kearns, D.B. Functional Regulators of Bacterial Flagella. Annu. Rev. Microbiol. 2019, 73, 225–246. [Google Scholar] [CrossRef]
- Takekawa, N.; Isumi, M.; Terashima, H.; Zhu, S.; Nishino, Y.; Sakuma, M.; Kojima, S.; Homma, M.; Imada, K. Structure of Vibrio FliL, a New Stomatin-like Protein That Assists the Bacterial Flagellar Motor Function. mBio 2019, 10, e00292-19. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Howell, J.K.; Bradley, S.D.; Zheng, Y.; Zhou, Z.H.; Norris, S.J. Cellular Architecture of Treponema pallidum: Novel Flagellum, Periplasmic Cone, and Cell Envelope as Revealed by Cryo Electron Tomography. J. Mol. Biol. 2010, 403, 546–561. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Beeby, M.E.; Murphy, G.; Leadbetter, J.R.; Hendrixson, D.R.; Briegel, A.; Li, Z.; Shi, J.I.; Tocheva, E.; Müller, A.; et al. Structural diversity of bacterial flagellar motors. EMBO J. 2011, 30, 2972–2981. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhang, K.; Boquoi, T.; Hu, B.; Motaleb, A.; Miller, K.A.; James, M.E.; Charon, N.W.; Manson, M.D.; Norris, S.J.; et al. Cryoelectron tomography reveals the sequential assembly of bacterial flagella in Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2013, 110, 14390–14395. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Nishikino, T.; Hu, B.; Kojima, S.; Homma, M.; Liu, J. Molecular architecture of the sheathed polar flagellum in Vibrio alginolyticus. Proc. Natl. Acad. Sci. USA 2017, 114, 10966–10971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, M.; Ghosal, D.; Subramanian, P.; Oikonomou, C.M.; Kjær, A.; Pirbadian, S.; Ortega, D.R.; Briegel, A.; El-Naggar, M.Y.; Jensen, G.J. The presence and absence of periplasmic rings in bacterial flagellar motors correlates with stator type. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.; Subramanian, P.; Ghosal, D.; Oikonomou, C.M.; Pirbadian, S.; Starwalt-Lee, R.; Mageswaran, S.K.; Ortega, D.R.A.; Gralnick, J.; El-Naggar, M.Y.; et al. In situ imaging of the bacterial flagellar motor disassembly and assembly processes. EMBO J. 2019, 38, e100957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Schniederberend, M.; Zhitnitsky, D.; Jain, R.; Galán, J.E.; Kazmierczak, B.I.; Liu, J. In Situ Structures of Polar and Lateral Flagella Revealed by Cryo-Electron Tomography. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhoff. The Phototrophic Alpha Proteobacteria, 3rd ed.; Dworkin, M.F.S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer-Verlag: New York, NY, USA, 2006; Volume 5. [Google Scholar]
- MacKenzie, C.; Choudhary, M.; Larimer, F.W.; Predki, P.F.; Stilwagen, S.; Armitage, J.P.; Barber, R.D.; Donohue, T.J.; Hosler, J.P.; Newman, J.E.; et al. The home stretch, a first analysis of the nearly completed genome of Rhodobacter sphaeroides 2.4.1. Photosynth. Res. 2001, 70, 19–41. [Google Scholar] [CrossRef] [Green Version]
- Porter, S.L.; Wilkinson, D.A.; Byles, E.D.; Wadhams, G.H.; Taylor, S.; Saunders, N.; Armitage, J.P. Genome Sequence of Rhodobacter sphaeroides Strain WS8N. J. Bacteriol. 2011, 193, 4027–4028. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.-K.; Kim, S.J.; Cha, S.H.; Oh, Y.-K.; Rhee, H.-J.; Kim, M.-S.; Lee, J.K. Complete Genome Sequence of Rhodobacter sphaeroides KD131. J. Bacteriol. 2008, 191, 1118–1119. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.W.; Lower, R.P.; Kim, N.K.; Young, J.P.W. Introducing the bacterial ‘chromid’: Not a chromosome, not a plasmid. Trends Microbiol. 2010, 18, 141–148. [Google Scholar] [CrossRef]
- DiCenzo, G.C.; Finan, T.M. The Divided Bacterial Genome: Structure, Function, and Evolution. Microbiol. Mol. Biol. Rev. 2017, 81, 3. [Google Scholar] [CrossRef] [Green Version]
- Armitage, J.P.; Macnab, R.M. Unidirectional, intermittent rotation of the flagellum of Rhodobacter sphaeroides. J. Bacteriol. 1987, 169, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Youle, M.; Rohwer, F.; Stacy, A.; Whiteley, M.; Steel, B.C.; Delalez, N.J.; Nord, A.; Berry, R.M.; Armitage, J.P.; Kamoun, S.; et al. The Microbial Olympics. Nat. Rev. Microbiol. 2012, 10, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packer, H.; Harrison, D.; Dixon, R.; Armitage, J. The effect of pH on the growth and motility of Rhodobacter sphaeroides WS8 and the nature of the driving force of the flagellar motor. Biochim. Biophys. Acta (BBA) Bioenerg. 1994, 1188, 101–107. [Google Scholar] [CrossRef]
- Pilizota, T.; Brown, M.T.; Leake, M.C.; Branch, R.W.; Berry, R.M.; Armitage, J.P. A molecular brake, not a clutch, stops the Rhodobacter sphaeroides flagellar motor. Proc. Natl. Acad. Sci. USA 2009, 106, 11582–11587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poggio, S.; Osorio, A.; Dreyfus, G.; Camarena, L. The flagellar hierarchy of Rhodobacter sphaeroides is controlled by the concerted action of two enhancer-binding proteins. Mol. Microbiol. 2005, 58, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Fabela, S.; Domenzain, C.; De La Mora, J.; Osorio, A.; Ramirez-Cabrera, V.; Poggio, S.; Dreyfus, G.; Camarena, L. A Distant Homologue of the FlgT Protein Interacts with MotB and FliL and Is Essential for Flagellar Rotation in Rhodobacter sphaeroides. J. Bacteriol. 2013, 195, 5285–5296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-González, C.; Domenzain, C.; Poggio, S.; Gonzalez-Halphen, D.; Dreyfus, G.; Camarena, L. Characterization of FlgP, an Essential Protein for Flagellar Assembly in Rhodobacter sphaeroides. J. Bacteriol. 2018, 201. [Google Scholar] [CrossRef] [Green Version]
- Arai, H.; Roh, J.H.; Kaplan, S. Transcriptome Dynamics during the Transition from Anaerobic Photosynthesis to Aerobic Respiration in Rhodobacter sphaeroides 2.4.1. J. Bacteriol. 2007, 190, 286–299. [Google Scholar] [CrossRef] [Green Version]
- Roh, J.H.; Smith, W.E.; Kaplan, S. Effects of Oxygen and Light Intensity on Transcriptome Expression in Rhodobacter sphaeroides 2.4.1. J. Biol. Chem. 2003, 279, 9146–9155. [Google Scholar] [CrossRef] [Green Version]
- Poggio, S.; Abreu-Goodger, C.; Fabela, S.; Osorio, A.; Dreyfus, G.; Vinuesa, P.; Camarena, L. A Complete Set of Flagellar Genes Acquired by Horizontal Transfer Coexists with the Endogenous Flagellar System in Rhodobacter sphaeroides. J. Bacteriol. 2007, 189, 3208–3216. [Google Scholar] [CrossRef] [Green Version]
- Vega-Baray, B.; Domenzain, C.; Rivera, A.; Alfaro-López, R.; Gómez-César, E.; Poggio, S.; Dreyfus, G.; Camarena, L. The Flagellar Set Fla2 in Rhodobacter sphaeroides Is Controlled by the CckA Pathway and Is Repressed by Organic Acids and the Expression of Fla1. J. Bacteriol. 2015, 197, 833–847. [Google Scholar] [CrossRef] [Green Version]
- De La Mora, J.; Uchida, K.; Del Campo, A.M.; Camarena, L.; Aizawa, S.-I.; Dreyfus, G. Structural Characterization of the Fla2 Flagellum of Rhodobacter sphaeroides. J. Bacteriol. 2015, 197, 2859–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, A.M.; Ballado, T.; De La Mora, J.; Poggio, S.; Camarena, L.; Dreyfus, G. Chemotactic Control of the Two Flagellar Systems of Rhodobacter sphaeroides Is Mediated by Different Sets of CheY and FliM Proteins. J. Bacteriol. 2007, 189, 8397–8401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campo, A.M.-D.; Ballado, T.; Camarena, L.; Dreyfus, G. In Rhodobacter sphaeroides, Chemotactic Operon 1 Regulates Rotation of the Flagellar System 2. J. Bacteriol. 2011, 193, 6781–6786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.; Porter, S.L.; Martin, A.C.; Hamblin, P.A.; Armitage, J.P. Fine tuning bacterial chemotaxis: Analysis of Rhodobacter sphaeroides behaviour under aerobic and anaerobic conditions by mutation of the major chemotaxis operons and cheY genes. EMBO J. 2000, 19, 4601–4613. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, S.I. The Flagellar World: Electron Microscopic Images of Bacterial flagella and Related Surface Structures from More Than 30 Species; Elsevier Inc.: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Poggio, S.; Aguilar, C.; Osorio, A.; Gonzalez-Pedrajo, B.; Dreyfus, G.; Camarena, L. Sigma(54) promoters control expression of genes encoding the hook and basal body complex in Rhodobacter sphaeroides. J. Bacteriol. 2000, 182, 5787–5792. [Google Scholar] [CrossRef] [Green Version]
- Poggio, S.; Osorio, A.; Dreyfus, G.; Camarena, L. The four different σ54 factors of Rhodobacter sphaeroides are not functionally interchangeable. Mol. Microbiol. 2002, 46, 75–85. [Google Scholar] [CrossRef]
- Domenzain, C.; Camarena, L.; Osorio, A.; Dreyfus, G.; Poggio, S. Evolutionary origin of the Rhodobacter sphaeroides specialized RpoN sigma factors. FEMS Microbiol. Lett. 2011, 327, 93–102. [Google Scholar] [CrossRef]
- Gao, F.; Danson, A.E.; Ye, F.; Jovanovic, M.; Buck, M.; Zhang, X. Bacterial Enhancer Binding Proteins—AAA+ Proteins in Transcription Activation. Biomolecules 2020, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Buck, M. A Perspective on the Enhancer Dependent Bacterial RNA Polymerase. Biomolecules 2015, 5, 1012–1019. [Google Scholar] [CrossRef] [Green Version]
- Peña-Sánchez, J.; Poggio, S.; Flores-Pérez, Ú.; Osorio, A.; Domenzain, C.; Dreyfus, G.; Camarena, L. Identification of the binding site of the σ 54 hetero-oligomeric FleQ/FleT activator in the flagellar promoters of Rhodobacter sphaeroides. Microbiology 2009, 155, 1669–1679. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Valle, J.; Domenzain, C.; De La Mora, J.; Poggio, S.; Dreyfus, G.; Camarena, L. The Master Regulators of the Fla1 and Fla2 Flagella of Rhodobacter sphaeroides Control the Expression of Their Cognate CheY Proteins. J. Bacteriol. 2016, 199, e00670-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, S.L.; Wadhams, G.H.; Armitage, J.P. Signal processing in complex chemotaxis pathways. Nat. Rev. Microbiol. 2011, 9, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.C.; Gould, M.; Byles, E.; Roberts, M.; Armitage, J.P. Two Chemosensory Operons of Rhodobacter sphaeroides Are Regulated Independently by Sigma 28 and Sigma 54. J. Bacteriol. 2006, 188, 7932–7940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Osorio, A.; Osorio, A.; Poggio, S.; Dreyfus, G.; Camarena, L. Architecture of divergent flagellar promoters controlled by CtrA in Rhodobacter sphaeroides. BMC Microbiol. 2018, 18, 129. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Valle, J.; Sanchez-Flores, A.; Poggio, S.; Dreyfus, G.; Camarena, L. The CtrA Regulon of Rhodobacter sphaeroides Favors Adaptation to a Particular Lifestyle. J. Bacteriol. 2020, 202, JB.00678-19. [Google Scholar] [CrossRef]
- Sockett, R.E.; Foster, J.C.A.; Armitage, J.P. Molecular Biology of the Rhodobacter sphaeroides Flagellum. Mol. Biol. Membr. Bound Complexes Phototrophic Bact. 1990, 53, 473–478. [Google Scholar] [CrossRef]
- González-Pedrajo, B.; Ballado, T.; Campos, A.; E Sockett, R.; Camarena, L.; Dreyfus, G. Structural and genetic analysis of a mutant of Rhodobacter sphaeroides WS8 deficient in hook length control. J. Bacteriol. 1997, 179, 6581–6588. [Google Scholar] [CrossRef] [Green Version]
- West, M.A.; Dreyfus, G. Isolation and Ultrastructural Study of the Flagellar Basal Body Complex from Rhodobacter sphaeroidesWS8 (Wild Type) and a Polyhook Mutant PG. Biochem. Biophys. Res. Commun. 1997, 238, 733–737. [Google Scholar] [CrossRef]
- Castillo, D.J.; Ballado, T.; Camarena, L.; Dreyfus, G. Functional analysis of a large non-conserved region of FlgK (HAP1) from Rhodobacter sphaeroides. Antonie Van Leeuwenhoek 2009, 95, 77–90. [Google Scholar] [CrossRef]
- Eddy, S.R. Multiple alignment using hidden Markov models. In Proceedings of the Third International Conference on Intelligent Systems for Molecular Biology, Cambridge, UK, 16–19 July 1995; Volume 3, pp. 114–120, ISBN 0-929280-83-0. [Google Scholar]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2014, 43, D257–D260. [Google Scholar] [CrossRef]
- Fahrner, K.A.; Block, S.M.; Krishnaswamy, S.; Parkinson, J.S.; Berg, H.C. A Mutant Hook-associated Protein (HAP3) Facilitates Torsionally Induced Transformations of the Flagellar Filament of Escherichia coli. J. Mol. Biol. 1994, 238, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Ballado, T.; Camarena, L.; González-Pedrajo, B.; Silva-Herzog, E.; Dreyfus, G. The Hook Gene (flgE) Is Expressed from the flgBCDEF Operon in Rhodobacter sphaeroides: Study of an flgE Mutant. J. Bacteriol. 2001, 183, 1680–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Kato, T.; Namba, K. Specific Arrangement of α-Helical Coiled Coils in the Core Domain of the Bacterial Flagellar Hook for the Universal Joint Function. Structure 2009, 17, 1485–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaikh, T.R.; Thomas, D.R.; Chen, J.Z.; Samatey, F.; Matsunami, H.; Imada, K.; Namba, K.; DeRosier, D.J. A partial atomic structure for the flagellar hook of Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 2005, 102, 1023–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horváth, P.; Kato, T.; Miyata, T.; Namba, K. Structure of Salmonella Flagellar Hook Reveals Intermolecular Domain Interactions for the Universal Joint Function. Biomolecules 2019, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, K.D.; Morimoto, Y.V.; Inoue, Y.; Fujii, T.; Miyata, T.; Makino, F.; Minamino, T.; Namba, K. Straight and rigid flagellar hook made by insertion of the FlgG specific sequence into FlgE. Sci. Rep. 2017, 7, 46723. [Google Scholar] [CrossRef] [Green Version]
- Osorio-Valeriano, M.; De La Mora, J.; Camarena, L.; Dreyfus, G. Biochemical Characterization of the Flagellar Rod Components of Rhodobacter sphaeroides: Properties and Interactions. J. Bacteriol. 2016, 198, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Burrage, A.M.; Vanderpool, E.; Kearns, D.B. Assembly Order of Flagellar Rod Subunits in Bacillus subtilis. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [Green Version]
- González-Pedrajo, B.; De La Mora, J.; Ballado, T.; Camarena, L.; Dreyfus, G. Characterization of the flgG operon of Rhodobacter sphaeroides WS8 and its role in flagellum biosynthesis. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2002, 1579, 55–63. [Google Scholar] [CrossRef]
- De La Mora, J.; Ballado, T.; González-Pedrajo, B.; Camarena, L.; Dreyfus, G. The Flagellar Muramidase from the Photosynthetic Bacterium Rhodobacter sphaeroides. J. Bacteriol. 2007, 189, 7998–8004. [Google Scholar] [CrossRef] [Green Version]
- De La Mora, J.; Osorio-Valeriano, M.; González-Pedrajo, B.; Ballado, T.; Camarena, L.; Dreyfus, G. The C Terminus of the Flagellar Muramidase SltF Modulates the Interaction with FlgJ in Rhodobacter sphaeroides. J. Bacteriol. 2012, 194, 4513–4520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herlihey, F.A.; Osorio-Valeriano, M.; Dreyfus, G.; Clarke, A.J. Modulation of the Lytic Activity of the Dedicated Autolysin for Flagellum Formation SltF by Flagellar Rod Proteins FlgB and FlgF. J. Bacteriol. 2016, 198, 1847–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armitage, J.P.; Pitta, T.P.; Vigeant, M.; Packer, H.L.; Ford, R.M. Transformations in Flagellar Structure of Rhodobacter sphaeroides and Possible Relationship to Changes in Swimming Speed. J. Bacteriol. 1999, 181, 4825–4833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terashima, H.; Koike, M.; Kojima, S.; Homma, M. The Flagellar Basal Body-Associated Protein FlgT Is Essential for a Novel Ring Structure in the Sodium-Driven Vibrio Motor. J. Bacteriol. 2010, 192, 5609–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, S.; Tomás, J.M. The FlgT Protein Is Involved in Aeromonas hydrophila Polar Flagella Stability and Not Affects Anchorage of Lateral Flagella. Front. Microbiol. 2016, 7, 7141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, D.E.; Urbach, J.M.; Mekalanos, J.J. A defined transposon mutant library and its use in identifying motility genes in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2008, 105, 8736–8741. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Nishikino, T.; Kojima, S.; Homma, M.; Liu, J. The Vibrio H-Ring Facilitates the Outer Membrane Penetration of the Polar Sheathed Flagellum. J. Bacteriol. 2018, 200, JB-00387. [Google Scholar] [CrossRef] [Green Version]
- Beeby, M.; Ribardo, D.A.; Brennan, C.A.; Ruby, E.G.; Jensen, G.J.; Hendrixson, D.R. Diverse high-torque bacterial flagellar motors assemble wider stator rings using a conserved protein scaffold. Proc. Natl. Acad. Sci. USA 2016, 113, E1917–E1926. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Cabrera, V.; Poggio, S.; Domenzain, C.; Osorio, A.; Dreyfus, G.; Camarena, L. A Novel Component of the Rhodobacter sphaeroides Fla1 Flagellum Is Essential for Motor Rotation. J. Bacteriol. 2012, 194, 6174–6183. [Google Scholar] [CrossRef] [Green Version]
- Hosking, E.R.; Vogt, C.; Bakker, E.P.; Manson, M.D. The Escherichia coli MotAB Proton Channel Unplugged. J. Mol. Biol. 2006, 364, 921–937. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camarena, L.; Dreyfus, G. Living in a Foster Home: The Single Subpolar Flagellum Fla1 of Rhodobacter sphaeroides. Biomolecules 2020, 10, 774. https://doi.org/10.3390/biom10050774
Camarena L, Dreyfus G. Living in a Foster Home: The Single Subpolar Flagellum Fla1 of Rhodobacter sphaeroides. Biomolecules. 2020; 10(5):774. https://doi.org/10.3390/biom10050774
Chicago/Turabian StyleCamarena, Laura, and Georges Dreyfus. 2020. "Living in a Foster Home: The Single Subpolar Flagellum Fla1 of Rhodobacter sphaeroides" Biomolecules 10, no. 5: 774. https://doi.org/10.3390/biom10050774
APA StyleCamarena, L., & Dreyfus, G. (2020). Living in a Foster Home: The Single Subpolar Flagellum Fla1 of Rhodobacter sphaeroides. Biomolecules, 10(5), 774. https://doi.org/10.3390/biom10050774