The Influence of Melatonin and Light on VEGF Secretion in Primary RPE Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Single-Eye RPE Cell Culture
2.2. Cell Treatment
2.2.1. Light Impulse
2.2.2. Treatment with Melatonin
2.2.3. Combined Treatment with Light and Melatonin
2.3. ELISA
2.4. Western Blot
2.5. Statistics
3. Results
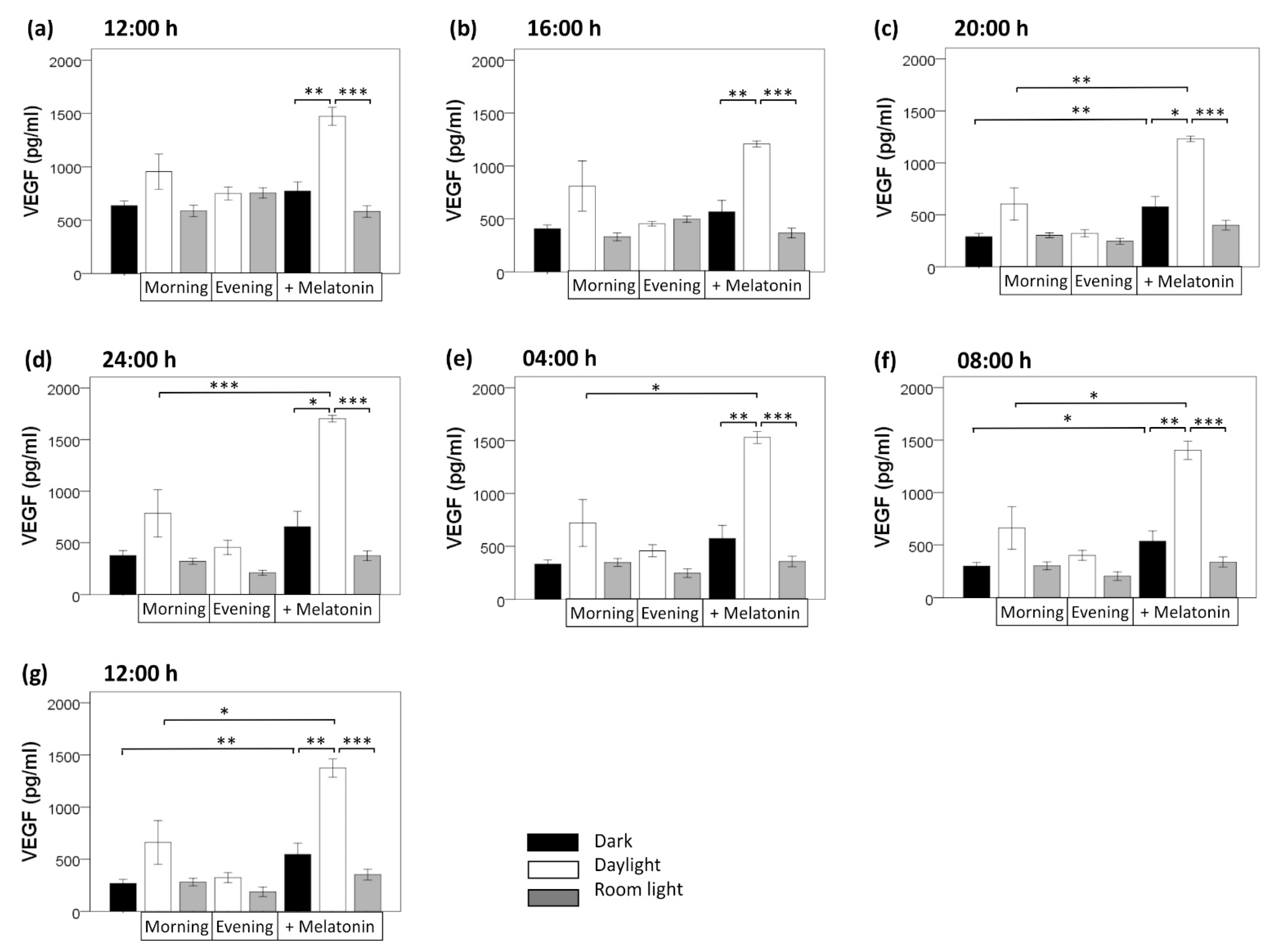
3.1. Circadian Variation
3.2. Light Induction
3.3. Melatonin Treatment
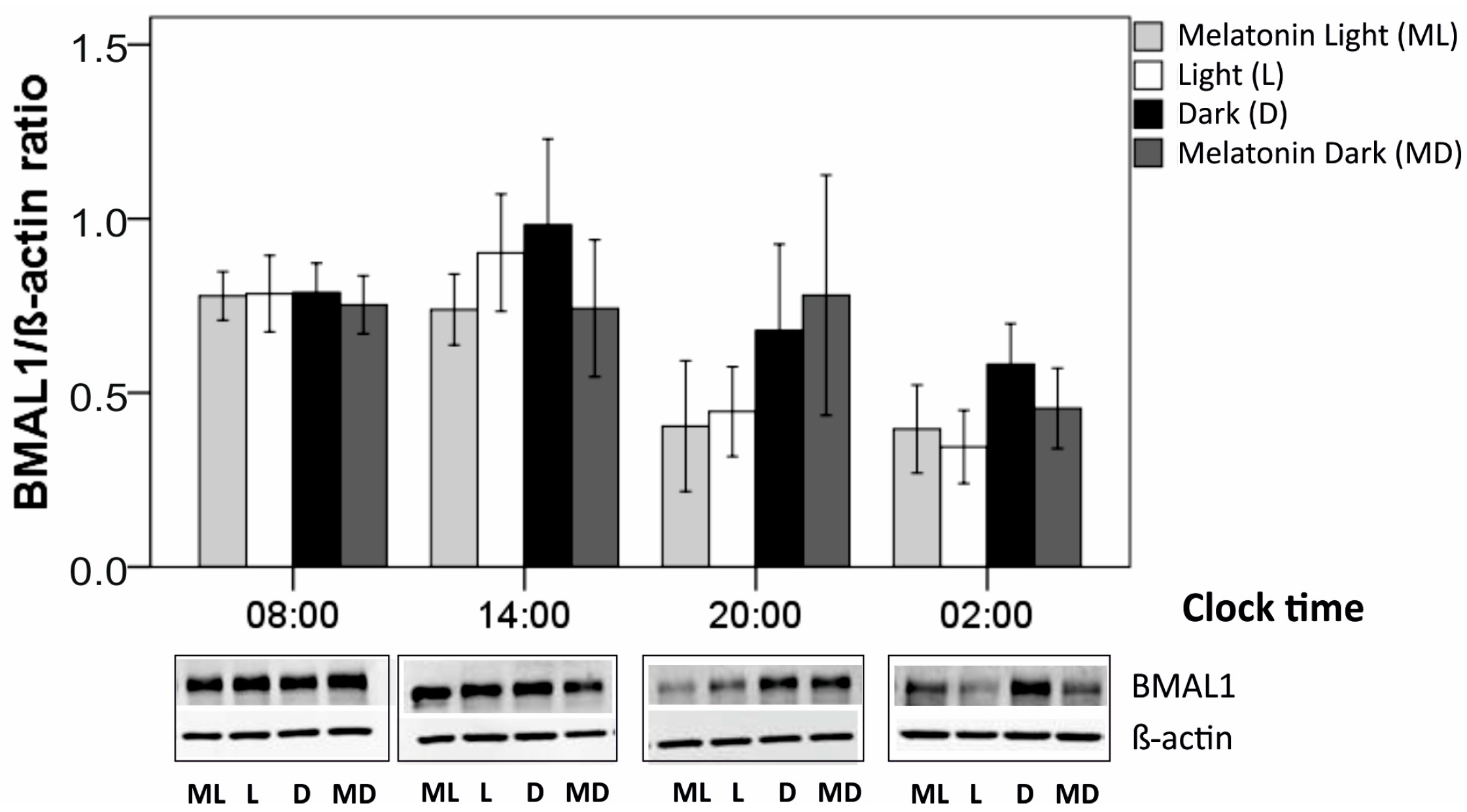
3.4. BMAL-1 expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Geniez, M.; Kurihara, T.; Sekiyama, E.; Maldonado, A.E.; D’Amore, P.A. An essential role for RPE-derived soluble VEGF in the maintenance of the choriocapillaris. Proc. Natl. Acad. Sci. USA 2009, 106, 18751–18756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, K.M.; Saint-Geniez, M.; Walshe, T.; Zahr, A.; D’Amore, P.A. Expression and role of VEGF in the adult retinal pigment epithelium. Invest. Ophthalmol. Vis. Sci. 2011, 52, 9478–9487. [Google Scholar] [CrossRef] [Green Version]
- Blaauwgeers, H.G.; Holtkamp, G.M.; Rutten, H.; Witmer, A.N.; Koolwijk, P.; Partanen, T.A.; Alitalo, K.; Kroon, M.E.; Kijlstra, A.; van Hinsbergh, V.W.; et al. Polarized vascular endothelial growth factor secretion by human retinal pigment epithelium and localization of vascular endothelial growth factor receptors on the inner choriocapillaris. Evidence for a trophic paracrine relation. Am. J. Pathol. 1999, 155, 421–428. [Google Scholar] [CrossRef]
- Klettner, A. Physiological Functions of VEGF in the Retina and Its Possible Implications for prolonged Anti-VEGF therapy. In Vascular Endothelial Growth Factor—Biology, Regulation and Clinical Significance; Parker, M., Ed.; Nova Science Publisher: New York, NY, USA, 2013; pp. 117–136. [Google Scholar]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef]
- Pe’er, J.; Shweiki, D.; Itin, A.; Hemo, I.; Gnessin, H.; Keshet, E. Hypoxia-induced expression of vascular endothelial growth factor by retinal cells is a common factor in neovascularizing ocular diseases. Lab. Investig. Lab. Invest. 1995, 72, 638–645. [Google Scholar]
- Penn, J.S.; Madan, A.; Caldwell, R.B.; Bartoli, M.; Caldwell, R.W.; Hartnett, M.E. Vascular endothelial growth factor in eye disease. Prog. Retin. Eye Res. 2008, 27, 331–371. [Google Scholar] [CrossRef] [Green Version]
- Saint-Geniez, M.; Maharaj, A.S.; Walshe, T.E.; Tucker, B.A.; Sekiyama, E.; Kurihara, T.; Darland, D.C.; Young, M.J.; D’Amore, P.A. Endogenous VEGF is required for visual function: Evidence for a survival role on muller cells and photoreceptors. PLoS ONE 2008, 3, e3554. [Google Scholar] [CrossRef] [Green Version]
- Byeon, S.H.; Lee, S.C.; Choi, S.H.; Lee, H.K.; Lee, J.H.; Chu, Y.K.; Kwon, O.W. Vascular endothelial growth factor as an autocrine survival factor for retinal pigment epithelial cells under oxidative stress via the VEGF-R2/PI3K/Akt. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1190–1197. [Google Scholar] [CrossRef] [Green Version]
- Peters, S.; Heiduschka, P.; Julien, S.; Ziemssen, F.; Fietz, H.; Bartz-Schmidt, K.U.; Schraermeyer, U. Ultrastructural findings in the primate eye after intravitreal injection of bevacizumab. Mol. Aspects Med. 2007, 143, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Kaya, L.; Flach, J.; Lassen, J.; Treumer, F.; Roider, J. Basal and apical regulation of VEGF-A and placenta growth factor in the RPE/choroid and primary RPE. Mol. Vis. 2015, 21, 736–748. [Google Scholar]
- Klettner, A.; Westhues, D.; Lassen, J.; Bartsch, S.; Roider, J. Regulation of constitutive vascular endothelial growth factor secretion in retinal pigment epithelium/choroid organ cultures: p38, nuclear factor kappaB, and the vascular endothelial growth factor receptor-2/phosphatidylinositol 3 kinase pathway. Mol. Vis. 2013, 19, 281–291. [Google Scholar]
- Faby, H.; Hillenkamp, J.; Roider, J.; Klettner, A. Hyperthermia-induced upregulation of vascular endothelial growth factor in retinal pigment epithelial cells is regulated by mitogen-activated protein kinases. Graefes Arch. Clin. Exp. Ophthalmol. 2014, 252, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Koinzer, S.; Meyer, T.; Roider, J. Toll-like receptor 3 activation in retinal pigment epithelium cells–Mitogen-activated protein kinase pathways of cell death and vascular endothelial growth factor secretion. Acta. Ophthalmol. 2013, 91, e211–e218. [Google Scholar] [CrossRef]
- Klettner, A.; Roider, J. Constitutive and oxidative-stress-induced expression of VEGF in the RPE are differently regulated by different Mitogen-activated protein kinases. Graefes Arch. Clin. Exp. Ophthalmol. 2009, 247, 1487–1492. [Google Scholar] [CrossRef]
- Kunchithapautham, K.; Rohrer, B. Sublytic membrane-attack-complex (MAC) activation alters regulated rather than constitutive vascular endothelial growth factor (VEGF) secretion in retinal pigment epithelium monolayers. J. Biol. Chem. 2011, 286, 23717–23724. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A.; Roider, J. Mechanisms of Pathological VEGF Production in the Retina and Modification with VEGF-Antagonists. In Studies on Retinal and Choroidal Disorders; Stratton, R.D., Hauswirth, W.W., Gardner, T.W., Eds.; Humana Press: New York, NY, USA, 2012; pp. 277–306. [Google Scholar]
- Klettner, A. Oxidative stress induced cellular signaling in RPE cells. Front. Biosci. 2012, 4, 392–411. [Google Scholar] [CrossRef]
- Tischer, E.; Mitchell, R.; Hartman, T.; Silva, M.; Gospodarowicz, D.; Fiddes, J.C.; Abraham, J.A. The human gene for vascular endothelial growth factor. Multiple protein forms are encoded through alternative exon splicing. J. Biol. Chem. 1991, 266, 11947–11954. [Google Scholar] [CrossRef]
- Ozaki, H.; Yu, A.Y.; Della, N.; Ozaki, K.; Luna, J.D.; Yamada, H.; Hackett, S.F.; Okamoto, N.; Zack, D.J.; Semenza, G.L.; et al. Hypoxia inducible factor-1alpha is increased in ischemic retina: Temporal and spatial correlation with VEGF expression. Invest. Ophthalmol. Vis. Sci. 1999, 40, 182–189. [Google Scholar]
- Oltra, M.; Vidal-Gil, L.; Maisto, R.; Sancho-Pelluz, J.; Barcia, J.M. Oxidative stress-induced angiogenesis is mediated by miR-205-5p. J. Cell. Mol. Sci. 2020, 24, 1428–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L.D.; Cao, Y. Clock controls angiogenesis. Cell Cycle 2013, 12, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamaleia, N.F.; Lisnyak, I.A.; Shishko, E.D.; Mamchur, A.A.; Prokopenko, I.V.; Kholin, V.V. Chronobiological approaches to antiangiogenic photodynamic therapy of tumors: The first experimental evaluation. Exp. Oncol. 2012, 34, 364–366. [Google Scholar] [PubMed]
- Koyanagi, S.; Kuramoto, Y.; Nakagawa, H.; Aramaki, H.; Ohdo, S.; Soeda, S.; Shimeno, H. A molecular mechanism regulating circadian expression of vascular endothelial growth factor in tumor cells. Cancer Res. 2003, 63, 7277–7283. [Google Scholar]
- Endo, I.; Mitsui, T.; Nishino, M.; Oshima, Y.; Matsumoto, T. Diurnal fluctuation of edema synchronized with plasma VEGF concentration in a patient with POEMS syndrome. Intern. Med. 2002, 41, 1196–1198. [Google Scholar] [CrossRef] [Green Version]
- Tosini, G.; Menaker, M. Circadian rhythms in cultured mammalian retina. Science 1996, 272, 419–421. [Google Scholar] [CrossRef]
- Ko, G.Y. Circadian regulation in the retina: From molecules to network. Eur. J. Neurosci. 2020, 51, 194–216. [Google Scholar] [CrossRef] [Green Version]
- Ruan, G.X.; Zhang, D.Q.; Zhou, T.; Yamazaki, S.; McMahon, D.G. Circadian organization of the mammalian retina. Proc. Natl. Acad. Sci. USA 2006, 103, 9703–9708. [Google Scholar] [CrossRef] [Green Version]
- McMahon, D.G.; Iuvone, P.M.; Tosini, G. Circadian organization of the mammalian retina: From gene regulation to physiology and diseases. Prog. Retin. Eye Res. 2014, 39, 58–76. [Google Scholar] [CrossRef] [Green Version]
- Bobu, C.; Hicks, D. Regulation of retinal photoreceptor phagocytosis in a diurnal mammal by circadian clocks and ambient lighting. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3495–3502. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, J.; Nakagawa, T.; Ogawa, K.; Mori, M. The effect of colchicine on the diurnal variation of phagocytosis in mouse retinal pigment epithelium. Exp. Eye Res. 1987, 44, 755–765. [Google Scholar] [CrossRef]
- Irschick, E.U.; Haas, G.; Geiger, M.; Singer, W.; Ritsch-Marte, M.; Konwalinka, G.; Frick, M.; Gottinger, W.; Huemer, H.P. Phagocytosis of human retinal pigment epithelial cells: Evidence of a diurnal rhythm, involvement of the cytoskeleton and interference of antiviral drugs. Ophthalmic Res. 2006, 38, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Jia, L.; Shelby, S.J.; Ganios, A.M.; Feathers, K.; Thompson, D.A.; Zacks, D.N. Circadian and noncircadian modulation of autophagy in photoreceptors and retinal pigment epithelium. Invest. Ophthalmol. Vis. Sci. 2014, 55, 3237–3246. [Google Scholar] [CrossRef] [PubMed]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, K.; Sengupta, A.; Tosini, M.; Contreras-Alcantara, S.; Tosini, G. Circadian regulation of the PERIOD 2:LUCIFERASE bioluminescence rhythm in the mouse retinal pigment epithelium-choroid. Mol. Vis. 2010, 16, 2605–2611. [Google Scholar]
- Baba, K.; Davidson, A.J.; Tosini, G. Melatonin Entrains PER2::LUC Bioluminescence Circadian Rhythm in the Mouse Cornea. Invest. Ophthalmol. Vis. Sci. 2015, 56, 4753–4758. [Google Scholar] [CrossRef] [Green Version]
- Laurent, V.; Sengupta, A.; Sánchez-Bretaño, A.; Hicks, D.; Tosini, G. Melatonin signaling affects the timing in the daily rhythm of phagocytic activity by the retinal pigment epithelium. Exp. Eye Res. 2017, 165, 90–95. [Google Scholar] [CrossRef]
- Liang, F.Q.; Green, L.; Wang, C.; Alssadi, R.; Godley, B.F. Melatonin protects human retinal pigment epithelial (RPE) cells against oxidative stress. Exp. Eye Res. 2004, 78, 1069–1075. [Google Scholar] [CrossRef]
- Blasiak, J.; Reiter, R.J.; Kaarniranta, K. Melatonin in Retinal Physiology and Pathology: The Case of Age-Related Macular Degeneration. Oxidative Med. Cell. Longev. 2016, 2016, 6819736. [Google Scholar] [CrossRef]
- Rastmanesh, R. Potential of melatonin to treat or prevent age-related macular degeneration through stimulation of telomerase activity. Med. Hypotheses. 2011, 76, 79–85. [Google Scholar] [CrossRef]
- Lai, Y.H.; Hu, D.N.; Rosen, R.; Sassoon, J.; Chuang, L.Y.; Wu, K.Y.; Wu, W.C. Hypoxia-induced vascular endothelial growth factor secretion by retinal pigment epithelial cells is inhibited by melatonin via decreased accumulation of hypoxia-inducible factors-1alpha protein. Clin. Exp. Ophthalmol. 2017, 45, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Tanaka, T.; Doi, H.; Kanai, H.; Maeno, T.; Itakura, H.; Iida, T.; Kimura, Y.; Kishi, S.; Kurabayashi, M. Visible light exposure induces VEGF gene expression through activation of retinoic acid receptor-alpha in retinoblastoma Y79 cells. Am. J. Physiol. Cell Physiol. 2005, 288, C913–C920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiencke, A.K.; Kiilgaard, J.F.; Nicolini, J.; Bundgaard, M.; Ropke, C.; La Cour, M. Growth of cultured porcine retinal pigment epithelial cells. Acta. Ophthalmol. Scand. 2003, 81, 170–176. [Google Scholar] [CrossRef]
- Klettner, A.; Roider, J. Comparison of bevacizumab, ranibizumab, and pegaptanib in vitro: Efficiency and possible additional pathways. Invest. Ophthalmol. Vis. Sci. 2008, 49, 4523–4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernt, M.; Neubauer, A.S.; Liegl, R.; Eibl, K.H.; Alge, C.S.; Lackerbauer, C.A.; Ulbig, M.W.; Kampik, A. Cytoprotective effects of a blue light-filtering intraocular lens on human retinal pigment epithelium by reducing phototoxic effects on vascular endothelial growth factor-alpha, Bax, and Bcl-2 expression. J. Cataract Refract. Surg. 2009, 35, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Hu, A.; Luo, Y.; Sun, W.; Hu, X.; Tang, S. Interleukin-4 and melatonin ameliorate high glucose and interleukin-1β stimulated inflammatory reaction in human retinal endothelial cells and retinal pigment epithelial cells. Mol. Vis. 2014, 20, 921–928. [Google Scholar]
- Fu, Y.; Tang, M.; Fan, Y.; Zou, H.; Sun, X.; Xu, X. Anti-apoptotic effects of melatonin in retinal pigment epithelial cells. Front. Biosci. 2012, 17, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Refinetti, R. Circadian Physiology, 3rd ed.; CRC press: Boca Raton, FL, USA, 2016; p. 89. [Google Scholar]
- Ikarashi, R.; Akechi, H.; Kanda, Y.; Ahmad, A.; Takeuchi, K.; Morioka, E.; Sugiyama, T.; Ebisawa, T.; Ikeda, M.; Ikeda, M. Regulation of molecular clock oscillations and phagocytic activity via muscarinic Ca(2+) signaling in human retinal pigment epithelial cells. Sci. Rep. 2017, 7, 44175. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A. Retinal Pigment Epithelium Cell Culture. In Retinal Pigment Epithelium in Health and Disease; Klettner, A., Dithmar, S., Eds.; Springer Nature: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Ueta, T.; Inoue, T.; Yuda, K.; Furukawa, T.; Yanagi, Y.; Tamaki, Y. Intense physiological light upregulates vascular endothelial growth factor and enhances choroidal neovascularization via peroxisome proliferator-activated receptor gamma coactivator-1alpha in mice. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1366–1371. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, X.; Zhang, D.; Zhou, F.; Wang, D.; Wei, Y.; Gao, F.; Xie, L.; Jia, G.; Wu, W.; et al. Blueberry anthocyanins: Protection against ageing and light-induced damage in retinal pigment epithelial cells. Br. J. Nutr. 2012, 108, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Lu, X.; Hu, Y.; Yang, B.; Tsui, C.K.; Yu, S.; Lu, L.; Liang, X. Melatonin attenuated retinal neovascularization and neuroglial dysfunction by inhibition of HIF-1alpha-VEGF pathway in oxygen-induced retinopathy mice. J. Pineal Res. 2018, 64, e12473. [Google Scholar] [CrossRef] [PubMed]
- Ferreira de Melo, I.M.; Martins Ferreira, C.G.; Lima da Silva Souza, E.H.; Almeida, L.L.; Bezerra de Sá, F.; Cavalcanti Lapa Neto, C.J.; Paz de Castro, M.V.; Teixeira, V.W.; Coelho Teixeira Á, A. Melatonin regulates the expression of inflammatory cytokines, VEGF and apoptosis in diabetic retinopathy in rats. Chem. Biol. Interact. 2020, 327, 109183. [Google Scholar] [CrossRef] [PubMed]
- Bardak, H.; Uğuz, A.C.; Bardak, Y. Protective effects of melatonin and memantine in human retinal pigment epithelium (ARPE-19) cells against 2-ethylpyridine-induced oxidative stress: Implications for age-related macular degeneration. Cutan. Ocul. Toxicol. 2018, 37, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Fu, Z.J.; Lo, A.C. Hypoxia-induced oxidative stress in ischemic retinopathy. Oxid. Med. Cell. Longev. 2012, 2012, 426769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behl, T.; Kaur, I.; Kotwani, A. Implication of oxidative stress in progression of diabetic retinopathy. Surv. Ophthalmol. 2016, 61, 187–196. [Google Scholar] [CrossRef] [PubMed]
- van Reyk, D.M.; Gillies, M.C.; Davies, M.J. The retina: Oxidative stress and diabetes. Redox Rep. Commun. Free Radic. Res. 2003, 8, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.; Zhang, N.; Sreekumar, P.G.; Spee, C.K.; Rodriguez, A.; Barron, E.; Hinton, D.R. Stimulation of apical and basolateral VEGF-A and VEGF-C secretion by oxidative stress in polarized retinal pigment epithelial cells. Mol. Vis. 2006, 12, 1649–1659. [Google Scholar]
- Gerber, H.P.; McMurtrey, A.; Kowalski, J.; Yan, M.; Keyt, B.A.; Dixit, V.; Ferrara, N. Vascular endothelial growth factor regulates endothelial cell survival through the phosphatidylinositol 3’-kinase/Akt signal transduction pathway. Requirement for Flk-1/KDR activation. J. Biol. Chem. 1998, 273, 30336–30343. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Ozawa, Y.; Kubota, S.; Hirasawa, M.; Miyake, S.; Noda, K.; Tsubota, K.; Kadonosono, K.; Ishida, S. Neuroprotective response after photodynamic therapy: Role of vascular endothelial growth factor. J. Neuroinflammation 2011, 8, 176. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, Y.; Hirata, A.; Ishikawa, S.; Okinami, S. Changes in choriocapillaris fenestration of rat eyes after intravitreal bevacizumab injection. Graefes. Arch. Clin. Exp. Ophthalmol. 2009, 247, 1089–1094. [Google Scholar] [CrossRef]
- Kurihara, T.; Westenskow, P.D.; Bravo, S.; Aguilar, E.; Friedlander, M. Targeted deletion of Vegfa in adult mice induces vision loss. J. Clin. Invest. 2012, 122, 4213–4217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marneros, A.G.; Fan, J.; Yokoyama, Y.; Gerber, H.P.; Ferrara, N.; Crouch, R.K.; Olsen, B.R. Vascular endothelial growth factor expression in the retinal pigment epithelium is essential for choriocapillaris development and visual function. Am. J. Pathol. 2005, 167, 1451–1459. [Google Scholar] [CrossRef] [Green Version]
- Gemenetzi, M.; Lotery, A.J.; Patel, P.J. Risk of geographic atrophy in age-related macular degeneration patients treated with intravitreal anti-VEGF agents. Eye 2017, 31, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Bian, J.; Wang, Z.; Dong, Y.; Chen, Y. Effect of monochromatic light on circadian rhythmic expression of clock genes and arylalkylamine N-acetyltransferase in chick retina. Chronobiol. Int. 2017, 34, 1149–1157. [Google Scholar] [CrossRef]
- Stepicheva, N.A.; Weiss, J.; Shang, P.; Yazdankhah, M.; Ghosh, S.; Bhutto, I.A.; Hose, S.; Zigler, J.S., Jr.; Sinha, D. Melatonin as the Possible Link Between Age-Related Retinal Regeneration and the Disrupted Circadian Rhythm in Elderly. Adv. Exp. Med. Biol. 2019, 1185, 45–49. [Google Scholar] [CrossRef]
- Schmid-Kubista, K.E.; Glittenberg, C.G.; Cezanne, M.; Holzmann, K.; Neumaier-Ammerer, B.; Binder, S. Daytime levels of melatonin in patients with age-related macular degeneration. Acta. Ophthalmol. 2009, 87, 89–93. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | N | 24h-Clock Time Comparison Chi2 | 24h-Clock Time Comparison Pa |
|---|---|---|---|
| Dark control | 40 | 102.99 | 5.9 × 10−20 |
| Daylight morning (7:40–8:00) | 8 | 24.21 | 4.8 × 10−4 |
| Room light morning (7:40–8:00) | 18 | 43.07 | 1.3 × 10−7 |
| Daylight evening (19:40–20:00) | 5 | 21.26 | 0.002 |
| Room light evening (19:40–20:00) | 5 | 22.97 | 0.001 |
| Melatonin dark | 19 | 23.89 | 0.001 |
| Melatonin daylight (7:40–8:00) | 5 | 19.20 | 0.004 |
| Melatonin room light (7:40–8:00) | 14 | 17.85 | 0.007 |
| Clock Time (hrs) | Ratio BMAL1/ β actin | MHb | VHb | VDb | MDb | Chi2 | Pa |
|---|---|---|---|---|---|---|---|
| 8:00 | N | 5 | 5 | 5 | 5 | ||
| Mean | 0.778 | 0.785 | 0.788 | 0.753 | 0.36 | 0.948 | |
| SEM | 0.070 | 0.109 | 0.084 | 0.083 | |||
| 14:00 | N | 5 | 5 | 5 | 5 | ||
| Mean | 0.739 | 0.902 | 0.983 | 0.743 | 2.04 | 0.564 | |
| SEM | 0.102 | 0.168 | 0-246 | 0.197 | |||
| 20.00 | N | 5 | 5 | 5 | 5 | ||
| Mean | 0.404 | 0.446 | 0.677 | 0.780 | 0.28 | 0.041 | |
| SEM | 0.188 | 0.129 | 0.248 | 0.345 | |||
| 02.00 | N | 5 | 5 | 5 | 5 | ||
| Mean | 0.396 | 0.345 | 0.582 | 0.455 | 9.00 | 0.029 | |
| SEM | 0.126 | 0.105 | 0.117 | 0.115 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klettner, A.; Kampers, M.; Töbelmann, D.; Roider, J.; Dittmar, M. The Influence of Melatonin and Light on VEGF Secretion in Primary RPE Cells. Biomolecules 2021, 11, 114. https://doi.org/10.3390/biom11010114
Klettner A, Kampers M, Töbelmann D, Roider J, Dittmar M. The Influence of Melatonin and Light on VEGF Secretion in Primary RPE Cells. Biomolecules. 2021; 11(1):114. https://doi.org/10.3390/biom11010114
Chicago/Turabian StyleKlettner, Alexa, Miriam Kampers, Daniela Töbelmann, Johann Roider, and Manuela Dittmar. 2021. "The Influence of Melatonin and Light on VEGF Secretion in Primary RPE Cells" Biomolecules 11, no. 1: 114. https://doi.org/10.3390/biom11010114
APA StyleKlettner, A., Kampers, M., Töbelmann, D., Roider, J., & Dittmar, M. (2021). The Influence of Melatonin and Light on VEGF Secretion in Primary RPE Cells. Biomolecules, 11(1), 114. https://doi.org/10.3390/biom11010114