Effects of Chronic Cannabidiol Treatment in the Rat Chronic Unpredictable Mild Stress Model of Depression

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Reagents
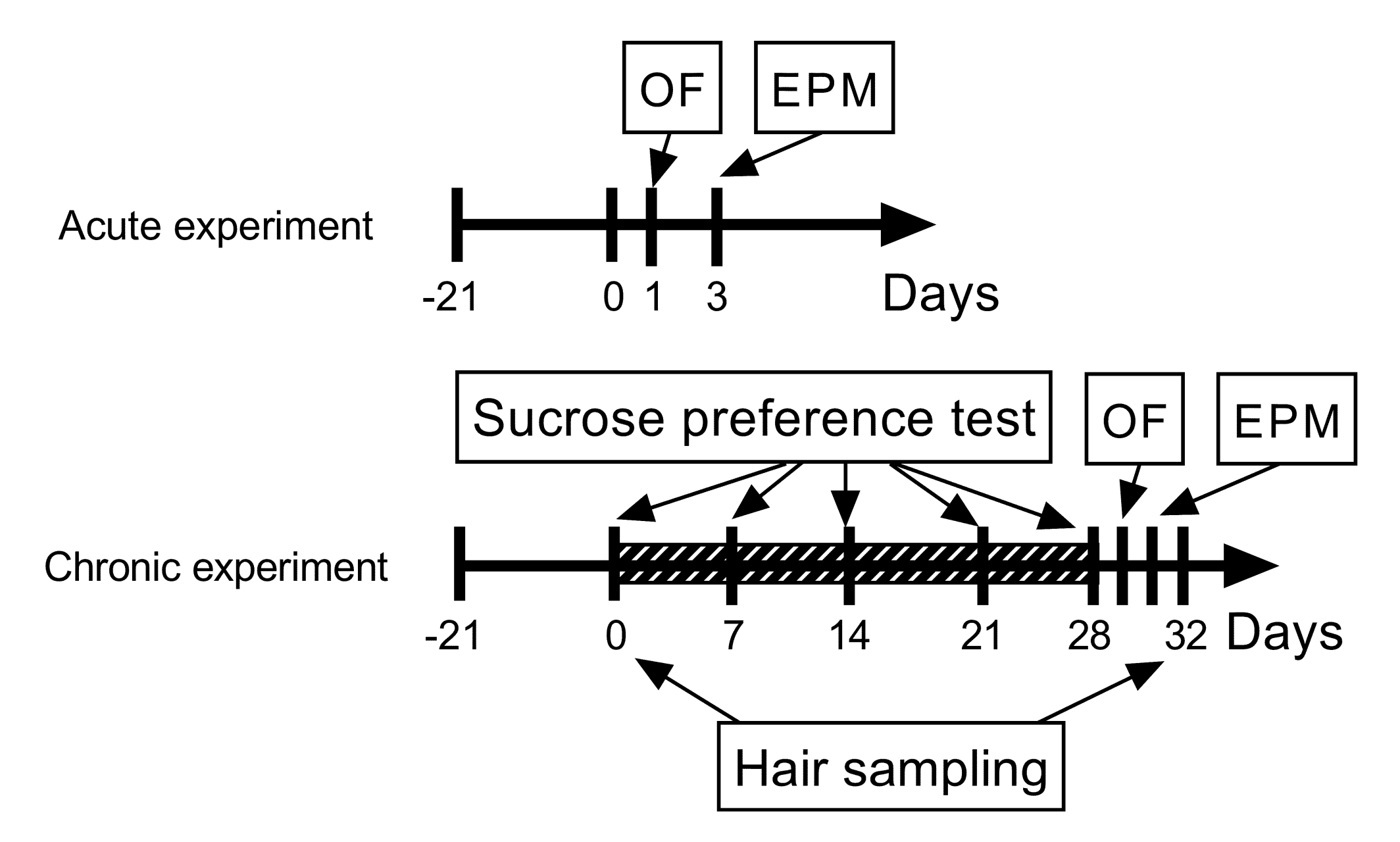
2.3. Chronic Unpredictable Mild Stress (CUMS)
2.4. Behavioral Assays
2.4.1. Sucrose Preference Test (SPT)
2.4.2. Open Field Test (OF)
2.4.3. Elevated Plus Maze Test (EPM)
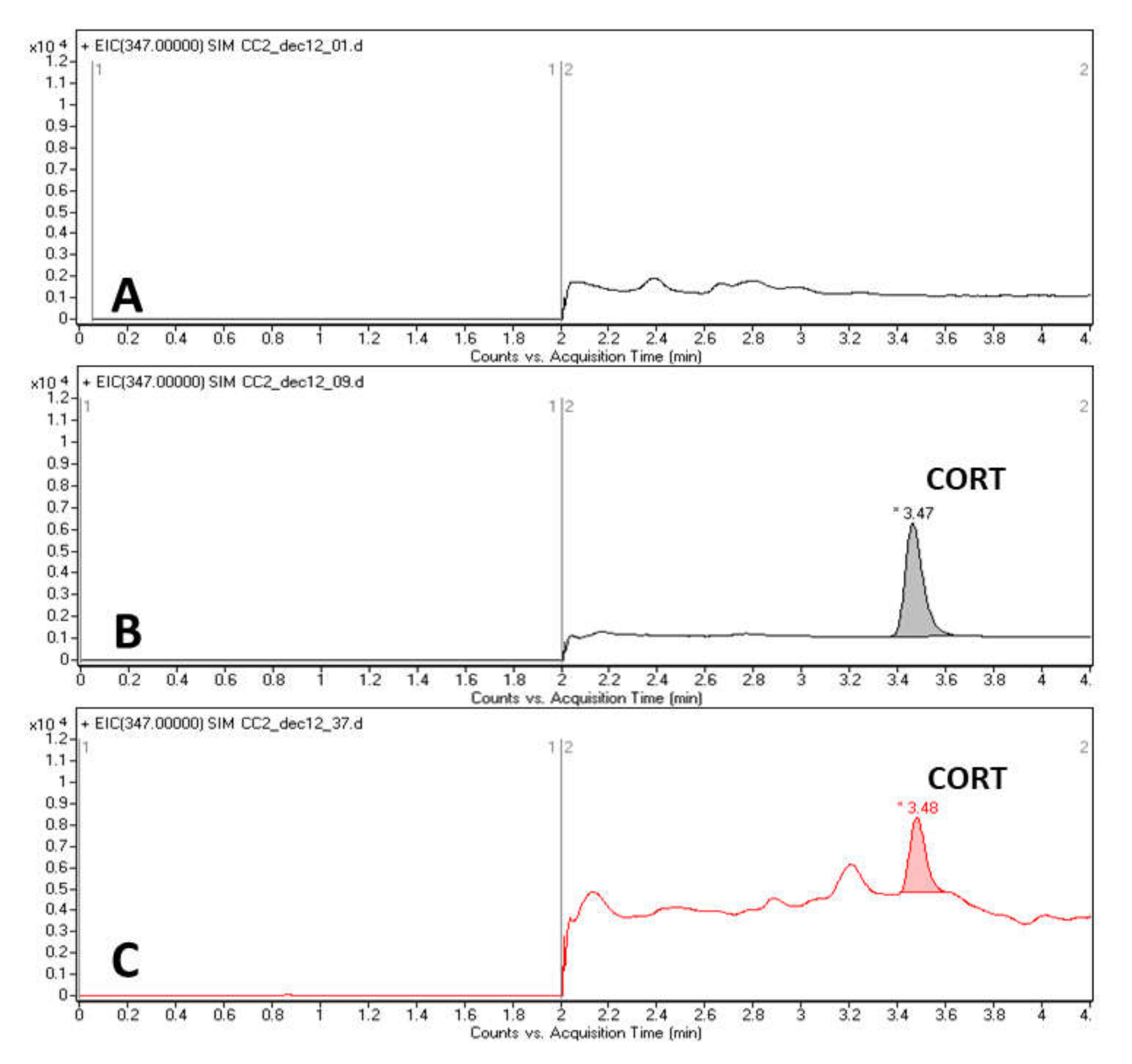
2.5. Hair Corticosterone Analysis
2.6. Statistical Analysis
3. Results
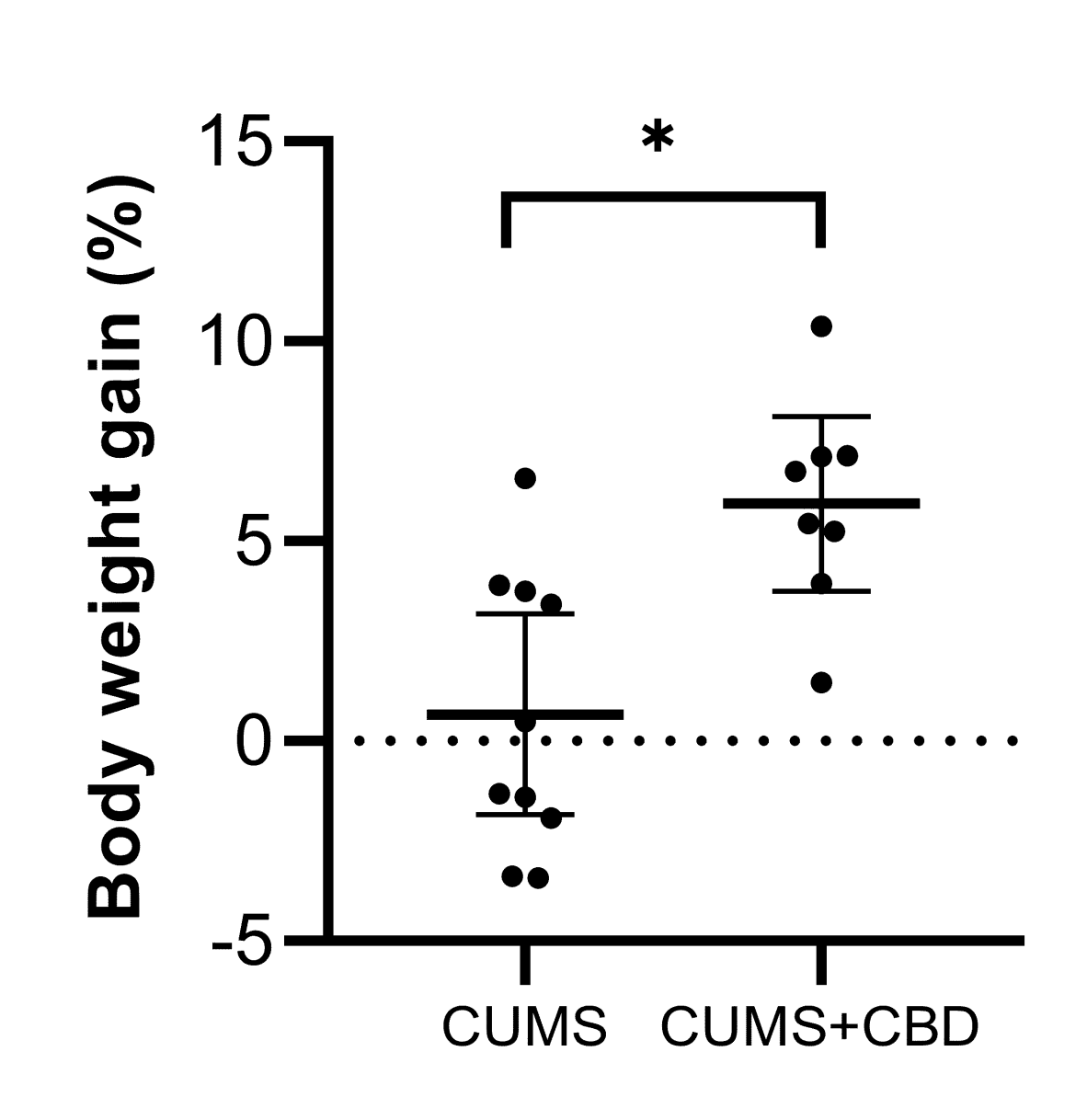
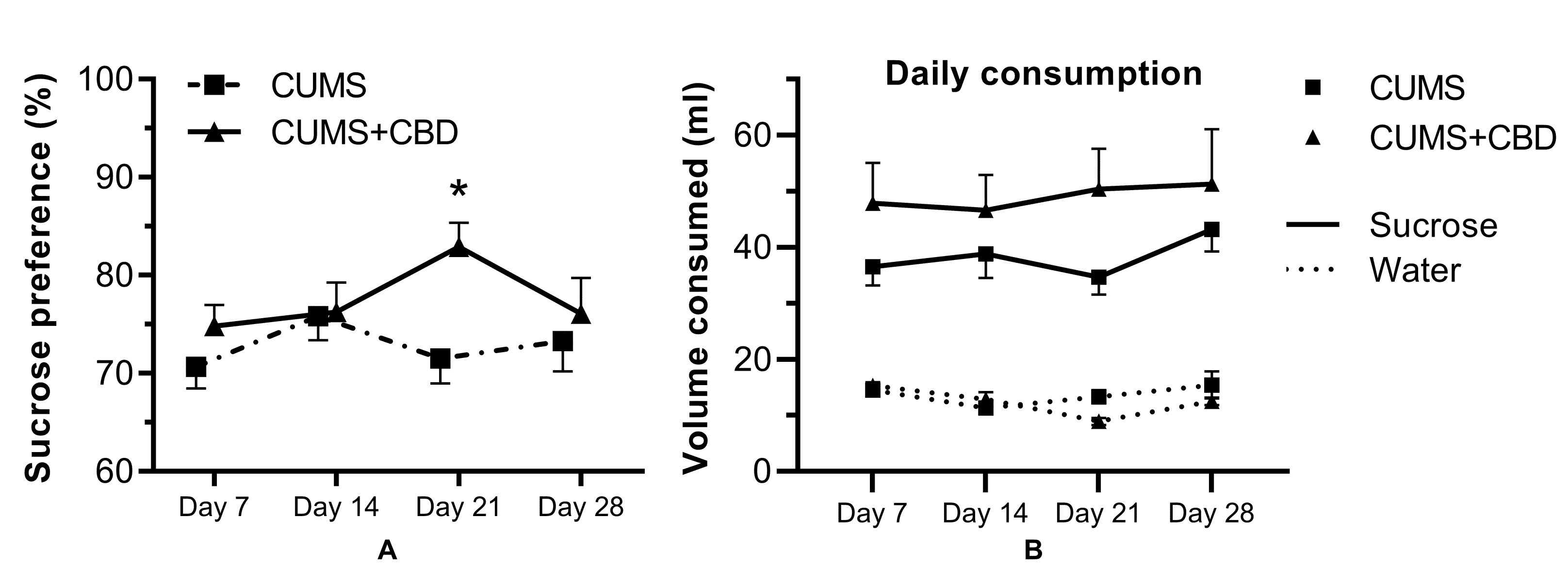
3.1. Sucrose Preference Test
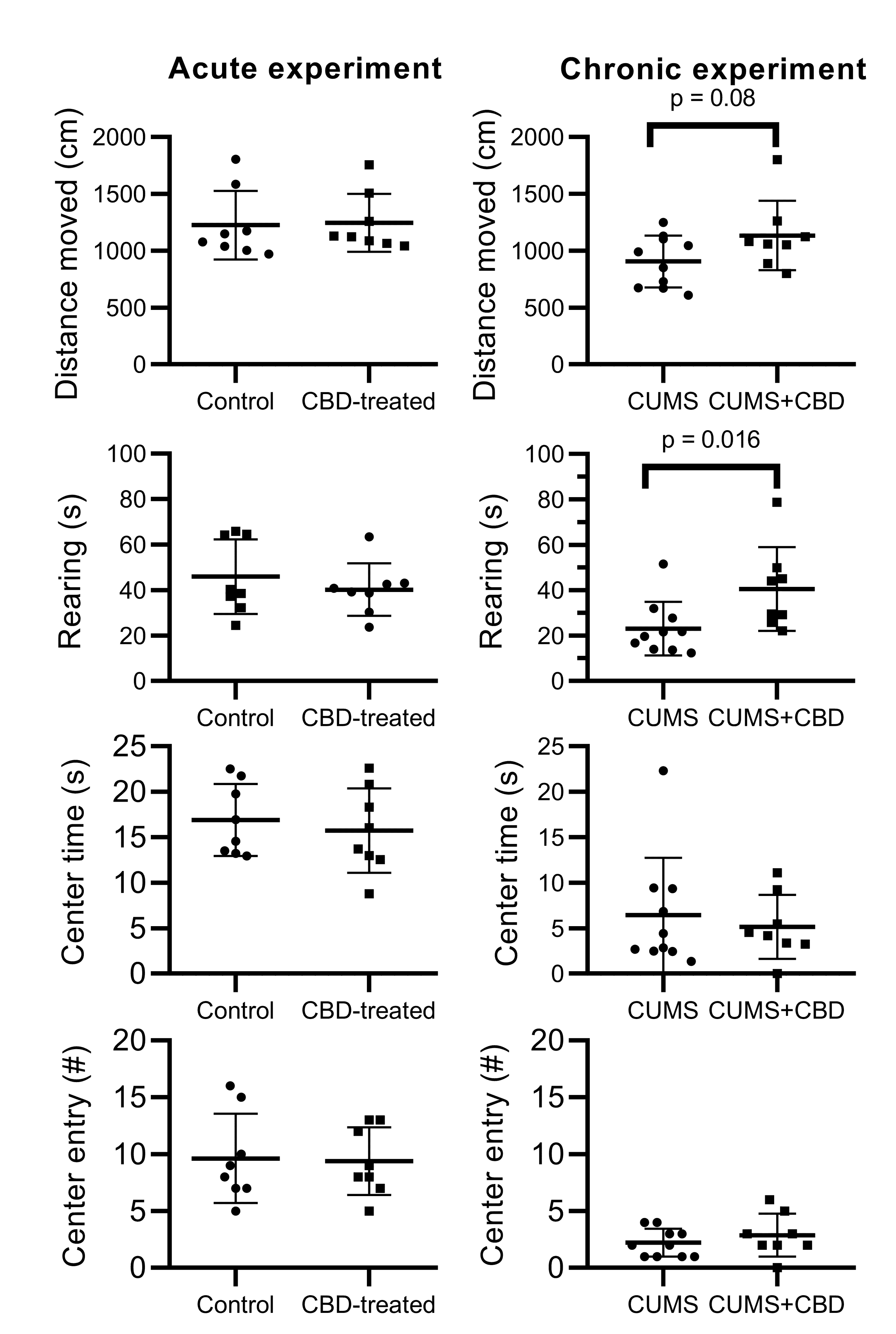
3.2. Open Field Test
3.3. Elevated Plus Maze Test
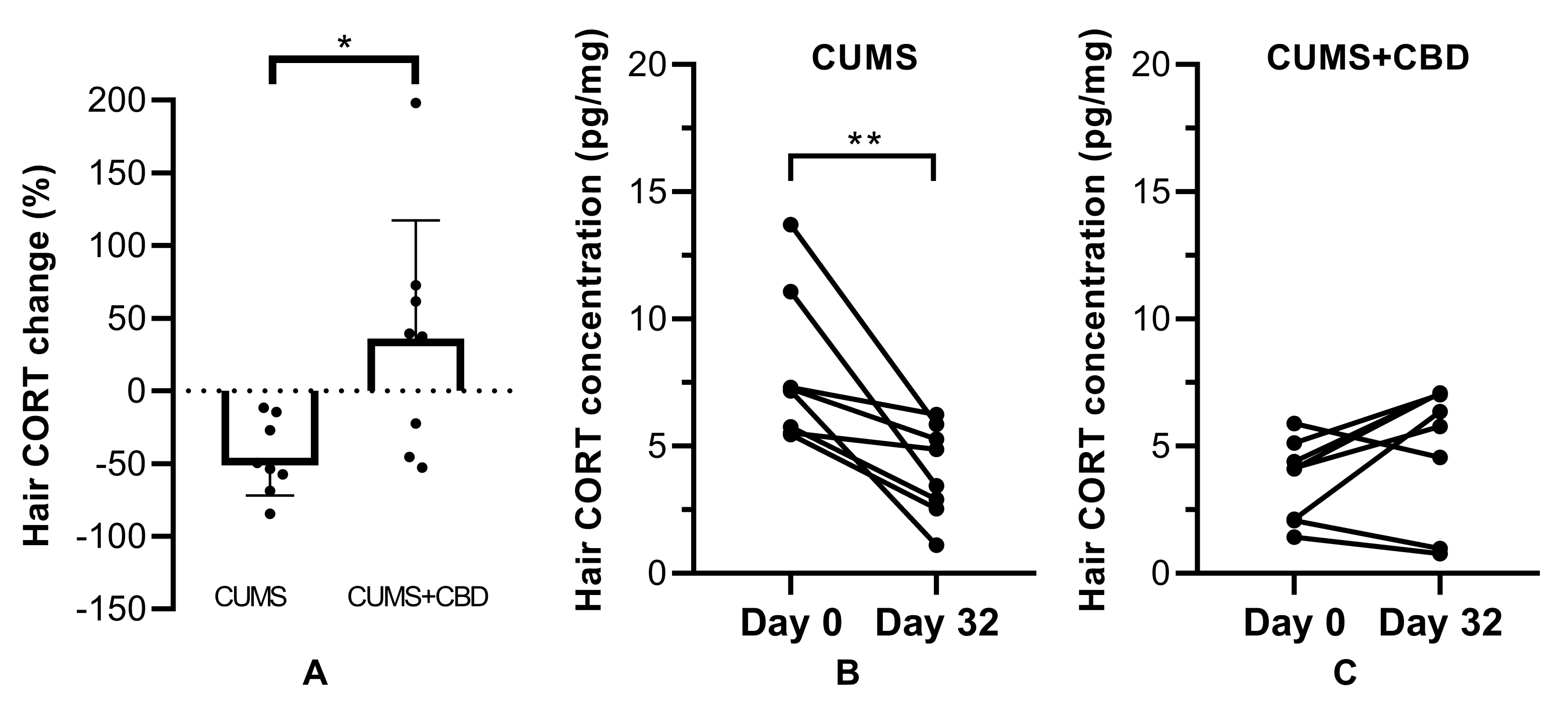
3.4. Hair Corticosterone
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Gambi, F.; De Berardis, D.; Sepede, G.; Quartesan, R.; Calcagni, E.; Salerno, R.; Conti, C.; Ferro, F. Cannabinoid Receptors and Their Relationships with Neuropsychiatric Disorders. Int. J. Immunopathol. Pharmacol. 2005, 18, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.W.; Borgelt, L.M.; Blackmer, A.B. Cannabidiol: A New Hope for Patients With Dravet or Lennox-Gastaut Syndromes. Ann. Pharmacother. 2019, 53, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Wise, J. European drug agency approves cannabis-based medicine for severe forms of epilepsy. BMJ 2019, 366, l5708. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, S.; Mammana, S.; Cavalli, E.; Bramanti, P.; Mazzon, E. Use of Cannabidiol in the Treatment of Epilepsy: Efficacy and Security in Clinical Trials. Molecules 2019, 24, 1459. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Trapero, M.; Pérez-Díaz, C.; Espejo-Porras, F.; De Lago, E.; Fernández-Ruiz, J. Pharmacokinetics of Sativex® in Dogs: Towards a Potential Cannabinoid-Based Therapy for Canine Disorders. Biomolecules 2020, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Turri, M.; Teatini, F.; Donato, F.; Zanette, G.; Tugnoli, V.; Deotto, L.; Bonetti, B.; Squintani, G. Pain Modulation after Oromucosal Cannabinoid Spray (SATIVEX®) in Patients with Multiple Sclerosis: A Study with Quantitative Sensory Testing and Laser-Evoked Potentials. Medicines 2018, 5, 59. [Google Scholar] [CrossRef] [Green Version]
- Campos, A.C.; Moreira, F.A.; Gomes, F.; Del Bel, E.; Guimarães, F.S. Multiple mechanisms involved in the large-spectrum therapeutic potential of cannabidiol in psychiatric disorders. Philos. Trans. R. Soc. B: Boil. Sci. 2012, 367, 3364–3378. [Google Scholar] [CrossRef]
- Crippa, J.A.S.; Derenusson, G.N.; Ferrari, T.B.; Wichert-Ana, L.; Souza-Duran, F.L.; Martin-Santos, R.; Simões, M.V.; Bhattacharyya, S.; Fusar-Poli, P.; Atakan, Z.; et al. Neural basis of anxiolytic effects of cannabidiol (CBD) in generalized social anxiety disorder: A preliminary report. J. Psychopharmacol. 2010, 25, 121–130. [Google Scholar] [CrossRef]
- Soares, V.P.; Campos, A.C. Evidences for the Anti-panic Actions of Cannabidiol. Curr. Neuropharmacol. 2017, 15, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Nardo, M.; Casarotto, P.C.; Gomes, F.; Guimaraes, F.S. Cannabidiol reverses the mCPP-induced increase in marble-burying behavior. Fundam. Clin. Pharmacol. 2013, 28, 544–550. [Google Scholar] [CrossRef]
- Bitencourt, R.M.; Takahashi, R.N. Cannabidiol as a Therapeutic Alternative for Post-traumatic Stress Disorder: From Bench Research to Confirmation in Human Trials. Front. Mol. Neurosci. 2018, 12, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherma, M.; Masia, P.; Deidda, M.; Fratta, W.; Tanda, G.; Fadda, P. New Perspectives on the Use of Cannabis in the Treatment of Psychiatric Disorders. Medicines 2018, 5, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaccorso, S.; Ricciardi, A.; Zangani, C.; Chiappini, S.; Schifano, F. Cannabidiol (CBD) use in psychiatric disorders: A systematic review. NeuroToxicology 2019, 74, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.; Fogaça, M.V.; Sartim, A.G.; Pereira, V.; Wegener, G.; Guimaraes, F.S.; Joca, S.R. Cannabidiol Induces Rapid and Sustained Antidepressant-Like Effects Through Increased BDNF Signaling and Synaptogenesis in the Prefrontal Cortex. Mol. Neurobiol. 2018, 56, 1070–1081. [Google Scholar] [CrossRef]
- Roehrig, C.S. Mental Disorders Top The List Of The Most Costly Conditions In The United States: $201 Billion. Heal. Aff. 2016, 35, 1130–1135. [Google Scholar] [CrossRef]
- Akil, H.; Gordon, J.; Hen, R.; Javitch, J.A.; Mayberg, H.; McEwen, B.; Meaney, M.J.; Nestler, E.J. Treatment resistant depression: A multi-scale, systems biology approach. Neurosci. Biobehav. Rev. 2018, 84, 272–288. [Google Scholar] [CrossRef]
- Molero, P.; Ramos-Quiroga, J.A.; Martín-Santos, R.; Calvo-Sánchez, E.; Gutiérrez-Rojas, L.; Meana, J.J. Antidepressant Efficacy and Tolerability of Ketamine and Esketamine: A Critical Review. CNS Drugs 2018, 32, 411–420. [Google Scholar] [CrossRef]
- Brien, O.; Lijffijt, M.; Wells, A.; Swann, A.C.; Mathew, S.J.; O’Brien, B. The Impact of Childhood Maltreatment on Intravenous Ketamine Outcomes for Adult Patients with Treatment-Resistant Depression. Pharmaceuticals 2019, 12, 133. [Google Scholar] [CrossRef] [Green Version]
- Shoval, G.; Shbiro, L.; Hershkovitz, L.; Hazut, N.; Zalsman, G.; Mechoulam, R.; Weller, A. Prohedonic Effect of Cannabidiol in a Rat Model of Depression. Neuropsychobiology 2016, 73, 123–129. [Google Scholar] [CrossRef]
- Shbiro, L.; Hen-Shoval, D.; Hazut, N.; Rapps, K.; Dar, S.; Zalsman, G.; Mechoulam, R.; Weller, A.; Shoval, G. Effects of cannabidiol in males and females in two different rat models of depression. Physiol. Behav. 2019, 201, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Reliability of the chronic mild stress model of depression: A user survey. Neurobiol. Stress 2016, 6, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.N.; Hellemans, K.G.; Verma, P.; Gorzalka, B.B.; Weinberg, J. Neurobiology of chronic mild stress: Parallels to major depression. Neurosci. Biobehav. Rev. 2012, 36, 2085–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-L.; Han, Q.-Q.; Gong, W.-Q.; Pan, D.-H.; Wang, L.-Z.; Hu, W.; Yang, M.; Li, B.; Yu, J.; Liu, Q. Microglial activation mediates chronic mild stress-induced depressive- and anxiety-like behavior in adult rats. J. Neuroinflammation 2018, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traslaviña, G.A.A.; Torres, F.P.; Filho, P.C.G.D.B.; Lucio-Oliveira, F.; Franci, C.R. Hypothalamic-pituitary-adrenal axis responsivity to an acute novel stress in female rats subjected to the chronic mild stress paradigm. Brain Res. 2019, 1723, 146402. [Google Scholar] [CrossRef] [PubMed]
- Antoniuk, S.; Bijata, M.; Ponimaskin, E.; Wlodarczyk, J. Chronic unpredictable mild stress for modeling depression in rodents: Meta-analysis of model reliability. Neurosci. Biobehav. Rev. 2019, 99, 101–116. [Google Scholar] [CrossRef]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2016, 6, 78–93. [Google Scholar] [CrossRef] [Green Version]
- Rich, E.L.; Romero, L.M. Exposure to chronic stress downregulates corticosterone responses to acute stressors. Am. J. Physiol. Integr. Comp. Physiol. 2005, 288, R1628–R1636. [Google Scholar] [CrossRef] [Green Version]
- Scorrano, F.; Carrasco, J.; Pastor-Ciurana, J.; Belda, X.; Rami-Bastante, A.; Lavitrano, M.; Armario, A. Validation of the long-term assessment of hypothalamic-pituitary-adrenal activity in rats using hair corticosterone as a biomarker. FASEB J. 2014, 29, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Uarquin, D.G.; Meyer, J.S.; Cardenas, F.P.; Rojas, M.J. Effect of Overcrowding on Hair Corticosterone Concentrations in Juvenile Male Wistar Rats. J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 749–755. [Google Scholar]
- Campos, A.C.; Ortega, Z.; Palazuelos, J.; Fogaça, M.V.; Aguiar, D.C.; Díaz-Alonso, J.; Ortega-Gutiérrez, S.; Vázquez-Villa, H.; Moreira, F.A.; Guzmán, M.; et al. The anxiolytic effect of cannabidiol on chronically stressed mice depends on hippocampal neurogenesis: Involvement of the endocannabinoid system. Int. J. Neuropsychopharmacol. 2013, 16, 1407–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Song, Y.; Bao, T.; Yu, M.; Xu, M.; Guo, Y.; Wang, Y.; Zhang, C.; Zhao, B. Antidepressant-like effects of acupuncture involved the ERK signaling pathway in rats. BMC Complement. Altern. Med. 2016, 16, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, M. Models of Affective Illness: Chronic Mild Stress in the Rat. Curr. Protoc. Pharmacol. 2012, 57, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.; Boss-Williams, K.A.; Weiss, J.M. Effects of chronic mild stress on rats selectively bred for behavior related to bipolar disorder and depression. Physiol. Behav. 2013, 119, 115–129. [Google Scholar] [CrossRef]
- Özkartal, C.; Aricioğlu, F.; Tuzun, E.; Kucukali, C.I. Chronic mild stress-induced anhedonia in rats is coupled with the upregulation of inflammasome sensors: A possible involvement of NLRP1. Psychiatry Clin. Psychopharmacol. 2018, 28, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Taksande, B.; Faldu, D.S.; Dixit, M.P.; Sakaria, J.N.; Aglawe, M.M.; Umekar, M.J.; Kotagale, N. Agmatine attenuates chronic unpredictable mild stress induced behavioral alteration in mice. Eur. J. Pharmacol. 2013, 720, 115–120. [Google Scholar] [CrossRef]
- A Walf, A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Kolcsar, M.; Zsolt, G.; Dogaru, M. Dose dependent effects of serotonergic agents on anxiety. Acta Physiol. Hung. 2014, 101, 479–487. [Google Scholar] [CrossRef]
- Elek, F.; Ferenc, B.; Zsolt, G.; Melinda, K.; Gabriella, D.-N.; Szende, V. Determination of corticosterone from rat hair samples by HPLC-MS method. Bull. Med Sci. 2019, 92, 27–34. [Google Scholar] [CrossRef]
- Fleischhauer, K.; Fruci, D.; Van Endert, P.; Herman, J.; Tanzarella, S.; Wallny, H.-J.; Coulie, P.; Bordignon, C.; Traversari, C. Characterization of antigenic peptides presented by HLA-B44 molecules on tumor cells expressing the geneMAGE-3. Int. J. Cancer 1996, 68, 622–628. [Google Scholar] [CrossRef]
- Jacobson, L.; Sapolsky, R. The Role of the Hippocampus in Feedback Regulation of the Hypothalamic-Pituitary-Adrenocortical Axis*. Endocr. Rev. 1991, 12, 118–134. [Google Scholar] [CrossRef]
- Belzung, C.; De Villemeur, E.B. The design of new antidepressants. Behav. Pharmacol. 2010, 21, 677–689. [Google Scholar] [CrossRef]
- Steiner, M.A.; Marsicano, G.; Nestler, E.J.; Holsboer, F.; Lutz, B.; Wotjak, C.T. Antidepressant-like behavioral effects of impaired cannabinoid receptor type 1 signaling coincide with exaggerated corticosterone secretion in mice. Psychoneuroendocrinology 2007, 33, 54–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linge, R.; Jiménez-Sánchez, L.; Campa, L.; Pilar-Cuéllar, F.; Vidal, R.; Pazos, A.; Adell, A.; Díaz, Á. Cannabidiol induces rapid-acting antidepressant-like effects and enhances cortical 5-HT/glutamate neurotransmission: Role of 5-HT1A receptors. Neuropharmacology 2016, 103, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Roelke, C.T.; Rademacher, D.J.; Cullinan, W.E.; Hillard, C.J. Endocannabinoid Signaling Negatively Modulates Stress-Induced Activation of the Hypothalamic-Pituitary-Adrenal Axis. Endocrinology 2004, 145, 5431–5438. [Google Scholar] [CrossRef]
- Hill, M.N.; Hillard, C.J.; Bambico, F.R.; Patel, S.; Gorzalka, B.B.; Gobbi, G. The Therapeutic Potential of the Endocannabinoid System for the Development of a Novel Class of Antidepressants. Trends Pharmacol. Sci. 2009, 30, 484–493. [Google Scholar] [CrossRef]
- Bakas, T.; Van Nieuwenhuijzen, P.S.; Devenish, S.; McGregor, I.S.; Arnold, J.; Chebib, M. The direct actions of cannabidiol and 2-arachidonoyl glycerol at GABA A receptors. Pharmacol. Res. 2017, 119, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, E.C.D.; Baldasso, G.M.; Bicca, M.A.; Paes, R.S.; Capasso, R.; Dutra, R.C. Terpenoids, Cannabimimetic Ligands, beyond the Cannabis Plant. Molecules 2020, 25, 1567. [Google Scholar] [CrossRef] [Green Version]
- Vieira, G.; Cavalli, J.; Gonçalves, E.C.D.; Braga, S.F.P.; Ferreira, R.S.; Santos, A.R.S.; Cola, M.; Raposo, N.R.B.; Capasso, R.; Dutra, R.C. Antidepressant-Like Effect of Terpineol in an Inflammatory Model of Depression: Involvement of the Cannabinoid System and D2 Dopamine Receptor. Biomolecules 2020, 10, 792. [Google Scholar] [CrossRef]
- Castagné, V.; Moser, P.; Roux, S.; Porsolt, R.D. Rodent Models of Depression: Forced Swim and Tail Suspension Behavioral Despair Tests in Rats and Mice. Curr. Protoc. Neurosci. 2011, 55. [Google Scholar] [CrossRef]
- Ignatowska-Jankowska, B.; Jankowski, M.; Swiergiel, A.H. Cannabidiol decreases body weight gain in rats: Involvement of CB2 receptors. Neurosci. Lett. 2011, 490, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Parray, H.A.; Yun, J.W. Cannabidiol promotes browning in 3T3-L1 adipocytes. Mol. Cell. Biochem. 2016, 416, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Osborne, A.; Solowij, N.; Babic, I.; Huang, X.-F.; Weston-Green, K. Improved Social Interaction, Recognition and Working Memory with Cannabidiol Treatment in a Prenatal Infection (poly I:C) Rat Model. Neuropsychopharmacology 2017, 42, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Wierucka-Rybak, M.; Wolak, M.; Bojanowska, E. The effects of leptin in combination with a cannabinoid receptor 1 antagonist, AM 251, or cannabidiol on food intake and body weight in rats fed a high-fat or a free-choice high sugar diet. J. Physiol. Pharmacol. 2014, 65, 487–496. [Google Scholar] [PubMed]
- He, L.-W.; Zeng, L.; Tian, N.; Li, Y.; He, T.; Tan, D.-M.; Zhang, Q.; Tan, Y. Optimization of food deprivation and sucrose preference test in SD rat model undergoing chronic unpredictable mild stress. Anim. Model. Exp. Med. 2020, 3, 69–78. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kashiwabara, A.; Sakai, T.; Igarashi, K.; Ogata, N.; Watanabe, H.; Ichiyanagi, K.; Yamanouchi, T. Long-term sucrose-drinking causes increased body weight and glucose intolerance in normal male rats. Br. J. Nutr. 2005, 93, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Goshen, I.; Kreisel, T.; Ben-Menachem-Zidon, O.; Licht, T.; Weidenfeld, J.; Ben-Hur, T.; Yirmiya, R. Brain interleukin-1 mediates chronic stress-induced depression in mice via adrenocortical activation and hippocampal neurogenesis suppression. Mol. Psychiatry 2007, 13, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.-G.; Hu, Y.; Wu, D.-L.; Zhu, L.; Chen, C.; Jin, X.; Luo, C.-X.; Wu, H.-Y.; Zhang, J.; Zhu, D. Hippocampal Telomerase Is Involved in the Modulation of Depressive Behaviors. J. Neurosci. 2011, 31, 12258–12269. [Google Scholar] [CrossRef]
- Remus, J.L.; Stewart, L.T.; Camp, R.; Novak, C.M.; Johnson, J.D. Interaction of metabolic stress with chronic mild stress in altering brain cytokines and sucrose preference. Behav. Neurosci. 2015, 129, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Strekalova, T.; Steinbusch, H.W. Measuring behavior in mice with chronic stress depression paradigm. Prog. Neuro-Psychopharmacology Boil. Psychiatry 2010, 34, 348–361. [Google Scholar] [CrossRef]
- Molendijk, M.L.; De Kloet, E.R. Immobility in the forced swim test is adaptive and does not reflect depression. Psychoneuroendocrinology 2015, 62, 389–391. [Google Scholar] [CrossRef] [PubMed]
- De Kloet, E.R.; Molendijk, M.L. Coping with the Forced Swim Stressor: Towards Understanding an Adaptive Mechanism. Neural Plast. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The Rodent Forced Swim Test Measures Stress-Coping Strategy, Not Depression-like Behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanelati, T.; Biojone, C.; Moreira, F.; Guimarães, F.; Joca, S.R. Antidepressant-like effects of cannabidiol in mice: Possible involvement of 5-HT1A receptors. Br. J. Pharmacol. 2009, 159, 122–128. [Google Scholar] [CrossRef] [Green Version]
- El-Alfy, A.T.; Ivey, K.; Robinson, K.; Ahmed, S.; Radwan, M.; Slade, D.; Khan, I.; ElSohly, M.; Ross, S. Antidepressant-like effect of Δ9-tetrahydrocannabinol and other cannabinoids isolated from Cannabis sativa L. Pharmacol. Biochem. Behav. 2010, 95, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Campos, A.C.; Guimaraes, F.S. Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacology 2008, 199, 223–230. [Google Scholar] [CrossRef]
- Sales, A.; Crestani, C.C.; Guimaraes, F.S.; Joca, S.R. Antidepressant-like effect induced by Cannabidiol is dependent on brain serotonin levels. Prog. Neuro-Psychopharmacology Boil. Psychiatry 2018, 86, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C. Acute stress enhances anxiolytic-like drug responses of mice tested in a black and white test box. Eur. Neuropsychopharmacology 1997, 7, 283–288. [Google Scholar] [CrossRef]
- Zimcikova, E.; Simko, J.; Karesova, I.; Kremlacek, J.; Malakova, J. Behavioral effects of antiepileptic drugs in rats: Are the effects on mood and behavior detectable in open-field test? Seizure 2017, 52, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Russell, E.; Koren, G.; Rieder, M.; Van Uum, S. Hair cortisol as a biological marker of chronic stress: Current status, future directions and unanswered questions. Psychoneuroendocrinology 2012, 37, 589–601. [Google Scholar] [CrossRef]
- Yu, T.; Xu, H.; Wang, W.; Li, S.; Chen, Z.; Deng, H. Determination of endogenous corticosterone in rodent’s blood, brain and hair with LC–APCI–MS/MS. J. Chromatogr. B 2015, 1002, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Davenport, M.D.; Tiefenbacher, S.; Lutz, C.K.; Novak, M.A.; Meyer, J. Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. Gen. Comp. Endocrinol. 2006, 147, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Giralt, M.; Armario, A. Individual housing does not influence the adaptation of the pituitary-adrenal axis and other physiological variables to chronic stress in adult male rats. Physiol. Behav. 1989, 45, 477–481. [Google Scholar] [CrossRef]
- Steudte, S.; Stalder, T.; Dettenborn, L.; Klumbies, E.; Foley, P.; Beesdo-Baum, K.; Kirschbaum, C. Decreased hair cortisol concentrations in generalised anxiety disorder. Psychiatry Res. Neuroimaging 2011, 186, 310–314. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stressors | Days of Experiments |
|---|---|
| Cage tilting | 1, 6, 10, 17, 23, 27, 28 |
| Damp bedding | 2, 8, 12, 19, 26 |
| Empty cage | 5, 13, 20, 27 |
| Cage swap | 18 |
| Foreign object | 2, 9, 18 |
| Water jet | 12, 16, 22, 25 |
| Paired housing | 5, 23 |
| Strobe flashing | 3, 6, 11, 17, 20, 22 |
| Hot air steam | 4, 11, 16, 24 |
| Food deprivation | 6, 13, 20, 27 |
| Water deprivation | 4, 9, 15, 24 |
| Parameter | Acute Experiment | Chronic Experiment | ||||
|---|---|---|---|---|---|---|
| Control (n = 8) | CBD Treated (n = 8) | p-Value | CUMS (n = 10) | CUMS + CBD (n = 8) | p-Value | |
| Open-arm entries (#) | 3.00 ± 0.906 | 2.75 ± 0.725 | 0.8326 | 2.30 ± 0.423 | 2.13 ± 0.295 | 0.7515 |
| Open-arm preference (%) | 25.84 (0.00–38.46) | 27.21 (0.00–66.67) | 0.9587 | 12.92 (10.0–22.04) | 10.82 (9.64–18.26) | 0.3714 |
| Open-arm time (s) | 17.12 ± 4.82 | 22.51 ± 7.72 | 0.5636 | 26.42 ± 6.616 | 27.27 ± 5.963 | 0.9287 |
| Closed-arm time (s) | 200.9 ± 13.51 | 193.3 ± 12.44 | 0.6840 | 196.0 ± 13.79 | 196.4 ± 10.68 | 0.9802 |
| Rearing (s) | 28.17 ± 3.43 | 18.55 ± 2.51 | 0.0401* | 21.92 ± 2.056 | 26.35 ± 2.897 | 0.1560 |
| Head dips (#) | 7.62 ± 2.28 | 7.75 ± 1.75 | 0.9660 | 5.50 ± 0.833 | 5.38 ± 0.905 | 0.9206 |
| Distance moved (cm) | 1021 ± 109.3 | 1014 ± 97.56 | 0.9609 | 973.1 ± 112.7 | 916.5 ± 79.33 | 0.7635 |
| Stretch attend posture (s) | 54.50 ±8.20 | 71.86 ± 10.14 | 0.2044 | 25.77 ± 4.66 | 39.22 ± 7.28 | 0.1255 |
| Velocity in open arm (cm/s) | 4.16 ± 0.72 | 4.59 ± 1.11 | 0.7515 | 3.83 ± 0.439 | 4.92 ± 0.403 | 0.0907 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gáll, Z.; Farkas, S.; Albert, Á.; Ferencz, E.; Vancea, S.; Urkon, M.; Kolcsár, M. Effects of Chronic Cannabidiol Treatment in the Rat Chronic Unpredictable Mild Stress Model of Depression. Biomolecules 2020, 10, 801. https://doi.org/10.3390/biom10050801
Gáll Z, Farkas S, Albert Á, Ferencz E, Vancea S, Urkon M, Kolcsár M. Effects of Chronic Cannabidiol Treatment in the Rat Chronic Unpredictable Mild Stress Model of Depression. Biomolecules. 2020; 10(5):801. https://doi.org/10.3390/biom10050801
Chicago/Turabian StyleGáll, Zsolt, Szidónia Farkas, Ákos Albert, Elek Ferencz, Szende Vancea, Melinda Urkon, and Melinda Kolcsár. 2020. "Effects of Chronic Cannabidiol Treatment in the Rat Chronic Unpredictable Mild Stress Model of Depression" Biomolecules 10, no. 5: 801. https://doi.org/10.3390/biom10050801
APA StyleGáll, Z., Farkas, S., Albert, Á., Ferencz, E., Vancea, S., Urkon, M., & Kolcsár, M. (2020). Effects of Chronic Cannabidiol Treatment in the Rat Chronic Unpredictable Mild Stress Model of Depression. Biomolecules, 10(5), 801. https://doi.org/10.3390/biom10050801