Kartogenin Enhances Chondrogenic Differentiation of MSCs in 3D Tri-Copolymer Scaffolds and the Self-Designed Bioreactor System

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Rat MSCs
2.2. Osteogenic, Chondrogenic, Adipogenic Differentiation Evaluation of rMSCs
2.3. rMSC Characteristic Analysis by Flow Cytometry (FC)
2.4. Synthesis of Tri-Copolymer Scaffolds
2.5. Measurement of Tri-Copolymer Scaffolds Cross-Linking Degree
2.6. Self-Designed Bioreactor System
2.7. Cell Seeding and Culture in the Self-Designed Bioreactor System
2.8. Scanning Electron Microscopy (SEM)
2.9. MSCs Condensation Examination in 2D and 3D Culture
2.10. Quantitative Real-Time PCR (Q-PCR)
2.11. Hematoxylin/Eosin and Immunohistochemical (IHC) Staining
2.12. Statistics Analysis
3. Results
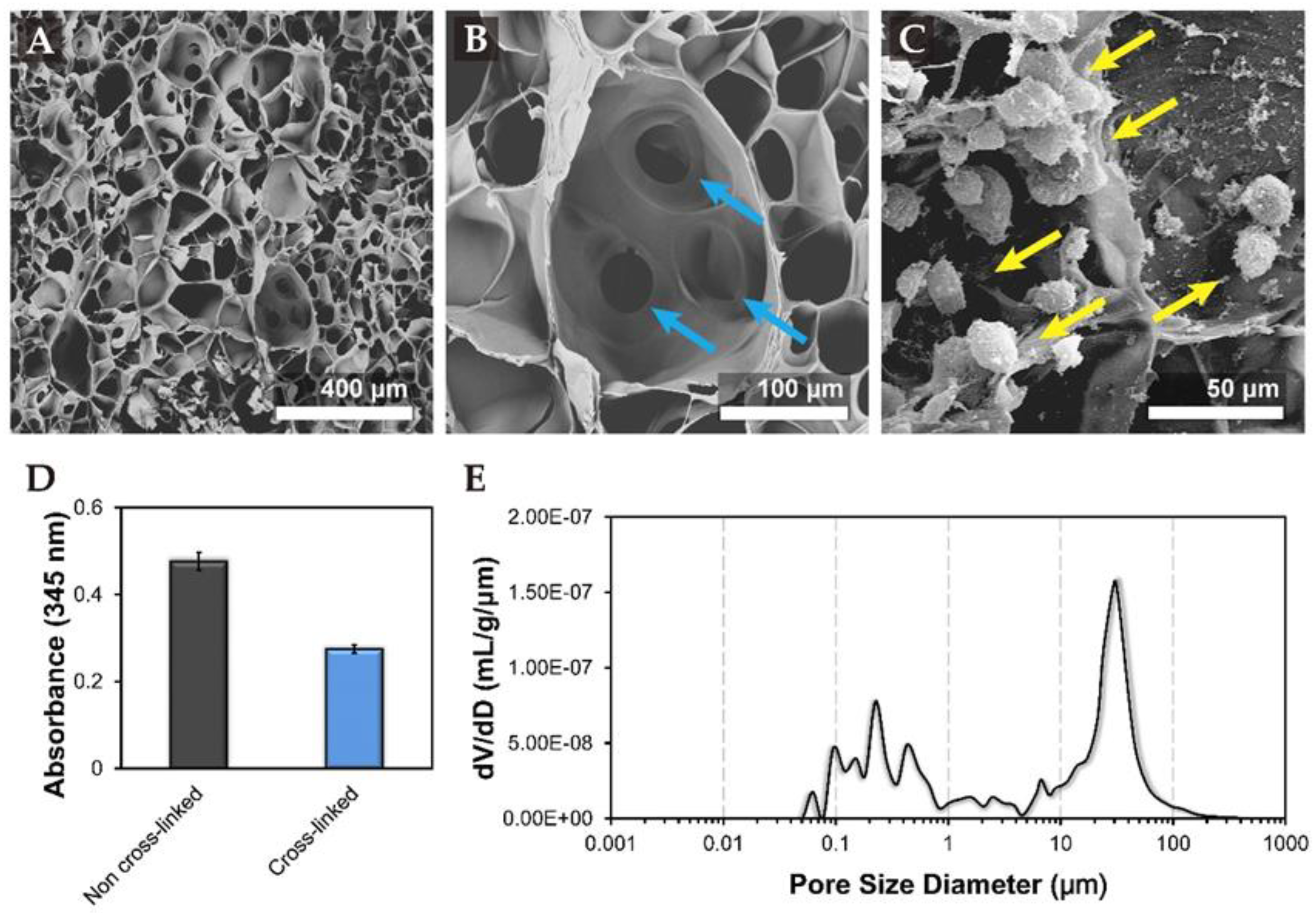
3.1. Characteristics of Tri-Copolymer Scaffold
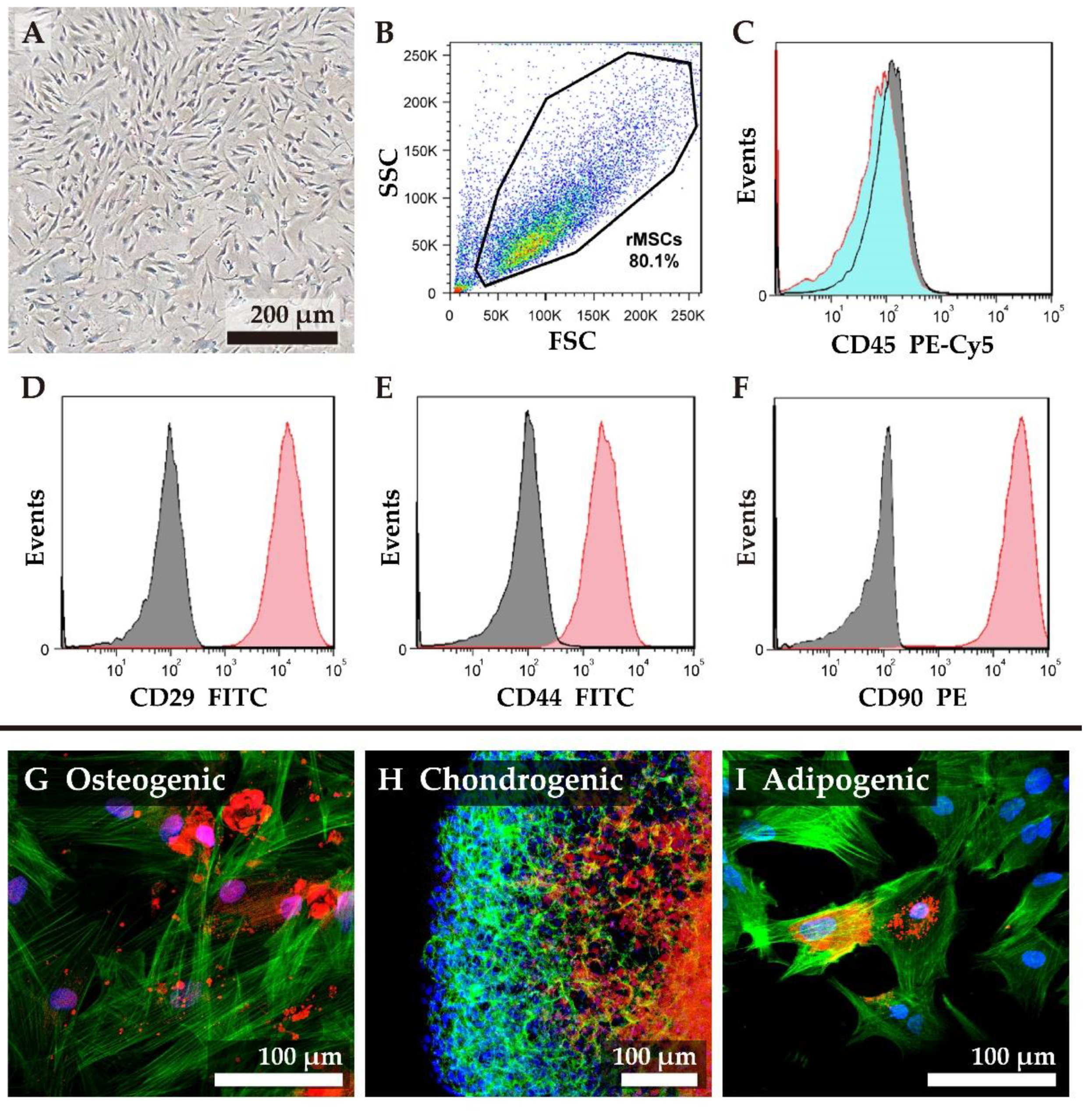
3.2. Isolation and Characterization of Rat MSCs
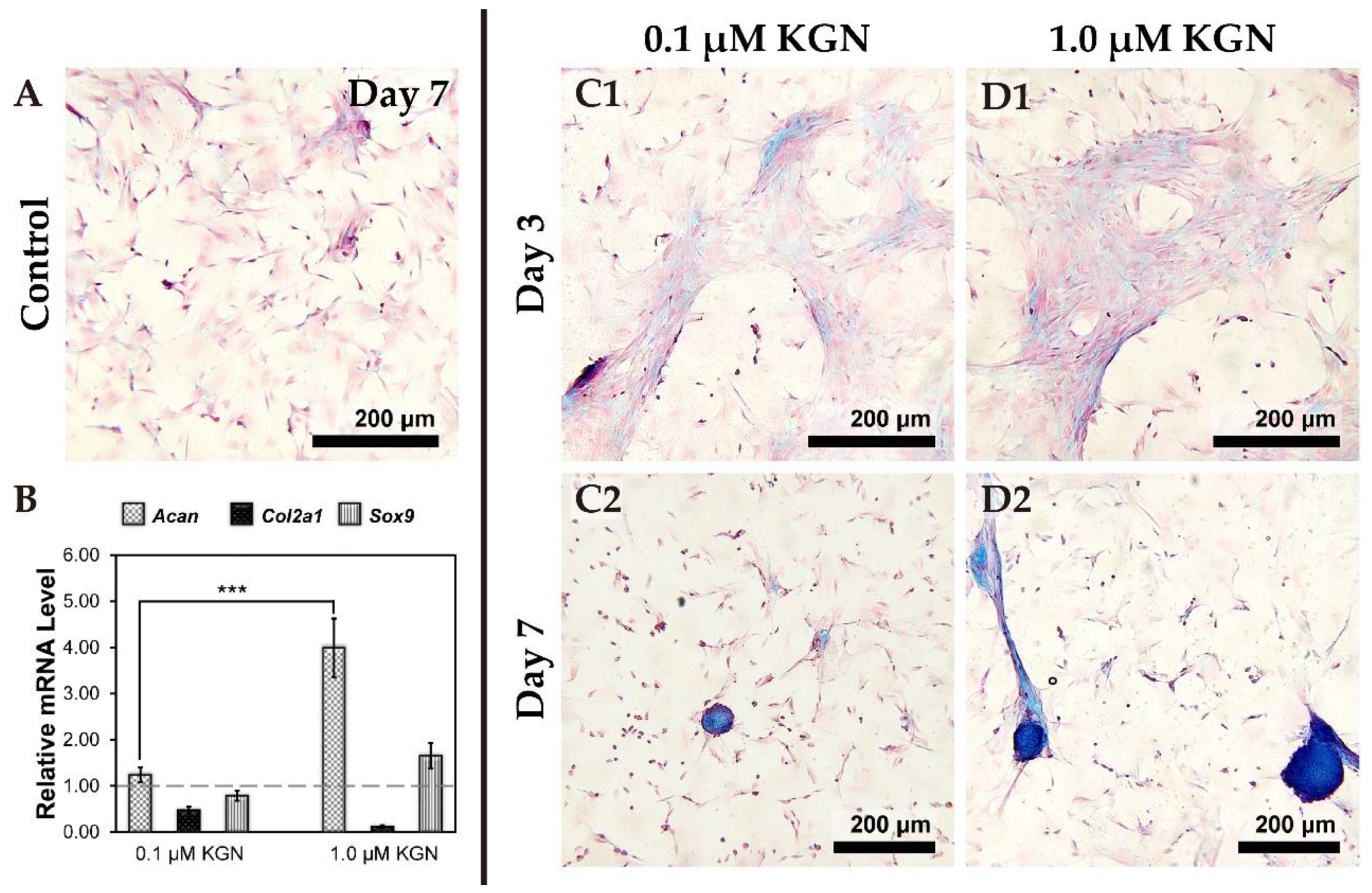
3.3. Determining the Optimum Dose of KGN for Chondrogenesis
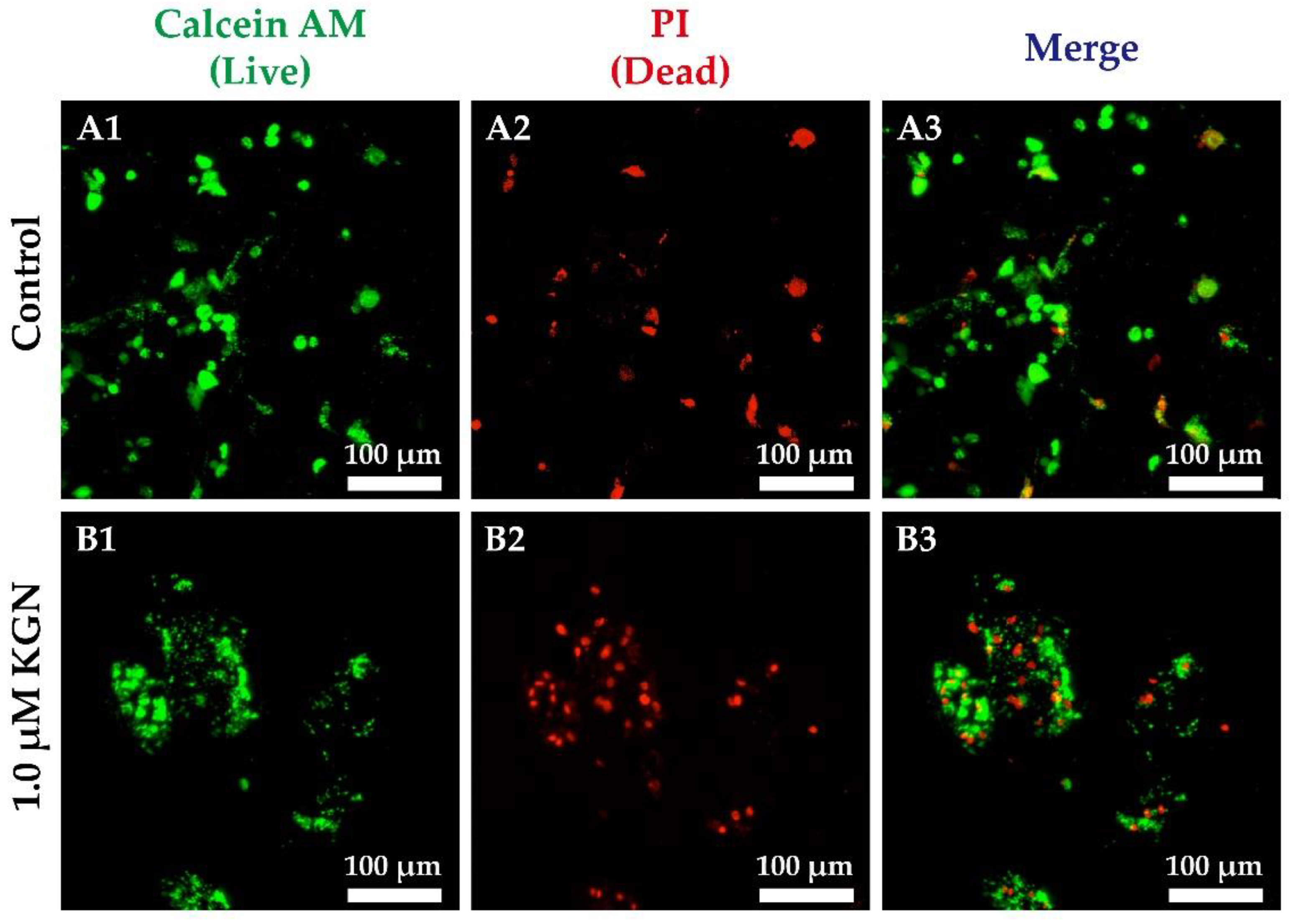
3.4. Live/Dead Staining in Static 2D and 3D Culture
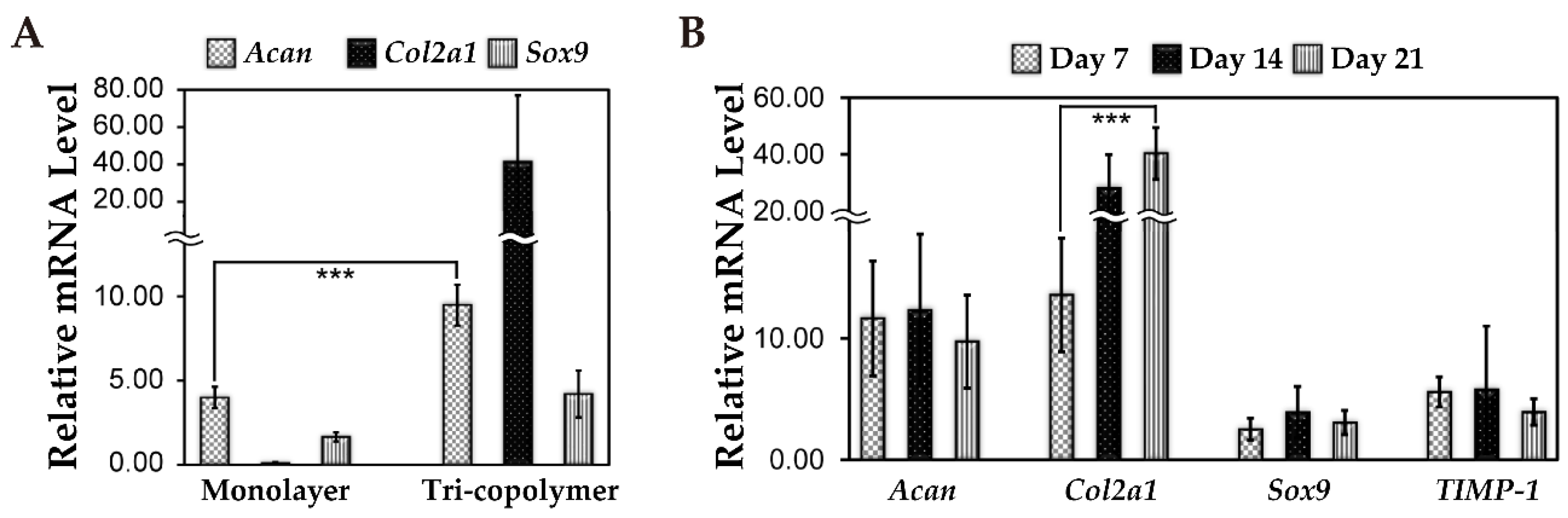
3.5. Cartilage-Related Gene Expression Under 3D Static and Dynamic Culture Conditions
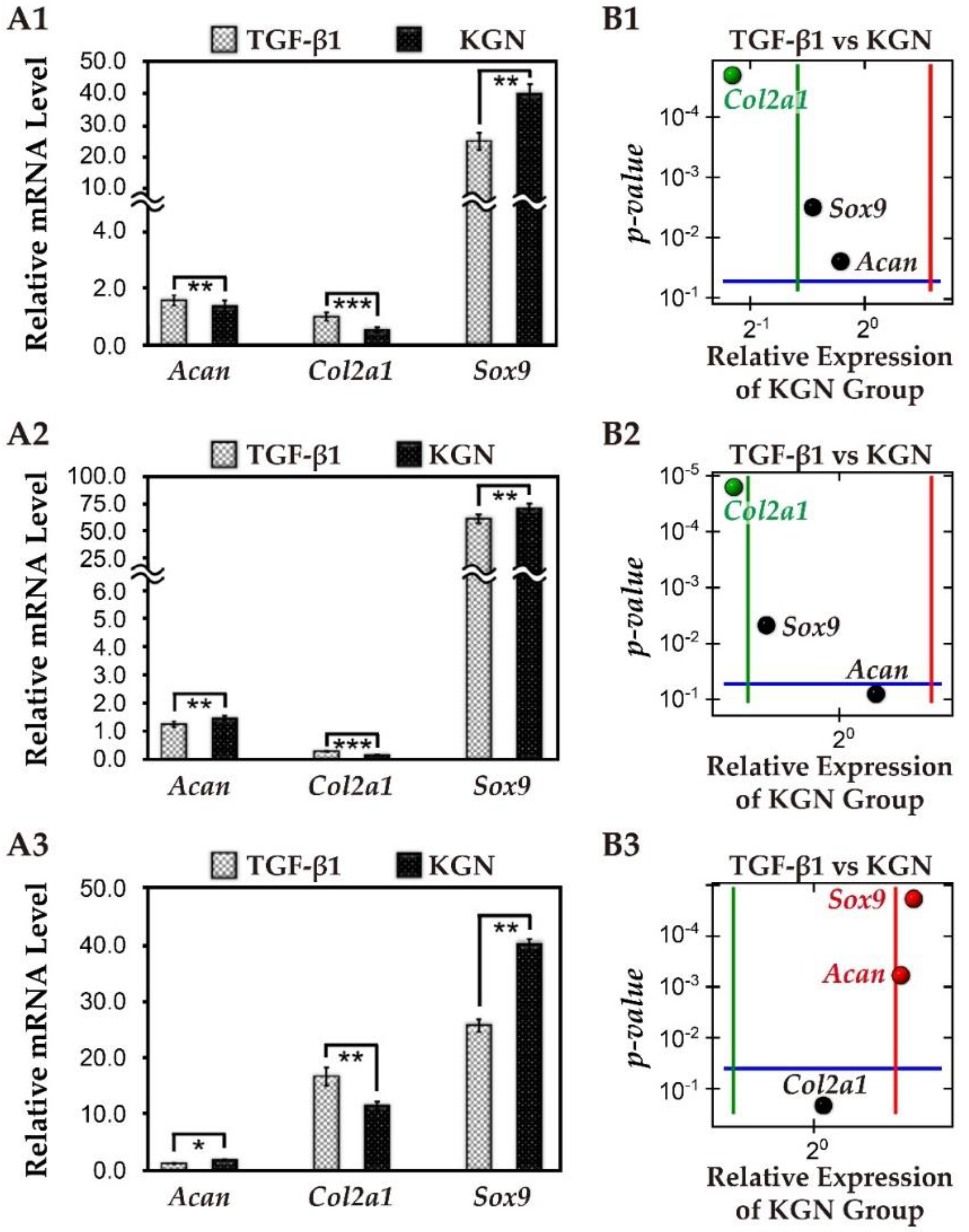
3.6. The Comparison of KGN and TGF-β1
3.7. SEM Images of Scaffold/Cell Hybrids Under Dynamic Perfusion
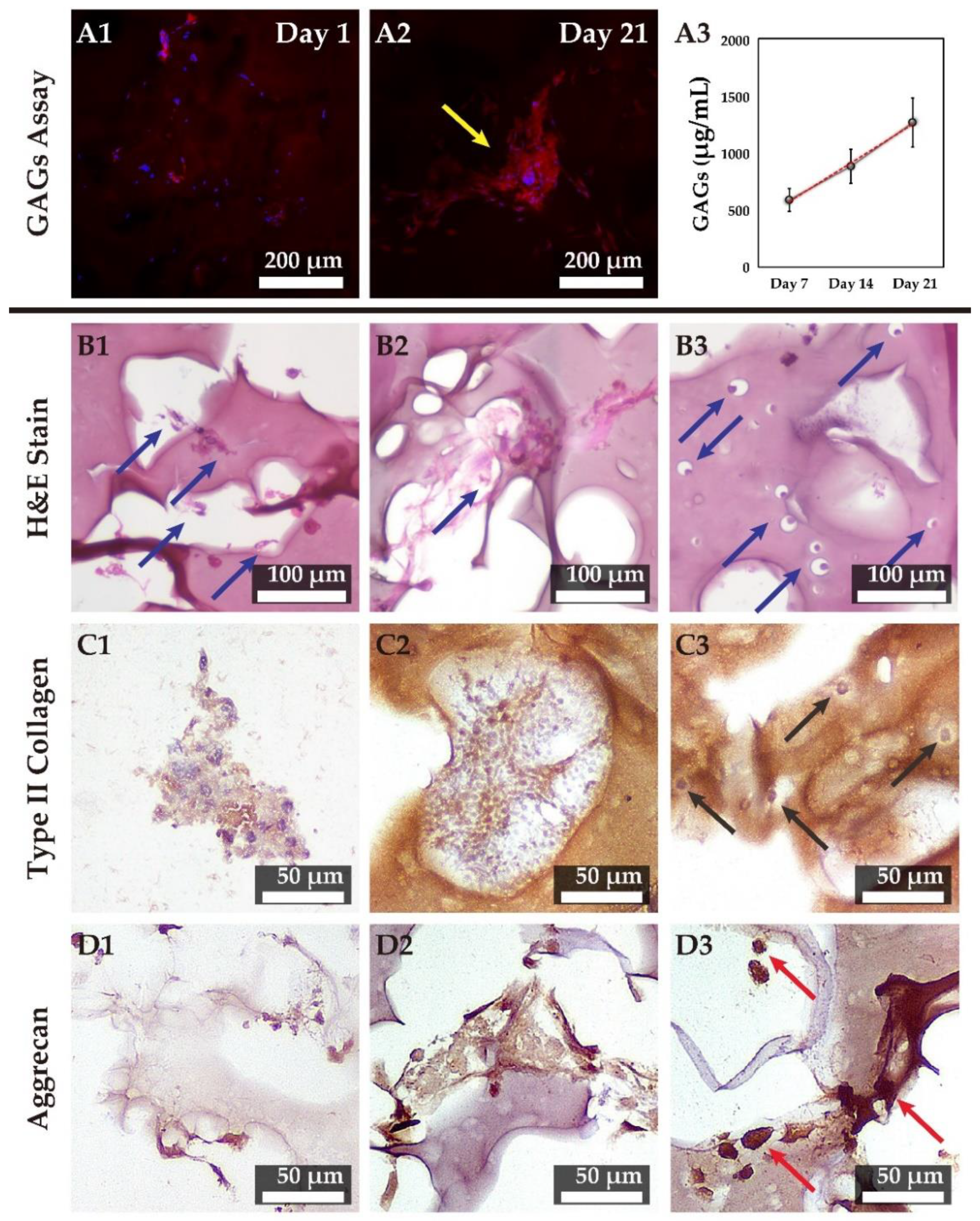
3.8. Chondrogenesis in 3D Tri-Copolymer Scaffolds and the Self-Designed Bioreactor System
3.9. Hematoxylin and Eosin Staining and IHC Examination of the Tri-Copolymer Scaffold/rMSCs Constructs
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Bora, F.W.; Miller, G. Joint physiology, cartilage metabolism, and the etiology of osteoarthritis. Hand Clin. 1987, 3, 325–336. [Google Scholar] [PubMed]
- Aichroth, P. Osteochondritis dissecans of the knee. A clinical survey. J. Bone Jt. Surg. Br. 1971, 53, 440–447. [Google Scholar] [CrossRef]
- Arden, N.K.; Perry, T.; Bannuru, R.R.; Bruyère, O.; Cooper, C.; Haugen, I.K.; Hochberg, M.C.; McAlindon, T.E.; Mobasheri, A.; Reginster, J.-Y. Non-surgical management of knee osteoarthritis: Comparison of ESCEO and OARSI 2019 guidelines. Nat. Rev. Rheumatol. 2021, 17, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Liu, H.-C.; Lin, C.-C.; Chou, C.-H.; Lin, F.-H. Gelatin-chondroitin-hyaluronan tri-copolymer scaffold for cartilage tissue engineering. Biomaterials 2003, 24, 4853–4858. [Google Scholar] [CrossRef]
- Tuli, R.; Li, W.-J.; Tuan, R.S. Current state of cartilage tissue engineering. Arthritis Res. Ther. 2003, 5, 235–238. [Google Scholar] [CrossRef]
- Eftekhari, A.; Dizaj, S.M.; Sharifi, S.; Salatin, S.; Saadat, Y.R.; Vahed, S.Z.; Samiei, M.; Ardalan, M.; Rameshrad, M.; Ahmadian, E.; et al. The Use of Nanomaterials in Tissue Engineering for Cartilage Regeneration; Current Approaches and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 536. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.M.; Barry, F.; Murphy, M.; Mahon, B.P. Mesenchymal stem cells avoid allogeneic rejection. J. Inflamm. 2005, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Goyal, D. The Classifications of the Chondral Lesions. In The Illustrative Book of Cartilage Repair; Goyal, D.R., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 43–56. [Google Scholar]
- Caplan, A.I. Adult mesenchymal stem cells for tissue engineering versus regenerative medicine. J. Cell. Physiol. 2007, 213, 341–347. [Google Scholar] [CrossRef]
- Johnson, K.A.; Zhu, S.; Tremblay, M.S.; Payette, J.N.; Wang, J.; Bouchez, L.C.; Meeusen, S.; Althage, A.; Cho, C.Y.; Wu, X.; et al. A Stem Cell-Based Approach to Cartilage Repair. Science 2012, 336, 717–721. [Google Scholar] [CrossRef] [Green Version]
- Lepage, S.I.; Robson, N.; Gilmore, H.; Davis, O.; Hooper, A.; John, S.S.; Kamesan, V.; Gelis, P.; Carvajal, D.; Hurtig, M.; et al. Beyond Cartilage Repair: The Role of the Osteochondral Unit in Joint Health and Disease. Tissue Eng. Part B Rev. 2019, 25, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Park, J.S.; Yeom, J.; Selvapalam, N.; Park, K.M.; Oh, K.; Yang, J.-A.; Park, K.H.; Hahn, S.K.; Kim, K. 3D tissue engi-neered supramolecular hydrogels for controlled chondrogenesis of human mesenchymal stem cells. Biomacromolecules 2014, 15, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.L.; Ko, J.-Y.; Kim, J.E.; Im, G.-I. Intra-articular delivery of kartogenin-conjugated chitosan nano/microparticles for cartilage regeneration. Biomaterials 2014, 35, 9984–9994. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yin, Z.; Liu, Y.; Feng, S.; Liu, Y.; Lu, F.; Xu, Y.; Min, P.; Hou, M.; Li, K. Regeneration of trachea graft with cartilage support, vascularization, and epithelization. Acta Biomater. 2019, 89, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Afflerbach, A.-K.; Kiri, M.D.; Detinis, T.; Maoz, B. Mesenchymal Stem Cells as a Promising Cell Source for Integration in Novel In Vitro Models. Biomolecules 2020, 10, 1306. [Google Scholar] [CrossRef]
- Clouet, J.; Vinatier, C.; Merceron, C.; Pot-Vaucel, M.; Maugars, Y.; Weiss, P.; Grimandi, G.; Guicheux, J. From osteoarthritis treatments to future regenerative therapies for cartilage. Drug Discov. Today 2009, 14, 913–925. [Google Scholar] [CrossRef]
- Pomeraniec, L.; Benayahu, D. Mesenchymal Cell Growth and Differentiation on a New Biocomposite Material: A Promising Model for Regeneration Therapy. Biomolecules 2020, 10, 458. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ding, J.; Zhang, Z.; Yang, M.; Yu, J.; Wang, J.; Chang, F.; Zhu, X. Kartogenin-Incorporated Thermogel Supports Stem Cells for Significant Cartilage Regeneration. ACS Appl. Mater. Interfaces 2016, 8, 5148–5159. [Google Scholar] [CrossRef]
- Fujimoto, T.; Kawashima, H.; Tanaka, T.; Hirose, M.; Toyama-Sorimachi, N.; Matsuzawa, Y.; Miyasaka, M. CD44 binds a chondroitin sulfate proteoglycan, aggrecan. Int. Immunol. 2001, 13, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Gillogly, S.D.; Voight, M.; Blackburn, T. Treatment of articular cartilage defects of the knee with autologous chondrocyte im-plantation. J. Orthop. Sports Phys. Ther. 1998, 28, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Huey, D.J.; Hu, J.C.; Athanasiou, K.A. Unlike Bone, Cartilage Regeneration Remains Elusive. Science 2012, 338, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zhu, P.; Huang, H.; Zheng, Y.; Liu, J.; Feng, L.; Guo, H.; Tang, S.; Guo, R. Functionalization of Novel Theranostic Hydrogels with Kartogenin-Grafted USPIO Nanoparticles to Enhance Cartilage Regeneration. ACS Appl. Mater. Interfaces 2019, 11, 34744–34754. [Google Scholar] [CrossRef] [PubMed]
- Asgari, N.; Bagheri, F.; Eslaminejad, M.B.; Ghanian, M.H.; Sayahpour, F.A.; Ghafari, A.M. Dual functional construct containing kartogenin releasing microtissues and curcumin for cartilage regeneration. Stem Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Liu, W.; He, Y.; Huang, J.; Duan, L.; Xiong, J.; Liu, L.; Wang, D. Recent advances in kartogenin for cartilage regeneration. J. Drug Target. 2019, 27, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Liu, N.; Zhou, R.; Zhao, X.; Han, Y.; Xia, F.; Cheng, J.; Duan, M.; Qian, Q.; Wang, X.; et al. Combination Therapy Using Kartogenin-Based Chondrogenesis and Complex Polymer Scaffold for Cartilage Defect Regeneration. ACS Biomater. Sci. Eng. 2020, 6, 6276–6284. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, N.; Kundu, S.C. Chondrogenic differentiation of rat MSCs on porous scaffolds of silk fibroin/chitosan blends. Biomaterials 2012, 33, 2848–2857. [Google Scholar] [CrossRef] [PubMed]
- Kakade, M.; Liener, I.E. Determination of available lysine in proteins. Anal. Biochem. 1969, 27, 273–280. [Google Scholar] [CrossRef]
- Sashidhar, R.B.; Capoor, A.K.; Ramana, D. Quantitation of epsilon-amino group using amino acids as reference standards by trinitrobenzene sulfonic acid. A simple spectrophotometric method for the estimation of hapten to carrier protein ratio. J. Immunol. Methods 1994, 167, 121–127. [Google Scholar] [CrossRef]
- Lin, F.-H.; Chiang, T.-S.; Chiou, L.-L.; Lee, H.-S.; Lin, F.-H. 3D cell clusters combined with a bioreactor system to enhance the drug metabolism activities of C3A hepatoma cell lines. J. Mater. Chem. B 2016, 4, 7000–7008. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Ke, C.-J.; Yen, K.-C.; Hsieh, H.-C.; Sun, J.-S.; Lin, F.-H. 3D Porous Calcium-Alginate Scaffolds Cell Culture System Improved Human Osteoblast Cell Clusters for Cell Therapy. Theranostics 2015, 5, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Tseng, K.-Y.; Lai, Y.-L.; Chen, Y.-S.; Lin, F.-H.; Lin, S. Overexpression of Insulin-Like Growth Factor 1 Enhanced the Osteogenic Capability of Aging Bone Marrow Mesenchymal Stem Cells. Theranostics 2017, 7, 1598–1611. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Tseng, K.-Y.; Wong, Z.-H.; Chen, Y.-P.; Chen, T.-Y.; Chen, H.-Y.; Chen, Z.-Y.; Lin, F.-H.; Wu, H.-M.; Lin, S. Co-operative impact of thiazolidinedione and fatty acid synthase on human osteogenesis. Aging 2019, 11, 2327. [Google Scholar] [CrossRef] [PubMed]
- Grassel, S.; Ahmed, N.; Gottl, C.; Grifka, J. Gene and protein expression profile of naive and osteo-chondrogenically differenti-ated rat bone marrow-derived mesenchymal progenitor cells. Int. J. Mol. Med. 2009, 23, 745. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, A.; Wang, Z.; Takahashi, S.; Tokuda, T.; Miura, N.; Hasegawa, J. Upregulation of mRNA of retinoid binding protein and fatty acid binding protein by cholesterol enriched-diet and effect of ginger on lipid metabolism. Life Sci. 2009, 84, 903–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tıglı, R.S.; Ghosh, S.; Laha, M.M.; Shevde, N.K.; Dahéron, L.; Gimble, J.; Gümüşderelioglu, M.; Kaplan, D.L. Comparative chondrogenesis of human cell sources in 3D scaffolds. J. Tissue Eng. Regen. Med. 2009, 3, 348–360. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Brown, W.E.; Lee, C.A.; Wang, D.; Paschos, N.; Hu, J.C.; Athanasiou, K.A. Surgical and tissue engineering strategies for articular cartilage and meniscus repair. Nat. Rev. Rheumatol. 2019, 15, 550–570. [Google Scholar] [CrossRef]
- Martin, I.; Galipeau, J.; Kessler, C.; Le Blanc, K.; Dazzi, F. Challenges for mesenchymal stromal cell therapies. Sci. Transl. Med. 2019, 11, eaat2189. [Google Scholar] [CrossRef]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2005, 97, 33–44. [Google Scholar] [CrossRef]
- Jing, Y.; Wang, Z.; Li, H.; Ma, C.; Feng, J.Q. Chondrogenesis Defines Future Skeletal Patterns Via Cell Transdifferentiation from Chondrocytes to Bone Cells. Curr. Osteoporos. Rep. 2020, 18, 199–209. [Google Scholar] [CrossRef]
- Maleski, M.P.; Knudson, C.B. Hyaluronan-mediated aggregation of limb bud mesenchyme and mesenchymal condensation during chondrogenesis. Exp. Cell Res. 1996, 225, 55–66. [Google Scholar] [CrossRef]
- Lu, C.-H.; Lin, K.-J.; Chiu, H.-Y.; Chen, C.-Y.; Yen, T.-C.; Hwang, S.-M.; Chang, Y.; Hu, Y.-C. Improved Chondrogenesis and Engineered Cartilage Formation from TGF-β3-Expressing Adipose-Derived Stem Cells Cultured in the Rotating-Shaft Bioreactor. Tissue Eng. Part A 2012, 18, 2114–2124. [Google Scholar] [CrossRef]
- Sechriest, V.F.; Miao, Y.J.; Niyibizi, C.; Westerhausen-Larson, A.; Matthew, H.W.; Evans, C.H.; Fu, F.H.; Suh, J.-K. GAG-augmented polysaccharide hydrogel: A novel biocompatible and biodegradable material to support chondrogenesis. J. Biomed. Mater. Res. 2000, 49, 534–541. [Google Scholar] [CrossRef]
- Armiento, A.R.; Stoddart, M.J.; Alini, M.; Eglin, D. Biomaterials for articular cartilage tissue engineering: Learning from biology. Acta Biomater. 2018, 65, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Ishizuka, S.; Knudson, C.B.; Knudson, W. Chondroprotective Effect of Kartogenin on CD44-Mediated Functions in Articular Cartilage and Chondrocytes. Cartilage 2014, 5, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Ochi, M.; Uchio, Y.; Adachi, N.; Matsusaki, M. Hyaluronic acid enhances proliferation and chondroitin sulfate synthesis in cultured chondrocytes embedded in collagen gels. J. Cell. Physiol. 1999, 179, 142–148. [Google Scholar] [CrossRef]
- Lairson, L.L.; Lyssiotis, C.A.; Zhu, S.; Schultz, P.G. Small Molecule–Based Approaches to Adult Stem Cell Therapies. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 107–125. [Google Scholar] [CrossRef] [Green Version]
- Caron, M.M.; Emans, P.J.; Surtel, D.A.; Cremers, A.; Voncken, J.W.; Welting, T.J.; Van Rhijn, L.W. Activation of NF-κB/p65 facilitates early chondrogenic differentiation during endochondral ossification. PLoS ONE 2012, 7, e33467. [Google Scholar] [CrossRef] [Green Version]
- Danisovic, L.; Varga, I.; Polak, S. Growth factors and chondrogenic differentiation of mesenchymal stem cells. Tissue Cell 2012, 44, 69–73. [Google Scholar] [CrossRef]
- Mueller, M.B.; Fischer, M.; Zellner, J.; Berner, A.; Dienstknecht, T.; Prantl, L.; Kujat, R.; Nerlich, M.; Tuan, R.S.; Angele, P. Hypertrophy in Mesenchymal Stem Cell Chondrogenesis: Effect of TGF-β Isoforms and Chondrogenic Conditioning. Cells Tissues Organs 2010, 192, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, C.G.; Karl, A.; Kerschbaum, M.; Berner, A.; Lang, S.; Schupfner, R.; Koch, M.; Angele, P.; Nerlich, M.; Mueller, M.B. TGF-β Signalling is Suppressed under Pro-Hypertrophic Conditions in MSC Chondrogenesis Due to TGF-β Receptor Down-regulation. Int. J. Stem Cells 2019, 12, 139. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers Sequences | Reference |
|---|---|---|
| Acan (NM_022190) | F-GGCCTTCCCTCTGGATTTAG | [26,33] |
| R-CCGCACTACTGTCCAAC | ||
| Col2a1 (NM_012929) | F-CCCCTGCAGTACATGCGG | [33] |
| R-CTCGACGTCATGCTGTCTCAAG | ||
| Sox9 (XM_003750950.1) | F-CTGAAGGGCTACGACTGGAC | [26,33] |
| R-TACTGGTCTGCCAGCTTCCT | ||
| TIMP-1 (NM_053819) | F-TTTCCGTTCCTTAAACGGCC | [33] |
| R-GATTCGACGCTGTGGGAAAT | ||
| β-Actin (NM_031144) | F-GTAGCCATCCAGGCTGTGTT | [34] |
| R-CCCTCATAGATGGGCAGAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-Y.; Li, C.; Ke, C.-J.; Sun, J.-S.; Lin, F.-H. Kartogenin Enhances Chondrogenic Differentiation of MSCs in 3D Tri-Copolymer Scaffolds and the Self-Designed Bioreactor System. Biomolecules 2021, 11, 115. https://doi.org/10.3390/biom11010115
Chen C-Y, Li C, Ke C-J, Sun J-S, Lin F-H. Kartogenin Enhances Chondrogenic Differentiation of MSCs in 3D Tri-Copolymer Scaffolds and the Self-Designed Bioreactor System. Biomolecules. 2021; 11(1):115. https://doi.org/10.3390/biom11010115
Chicago/Turabian StyleChen, Ching-Yun, Chunching Li, Cherng-Jyh Ke, Jui-Sheng Sun, and Feng-Huei Lin. 2021. "Kartogenin Enhances Chondrogenic Differentiation of MSCs in 3D Tri-Copolymer Scaffolds and the Self-Designed Bioreactor System" Biomolecules 11, no. 1: 115. https://doi.org/10.3390/biom11010115
APA StyleChen, C. -Y., Li, C., Ke, C. -J., Sun, J. -S., & Lin, F. -H. (2021). Kartogenin Enhances Chondrogenic Differentiation of MSCs in 3D Tri-Copolymer Scaffolds and the Self-Designed Bioreactor System. Biomolecules, 11(1), 115. https://doi.org/10.3390/biom11010115