
Sterol Extraction from Isolated Plant Plasma Membrane Vesicles Affects H+-ATPase Activity and H+-Transport

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasma Membrane Isolation
2.2. Analysis of Membrane Lipids
2.3. Sterol Extraction
2.4. Vesicle Size
2.5. ATP Hydrolysis
2.6. ATP-Dependent ∆pH and ∆Ψ Generation
2.7. Passive H+-Permeability
2.8. Detergent-Resistant Plasma Membrane Fractions
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
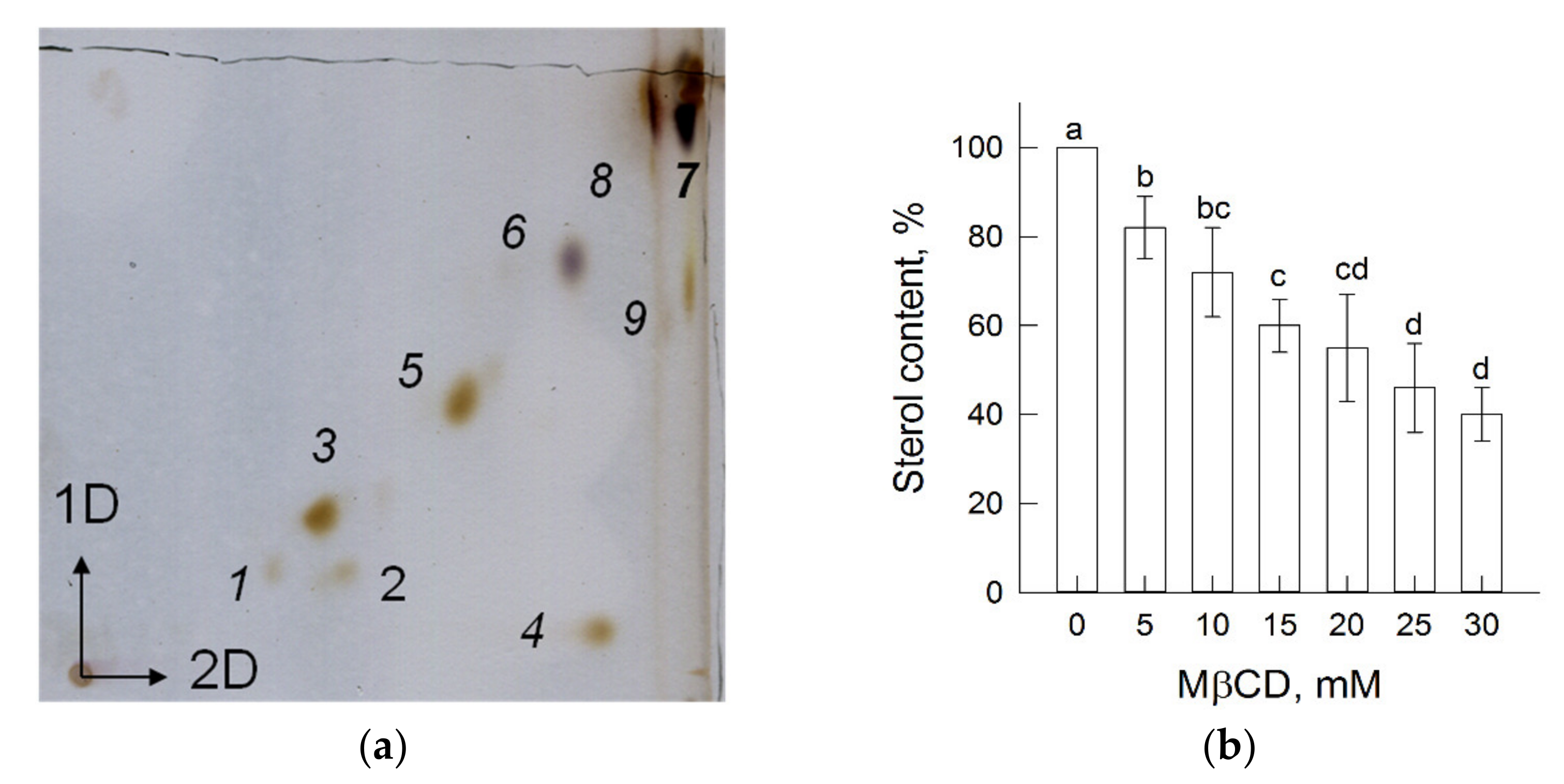
3.1. Plasma Membrane Lipids and Sterol Extraction
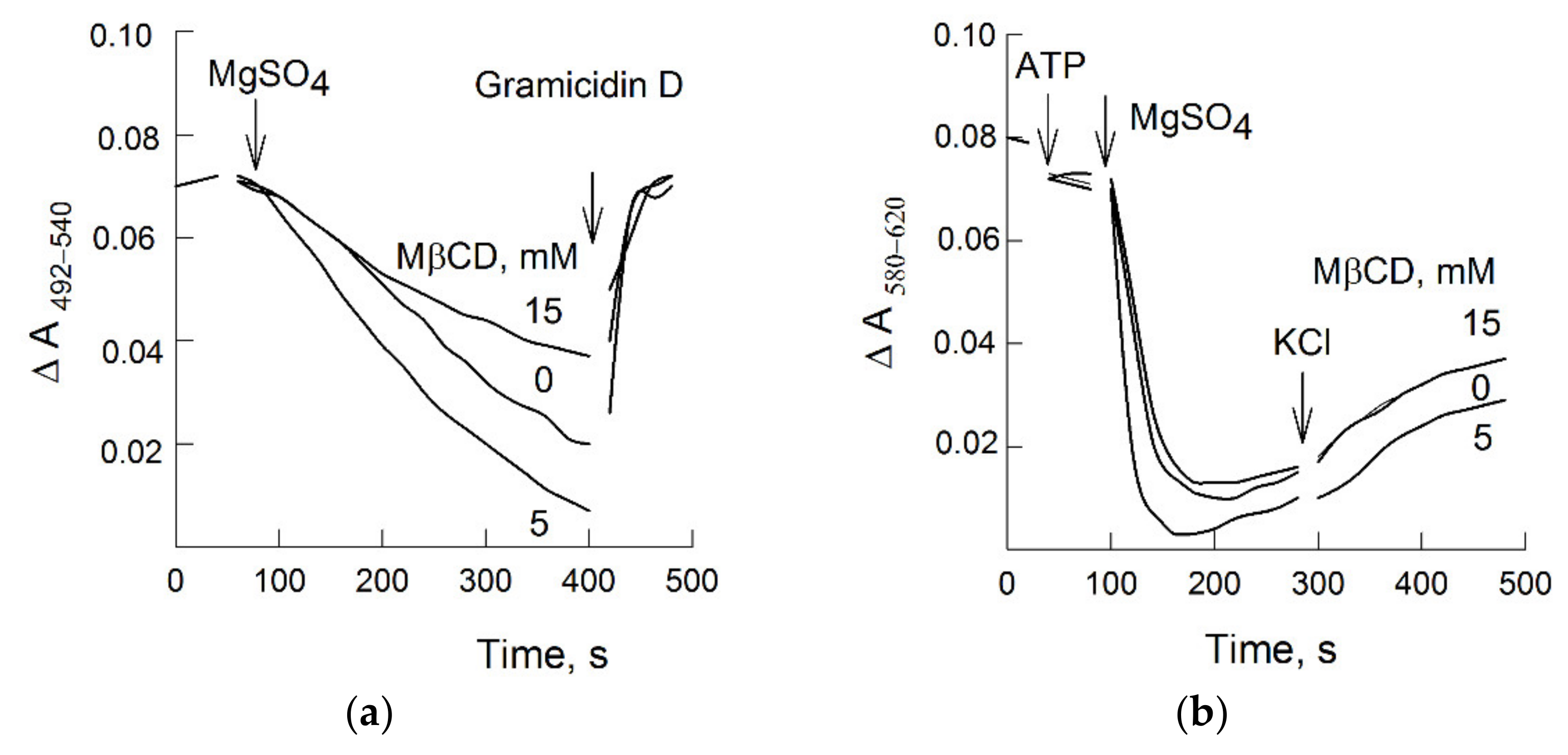
3.2. MβCD and H+-Pumping
3.3. ATP Hydrolysis
3.4. MβCD and Plasma Membrane H+-Leakage
3.5. Sterol Extraction and Vesicle Size
3.6. Sterols and ∆Ψ Generation
3.7. ATP Hydrolysis in the Presence of Dodecyl Maltoside (DDM)
3.8. H+-ATPase in Detergent-Resistant Fractions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Powerhouses for nutrient uptake. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sondergaard, T.E.; Schulz, A.; Palmgren, M.G. Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase. Plant Physiol. 2004, 136, 2475–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haruta, M.; Sussman, M.R. The effect of a genetically reduced plasma membrane protonmotive force on vegetative growth of Arabidopsis. Plant Physiol. 2012, 158, 1158–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühlbrandt, W. Biology, structure and mechanism of P-type ATPases. Nat. Rev. Mol. Cell Biol. 2004, 5, 282–295. [Google Scholar] [CrossRef]
- Dyla, M.; Kjærgaard, M.; Poulsen, H.; Nissen, P. Structure and mechanism of P-Type ATPase ion pumps. Annu. Rev. Biochem. 2020, 89, 583–603. [Google Scholar] [CrossRef]
- Buch-Pedersen, M.J.; Palmgren, M.G. Conserved Asp684 in transmembrane segment M6 of the plant plasma membrane P-type proton pump AHA2 is a molecular determinant of proton translocation. J. Biol. Chem. 2003, 278, 17845–17851. [Google Scholar] [CrossRef] [Green Version]
- Ekberg, K.; Wielandt, A.G.; Buch-Pedersen, M.J.; Palmgren, M.G. A conserved asparagine in a P-type proton pump is required for efficient gating of protons. J. Biol. Chem. 2013, 288, 9610–9618. [Google Scholar] [CrossRef] [Green Version]
- Haruta, M.; Gray, W.M.; Sussman, M.R. Regulation of the plasma membrane proton pump (H+-ATPase) by phosphorylation. Curr. Opin. Plant Biol. 2015, 28, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma membrane H+-ATPase regulation in the center of plant physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Venema, K.; Palmgren, M.G. Metabolic modulation of transport coupling ratio in yeast plasma membrane H+ATPase. J. Biol. Chem. 1995, 270, 19659–19667. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, J.T.; Kanashova, T.; Dittmar, G.; Palmgren, M. Isolation of native plasma membrane H+-ATPase (Pma1p) in both the active and basal activation states. FEBS Open Bio 2018, 8, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Baunsgaard, L.; Fuglsang, A.T.; Jahn, T.; Korthout, H.A.; de Boer, A.H.; Palmgren, M.G. The 14-3-3 proteins associate with the plant plasma membrane H+-ATPase to generate a fusicoccin binding complex and a fusicoccin responsive system. Plant J. 1998, 13, 661–671. [Google Scholar] [CrossRef]
- Olivari, C.; Meanti, C.; De Michelis, M.I.; Rasi-Caldogno, F. Fusicoccin binding to its plasma membrane receptor and the activation of the plasma membrane H+-ATPase. IV. Fusicoccin induces the association between the plasma membrane H+-ATPase and the fusicoccin receptor. Plant Physiol. 1998, 116, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Palmgren, M.G.; Larsson, C.; Sommarin, M. Proteolytic activation of the plant plasma membrane H+-ATPase by removal of a terminal segment. J. Biol. Chem. 1990, 265, 13423–13426. [Google Scholar] [CrossRef]
- Wielandt, A.G.; Pedersen, J.T.; Falhof, J.; Kemmer, G.C.; Lund, A.; Ekberg, K.; Fuglsang, A.T.; Pomorski, T.G.; Buch-Pedersen, M.G.; Palmgren, M. Specific activation of the plant P-type plasma membrane H+-ATPase by lysophospholipids depends on the autoinhibitory N- and C-terminal domains. J. Biol. Chem. 2015, 290, 16281–16291. [Google Scholar] [CrossRef] [Green Version]
- Johansson, F.; Olbe, M.; Sommarin, M.; Larsson, C. Brij 58, a polyoxyethylene acyl ether, creates membrane vesicles of uniform sidedness. A new tool to obtain inside-out (cytoplasmic side-out) plasma membrane vesicles. Plant J. 1995, 7, 165–173. [Google Scholar] [CrossRef]
- Sandstrom, R.P.; Cleland, R.E. Selective delipidation of the plasma membrane by surfactants: Enrichment of sterols and activation of ATPase. Plant Physiol. 1989, 90, 1524–1531. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.G. Lipid-protein interactions in biological membranes: A structural perspective. Biochim. Biophys. Acta 2003, 1612, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.G. How lipids affect the activities of integral membrane proteins. Biochim. Biophys. Acta 2004, 1666, 62–87. [Google Scholar] [CrossRef] [Green Version]
- Kasamo, K. Regulation of plasma membrane H+-ATPase activity by the membrane environment. J. Plant Res. 2003, 116, 517–523. [Google Scholar] [CrossRef]
- Habeck, M.; Haviv, H.; Katz, A.; Kapri-Pardes, E.; Ayciriex, S.; Shevchenko, A.; Ogawa, H.; Toyoshima, C.; Karlish, S.J.D. Stimulation, inhibition, or stabilization of Na,K-ATPase caused by specific lipid interactions at distinct sites. J. Biol. Chem. 2015, 290, 4829–4842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Cedillo, F.; González-Solís, A.; Gutiérrez-Angoa, L.; Cano-Ramírez, D.L.; Gavilanes-Ruiz, M. Plant lipid environment and membrane enzymes: The case of the plasma membrane H+-ATPase. Plant Cell Rep. 2015, 34, 617–629. [Google Scholar] [CrossRef]
- Hossain, K.R.; Clarke, R.J. General and specific interactions of the phospholipid bilayer with P-type ATPases. Biophys. Rev. 2019, 11, 353–364. [Google Scholar] [CrossRef]
- Malinsky, J.; Opekarová, M.; Grossmann, G.; Tanner, W. Membrane microdomains, rafts, and detergent-resistant membranes in plants and fungi. Annu. Rev. Plant Biol. 2013, 64, 501–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuck, S.; Honsho, M.; Ekroos, K.; Shevchenko, A.; Simons, K. Resistance of cell membranes to different detergents. Proc. Natl. Acad. Sci. USA 2003, 100, 5795–5800. [Google Scholar] [CrossRef] [Green Version]
- Mongrand, S.; Morel, J.; Laroche, J.; Claverol, S.; Carde, J.P.; Hartmann, M.A.; Bonneu, M.; Simon-Plas, F.; Lessire, R.; Bessoule, J.J. Lipid rafts in higher plant cells: Purification and characterization of Triton X-100-insoluble microdomains from tobacco plasma membrane. J. Biol. Chem. 2004, 279, 36277–36286. [Google Scholar] [CrossRef] [Green Version]
- Borner, G.H.H.; Sherrier, D.J.; Weimar, T.; Michaelson, L.V.; Hawkins, N.D.; MacAskill, A.; Napier, J.A.; Beale, M.H.; Lilley, K.S.; Dupree, P. Analysis of Detergent-Resistant Membranes in Arabidopsis. Evidence for Plasma Membrane Lipid Rafts. Plant Physiol. 2005, 137, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Laloi, M.; Perret, A.M.; Chatre, L.; Melser, S.; Cantrel, C.; Vaultier, M.N.; Zachowski, A.; Bathany, K.; Schmitter, J.M.; Vallet, M.; et al. Insights into the role of specific lipids in the formation and delivery of lipid microdomains to the plasma membrane of plant cells. Plant Physiol. 2007, 143, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Minami, A.; Fujiwara, M.; Furuto, A.; Fukao, Y.; Yamashita, T.; Kamo, M.; Kawamura, Y.; Uemura, M. Alterations in detergent-resistant plasma membrane microdomains in Arabidopsis thaliana during cold acclimation. Plant Cell Physiol. 2009, 50, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Schrick, K.; Cordova, C.; Li, G.; Murray, L.; Fujioka, S. A dynamic role for sterols in embryogenesis of Pisum sativum. Phytochemistry 2011, 72, 465–475. [Google Scholar] [CrossRef]
- Ferrer, A.; Altabella, T.; Arró, M.; Boronat, A. Emerging roles for conjugated sterols in plants. Prog. Lipid Res. 2017, 67, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hemmerlin, A.; Harwood, J.L.; Bach, T.J. A raison d’être for two distinct pathways in the early steps of plant isoprenoid biosynthesis? Prog. Lipid Res. 2012, 51, 95–148. [Google Scholar] [CrossRef] [PubMed]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant sterols: Diversity, biosynthesis, and physiological functions. Biochemistry 2016, 81, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A.; Szakiel, A. The role of sterols in plant response to abiotic stress. Phytochem. Rev. 2020, 19, 1525–1538. [Google Scholar] [CrossRef]
- Schaller, H. The role of sterols in plant growth and development. Prog. Lipid Res. 2003, 42, 163–175. [Google Scholar] [CrossRef]
- Nakamoto, M.; Schmit, A.C.; Heintz, D.; Schaller, H.; Ohta, D. Diversification of sterol methyltransferase enzymes in plants and a role for β-sitosterol in oriented cell plate formation and polarized growth. Plant J. 2015, 84, 860–874. [Google Scholar] [CrossRef]
- Cornelius, F.; Habeck, M.; Kanai, R.; Toyoshima, C.; Karlish, S.J. General and specific lipid-protein interactions in Na,K-ATPase. Biochim. Biophys. Acta 2015, 1848, 1729–1743. [Google Scholar] [CrossRef] [Green Version]
- Grandmougin-Ferjani, A.; Schuler-Muller, I.; Hartmann, M.A. Sterol modulation of the plasma membrane H+-ATPase activity from corn roots reconstituted into soybean lipids. Plant Physiol. 1997, 113, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Rossard, S.; Roblin, G.; Atanassova, R. Ergosterol triggers characteristic elicitation steps in Beta vulgaris leaf tissues. J. Exp. Bot. 2010, 61, 1807–1816. [Google Scholar] [CrossRef]
- Belugin, B.V.; Zhestkova, I.M.; Trofimova, M.S. Affinity of PIP-aquaporins to sterol-enriched domains in plasma membrane of the cells of etiolated pea seedlings. Biochem. Moscow Suppl. Ser. A 2011, 5, 56–63. [Google Scholar] [CrossRef]
- Larsson, C.; Sommarin, M.; Widell, S. Isolation of highly purified plasma membranes and the separation of inside-out and right-side-out vesicles. Meth. Enzymol. 1994, 228, 451–469. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 379, 911–917. [Google Scholar] [CrossRef]
- Yoshida, S.; Uemura, M. Lipid composition of plasma membranes and tonoplasts isolated from etiolated seedlings of mung bean (Vigna radiata L.). Plant Physiol. 1986, 82, 807–812. [Google Scholar] [CrossRef] [Green Version]
- Palmgren, M.G. An H+-ATPase assay: Proton pumping and ATPase activity determined simultaneously in the same sample. Plant Physiol. 1990, 94, 882–886. [Google Scholar] [CrossRef] [Green Version]
- Ershov, P.V.; Reshetova, O.S.; Trofimova, M.S.; Babakov, A.V. Activity of ion transporters and salt tolerance in barley. Russ. J. Plant Physiol. 2005, 52, 765–773. [Google Scholar] [CrossRef]
- Apell, H.J.; Bersch, B. Oxonol VI as an optical indicator for membrane potentials in lipid vesicles. Biochim. Biophys. Acta 1987, 903, 480–494. [Google Scholar] [CrossRef] [Green Version]
- Clerc, S.; Barenholz, Y. A quantitative model for using acridine orange as a transmembrane pH gradient probe. Anal. Biochem. 1998, 259, 104–111. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [Green Version]
- Ryzhakov, A.; Do Thi, T.; Stappaerts, J.; Bertoletti, L.; Kimpe, K.; Sá Couto, A.R.; Saokham, P.; Van den Mooter, G.; Augustijns, P.; Somsen, G.W.; et al. Self-Assembly of Cyclodextrins and Their Complexes in Aqueous Solutions. J. Pharm. Sci. 2016, 105, 2556–2569. [Google Scholar] [CrossRef] [Green Version]
- Roche, Y.; Gerbeau-Pissot, P.; Buhot, B.; Thomas, D.; Bonneau, L.; Gresti, J.; Mongrand, S.; Perrier-Cornet, J.M.; Simon-Plas, F. Depletion of phytosterols from the plant plasma membrane provides evidence for disruption of lipid rafts. FASEB J. 2008, 22, 3980–3991. [Google Scholar] [CrossRef] [Green Version]
- Reisinger, V.; Eichacker, L.A. Solubilization of membrane protein complexes for blue native PAGE. J. Proteom. 2008, 71, 277–283. [Google Scholar] [CrossRef]
- Haines, T.H. Do sterols reduce proton and sodium leaks through lipid bilayers? Prog. Lipid Res. 2001, 40, 299–324. [Google Scholar] [CrossRef]
- Gensure, R.H.; Zeidel, M.L.; Hill, W.G. Lipid raft components cholesterol and sphingomyelin increase H+/OH− permeability of phosphatidylcholine membranes. Biochem. J. 2006, 398, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Dufourc, E.J. Sterols and membrane dynamics. J. Chem. Biol. 2008, 1, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Mlayeh, L.; Chatkaew, S.; Léonetti, M.; Homblé, F. Modulation of plant mitochondrial VDAC by phytosterols. Biophys. J. 2010, 99, 2097–2106. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Lev, B.; Hossain, K.R.; Gorman, A.; Diaz, D.; Pham, T.H.N.; Cornelius, F.; Allen, T.W.; Clarke, R.J. Cholesterol depletion inhibits N+,K+-ATPase activity in a near-native membrane environment. J. Biol. Chem. 2019, 294, 5956–5969. [Google Scholar] [CrossRef]
- Kierszniowska, S.; Seiwert, B.; Schulze, W.X. Definition of Arabidopsis sterol-rich membrane microdomains by differential treatment with methyl-beta-cyclodextrin and quantitative proteomics. Mol. Cell Proteom. 2009, 8, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, S.A.; Gunther, G.; Tricerri, M.A.; Gratton, E. Methyl-β-cyclodextrins preferentially remove cholesterol from the liquid disordered phase in giant unilamellar vesicles. J. Membr. Biol. 2011, 241, 1–10. [Google Scholar] [CrossRef] [Green Version]
- López, C.; de Vries, A.; Marrink, S. Computational microscopy of cyclodextrin mediated cholesterol extraction from lipid model membranes. Sci. Rep. 2013, 3, 2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, P.; Hartmann, M.A.; Perret, A.M.; Sturbois-Balcerzak, B.; Cassagne, C. Transport of sterols to the plasma membrane of leek seedlings. Plant Physiol. 1998, 117, 931–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.; Gibbs, H.C.; Yeh, A.T.; Griffing, L.R. The Sterol Trafficking Pathway in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 616631. [Google Scholar] [CrossRef] [PubMed]
- Mikes, V.; Milat, M.L.; Ponchet, M.; Ricci, P.; Blein, J.P. The fungal elicitor cryptogein is a sterol carrier protein. FEBS Lett. 1997, 416, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Li, R.L.; Zhang, L.; Wang, Q.L.; Niehaus, K.; Baluska, F.; Samaj, J.; Lin, J.X. Lipid microdomain polarization is required for NADPH oxidase-dependent ROS signaling in Picea meyeri pollen tube tip growth. Plant J. 2009, 60, 303–313. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MβCD | KM | Vmax | H+-Pumping | H+-Permeability |
|---|---|---|---|---|
| mM | mM | µmol ATP mg−1 Protein min−1 | ∆A 103 min−1 | ∆A 103 min−1 |
| 0 | 0.220 ± 0.07 | 0.82 ± 0.01 | 8 ± 3 | 2 ± 1 |
| 2 | 0.198 ± 0.05 | 0.83 ± 0.03 | 25 ± 8 * | 7 ± 3 * |
| 5 | 0.188 ± 0.04 | 1.18 ± 0.04 | 21 ± 6 * | 11 ± 5 * |
| 10 | 0.204 ± 0.08 | 1.32 ± 0.05 * | 14 ± 4 | 14 ± 5 * |
| 15 | 0.192 ± 0.07 | 0.70 ± 0.02 | 8 ± 4 | n.d. |
| MβCD | Sterol Content | KM | Vmax | ||
|---|---|---|---|---|---|
| mM | μg mg−1 of Protein | mM | µmol ATP mg−1 Protein min−1 | ||
| −DDM | +DDM | −DDM | +DDM | ||
| 0 | 85.6 ± 8.6 | 0.196 ± 0.03 | 0.201 ± 0.02 | 0.73 ± 0.02 | 1.19 ± 0.03 * |
| 5 | 65.5 ± 9.4 | 0.200 ± 0.04 | 0.219 ± 0.03 | 1.21 ± 0.03 * | 1.27 ± 0.05 * |
| 15 | 38.4 ± 8.2 | 0.215 ± 0.03 | 0.197 ± 0.03 | 0.93 ± 0.03 | 1.17 ± 0.03 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lapshin, N.K.; Piotrovskii, M.S.; Trofimova, M.S. Sterol Extraction from Isolated Plant Plasma Membrane Vesicles Affects H+-ATPase Activity and H+-Transport. Biomolecules 2021, 11, 1891. https://doi.org/10.3390/biom11121891
Lapshin NK, Piotrovskii MS, Trofimova MS. Sterol Extraction from Isolated Plant Plasma Membrane Vesicles Affects H+-ATPase Activity and H+-Transport. Biomolecules. 2021; 11(12):1891. https://doi.org/10.3390/biom11121891
Chicago/Turabian StyleLapshin, Nikita K., Michail S. Piotrovskii, and Marina S. Trofimova. 2021. "Sterol Extraction from Isolated Plant Plasma Membrane Vesicles Affects H+-ATPase Activity and H+-Transport" Biomolecules 11, no. 12: 1891. https://doi.org/10.3390/biom11121891
APA StyleLapshin, N. K., Piotrovskii, M. S., & Trofimova, M. S. (2021). Sterol Extraction from Isolated Plant Plasma Membrane Vesicles Affects H+-ATPase Activity and H+-Transport. Biomolecules, 11(12), 1891. https://doi.org/10.3390/biom11121891