
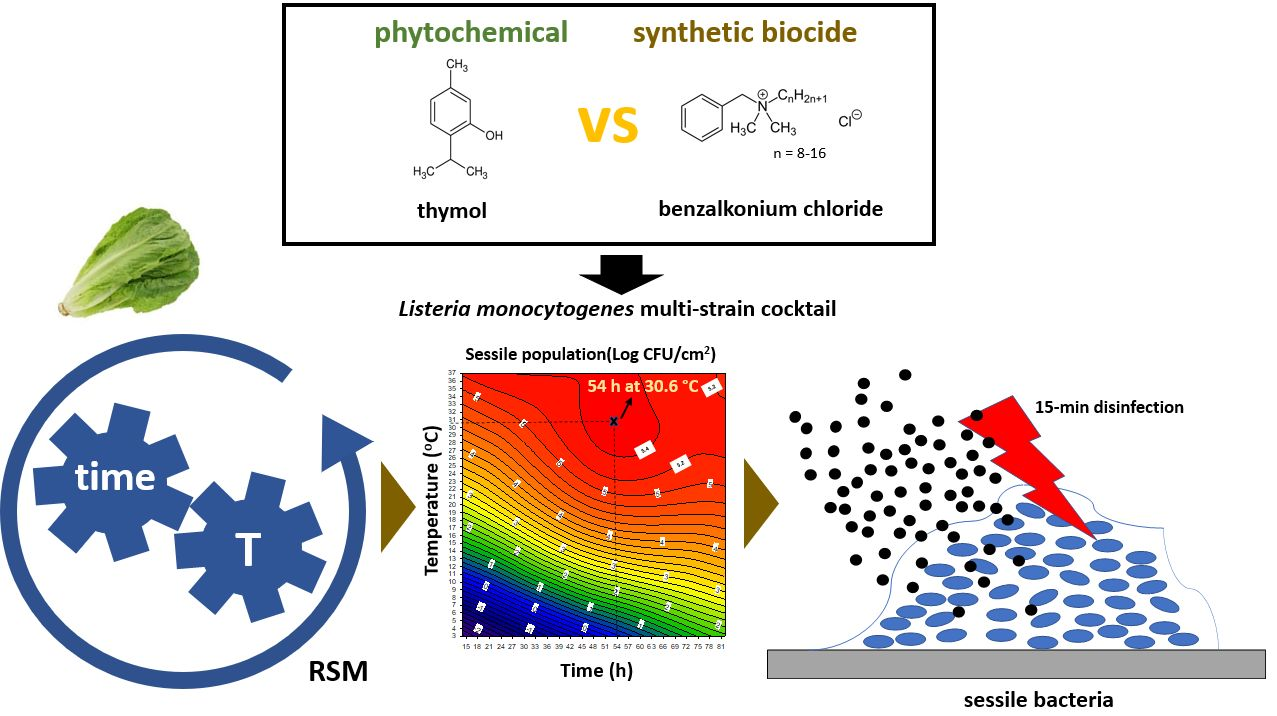
Advanced Killing Potential of Thymol against a Time and Temperature Optimized Attached Listeria monocytogenes Population in Lettuce Broth



Abstract
:
1. Introduction
2. Materials and Methods
2.1. L. monocytogenes Strains and Preparation of Their Mixed Working Suspension
2.2. Preparation of the Lettuce Broth
2.3. Experimental Design to Study the Combined Influence of Time and Temperature on Attachment
2.4. Disinfectants and Other Chemicals
2.5. Determination of the Minimum Inhibitory and Bactericidal Concentrations (MICs, MBCs)
2.6. Attachment of L. monocytogenes to SS under Various Time and Temperature Combinations and Quantification of the Sessile and Planktonic Populations
2.7. Disinfection of the Mixed Sessile Community and Calculation of the Log Reductions
2.8. Recovery of L. monocytogenes Colonies and DNA Extraction
2.9. Discrimination of L. monocytogenes Strains through Rep-PCR
2.10. Statistics and Graphics
3. Results
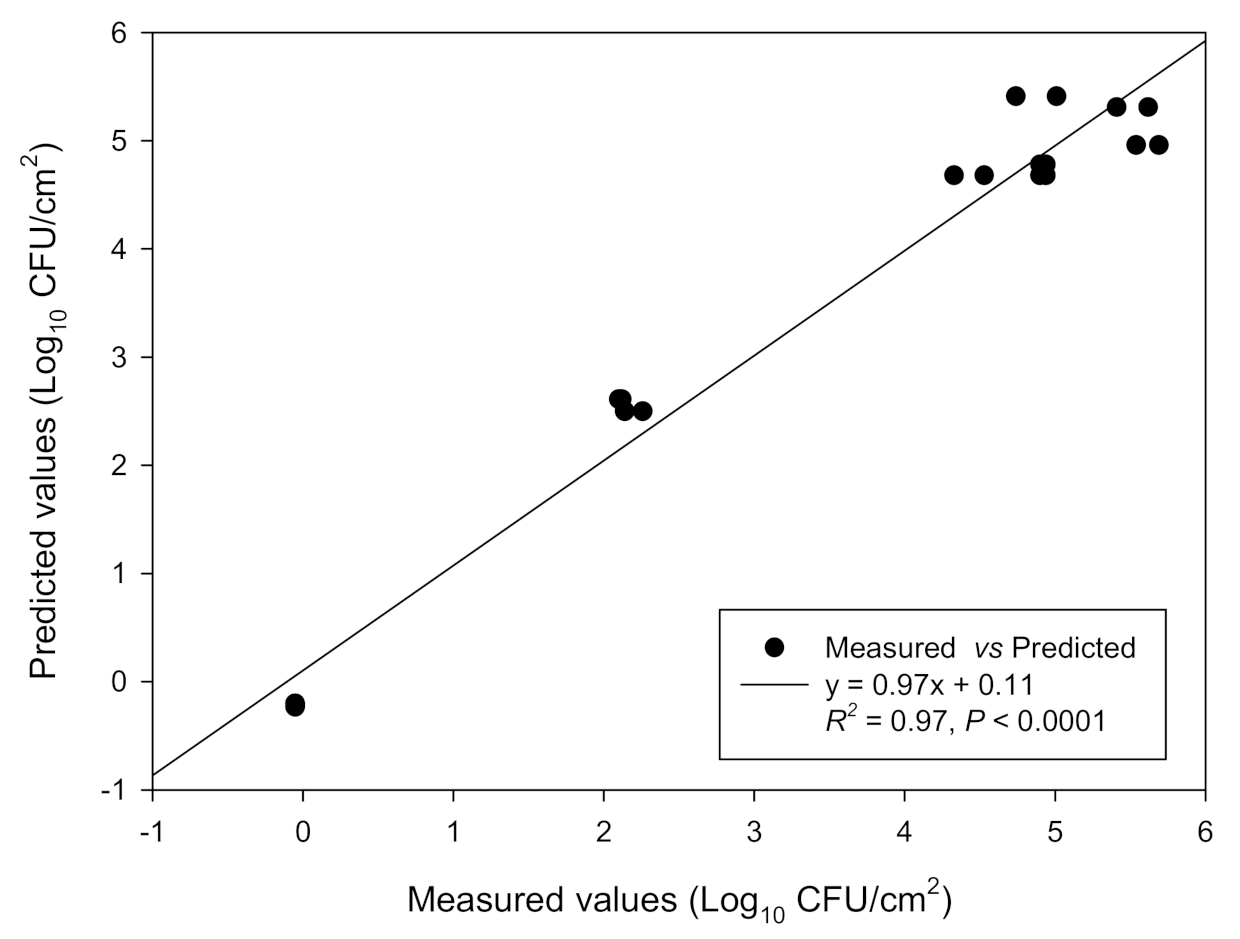
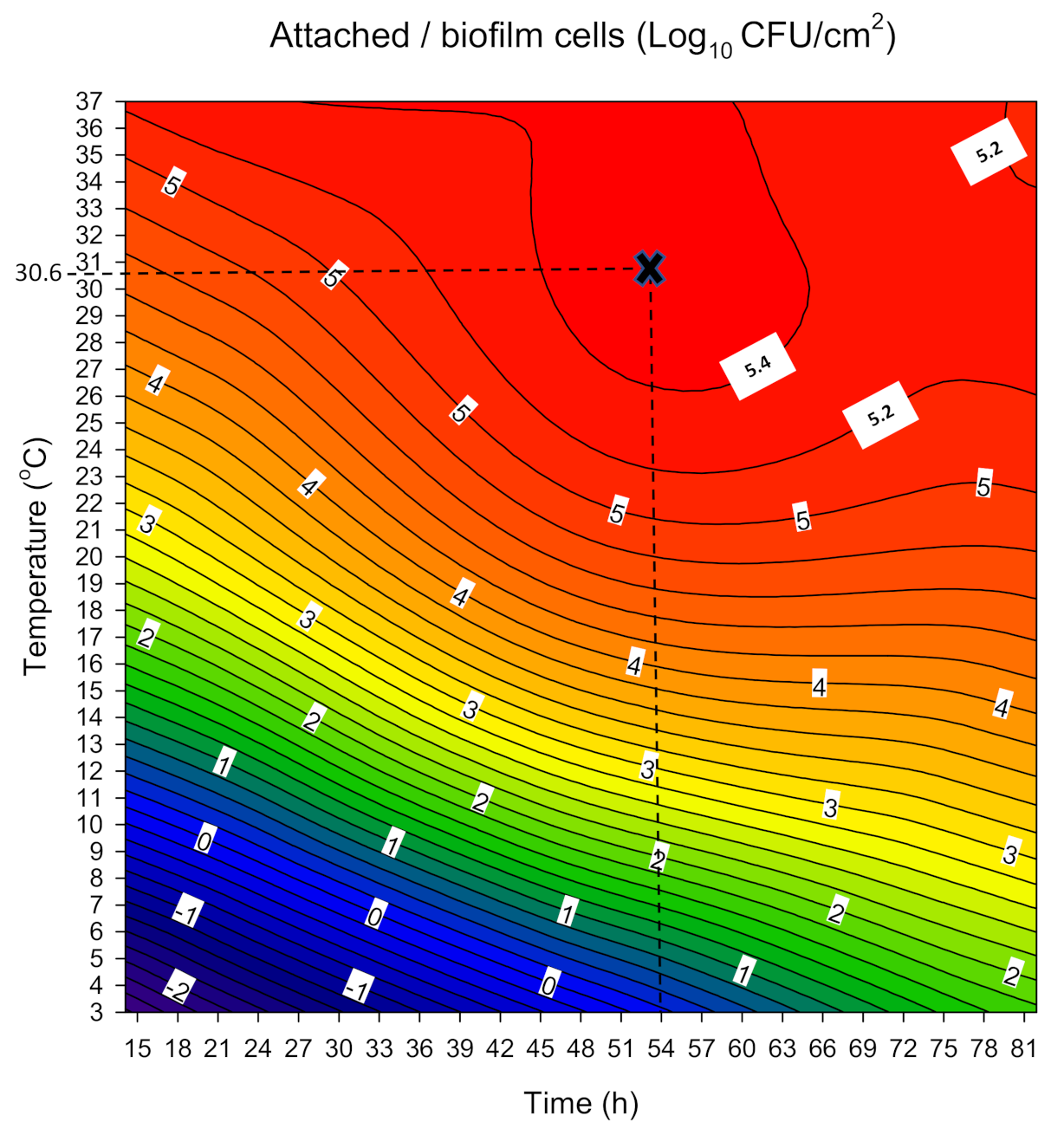
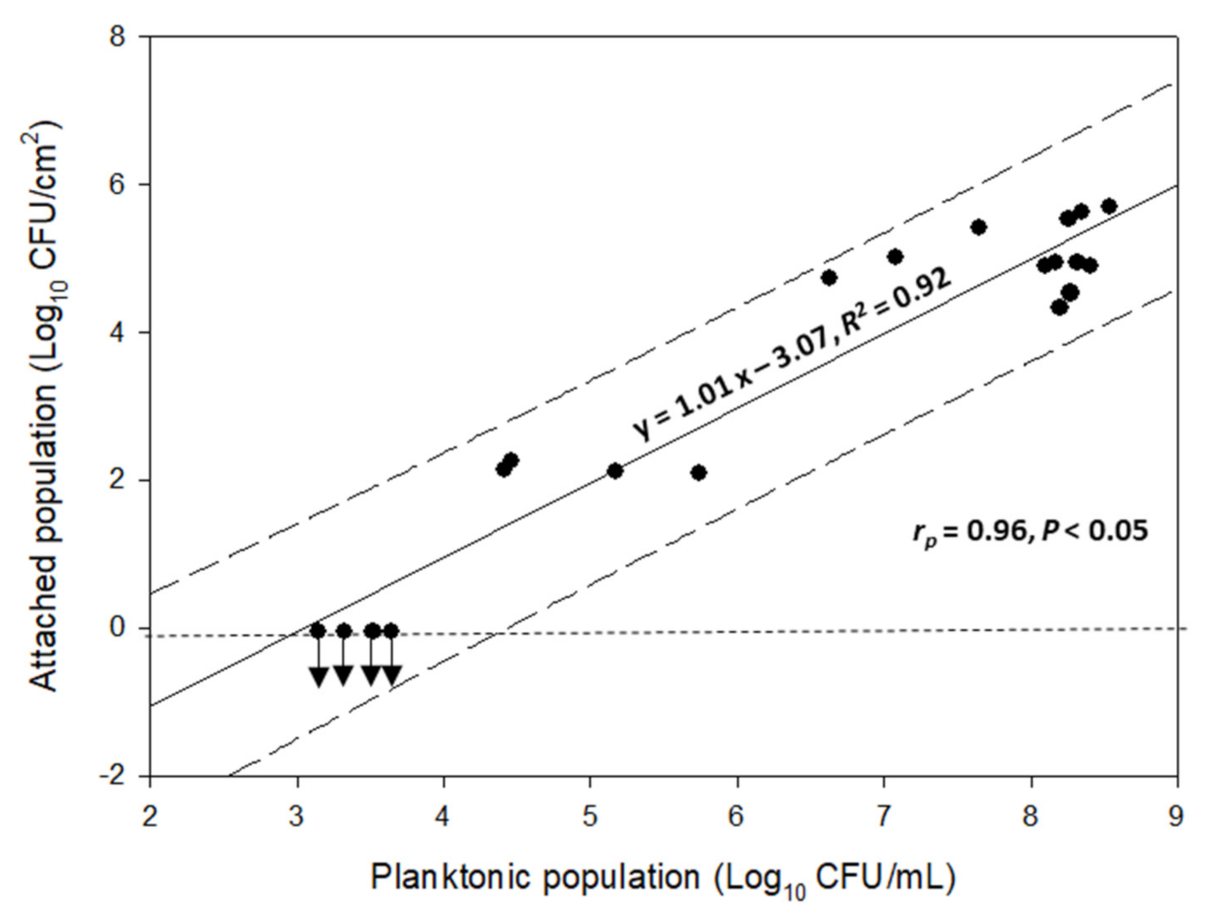
3.1. Combined Influence of Time and Temperature on Sessile and Planktonic Cell Numbers
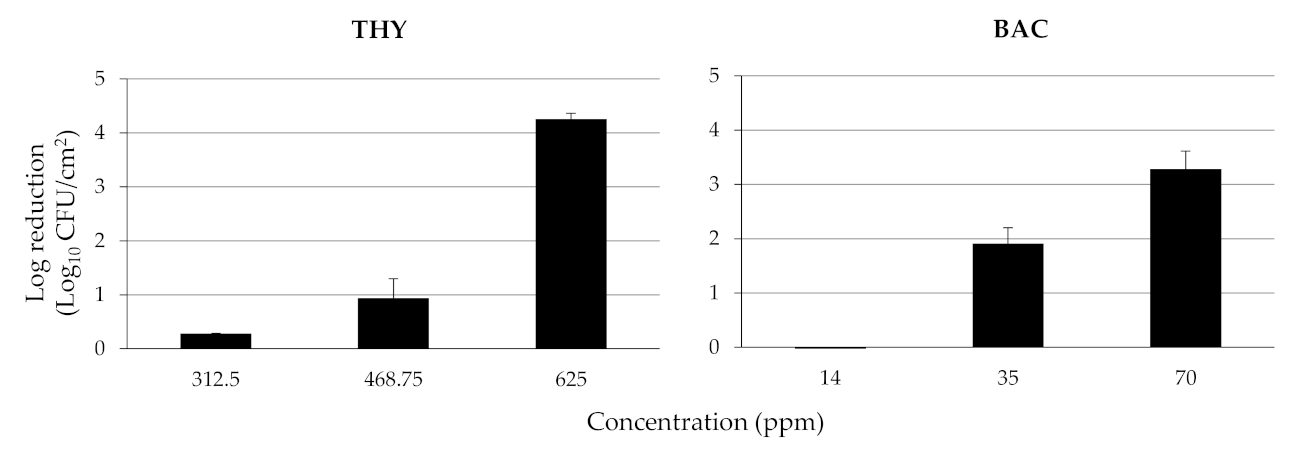
3.2. Calculation of MICs and MBCs of THY and BAC against Planktonic Cells and Disinfection of the Mixed Sessile Community
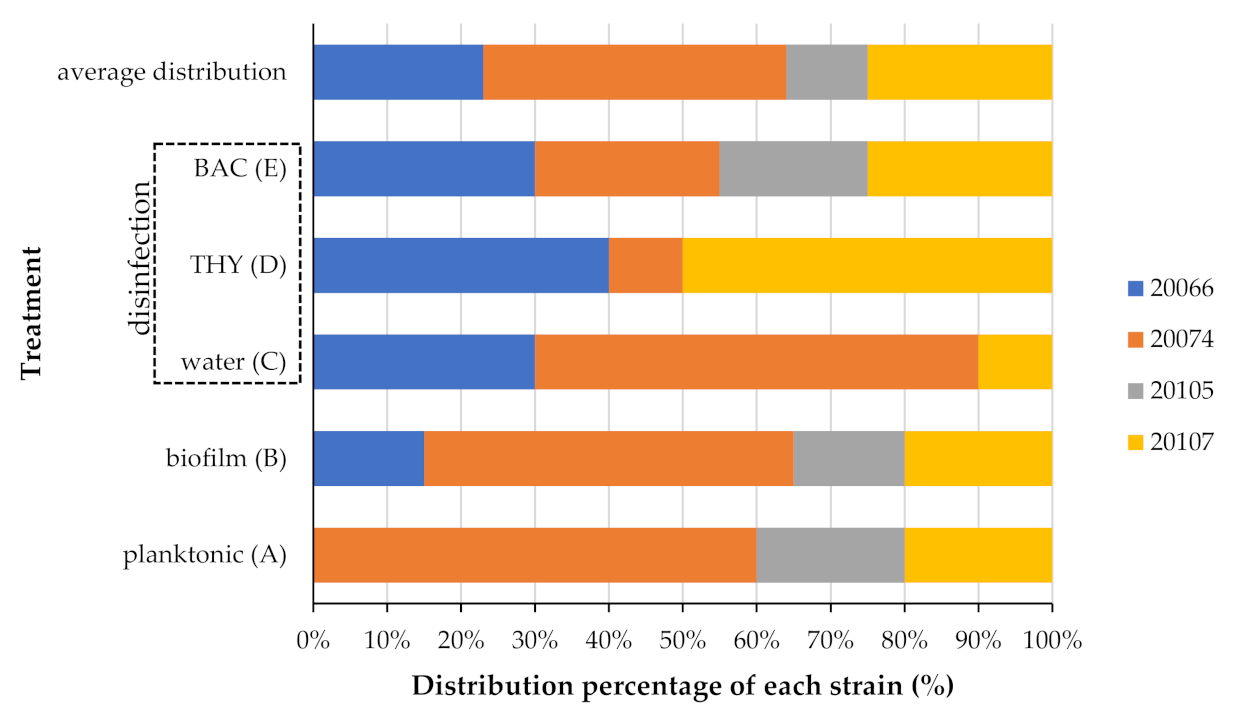
3.3. Strain Variability on Planktonic Growth, Attachment, and Disinfection Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jordan, K.; McAuliffe, O. Listeria monocytogenes in foods. Adv. Food Nutr. Res. 2018, 86, 181–213. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2018 zoonoses report. EFSA J. 2019, 17, 276. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; Moorhouse, E.; Monaghan, J.; Taylor, C.; Singleton, I. Sources and survival of Listeria monocytogenes on fresh, leafy produce. J. Appl. Microbiol. 2018, 125, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Matereke, L.T.; Okoh, A.I. Listeria monocytogenes virulence, antimicrobial resistance and environmental persistence: A review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Vázquez-Sánchez, D.; López Cabo, M. Current knowledge on Listeria monocytogenes biofilms in food-related environments: Incidence, resistance to biocides, ecology and biocontrol. Foods 2018, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, G.; Piccolomini, R.; Paludi, D.; D’Orio, V.; Vergara, A.; Conter, M.; Ianieri, A. Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact surfaces: Relationship with motility and cell surface hydrophobicity. J. Appl. Microbiol. 2008, 104, 1552–1561. [Google Scholar] [CrossRef]
- Folsom, J.P.; Siragusa, G.R.; Frank, J.F. Formation of biofilm at different nutrient levels by various genotypes of Listeria monocytogenes. J. Food Prot. 2006, 69, 826–834. [Google Scholar] [CrossRef]
- Kadam, S.R.; Den Besten, H.M.; Van der Veen, S.; Zwietering, M.H.; Moezelaar, R.; Abee, T. Diversity assessment of Listeria monocytogenes biofilm formation: Impact of growth condition, serotype and strain origin. Int. J. Food Microbiol. 2013, 165, 259–264. [Google Scholar] [CrossRef]
- Nilsson, R.E.; Ross, T.; Bowman, J.P. Variability in biofilm production by Listeria monocytogenes correlated to strain origin and growth conditions. Int. J. Food Microbiol. 2011, 150, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.; Cruz, C.D.; Palmer, J.; Fletcher, G.C.; Flint, S. Biofilm formation of the L. monocytogenes strain 15G01 is influenced by changes in environmental conditions. J. Microbiol. Methods 2015, 119, 189–195. [Google Scholar] [CrossRef]
- Tresse, O.; Shannon, K.; Pinon, A.; Malle, P.; Vialette, M.; Midelet-Bourdin, G. Variable adhesion of Listeria monocytogenes isolates from food-processing facilities and clinical cases to inert surfaces. J. Food Prot. 2007, 70, 1569–1578. [Google Scholar] [CrossRef]
- Piercey, M.J.; Hingston, P.A.; Truelstrup Hansen, L. Genes involved in Listeria monocytogenes biofilm formation at a simulated food processing plant temperature of 15 °C. Int. J. Food Microbiol. 2016, 223, 63–74. [Google Scholar] [CrossRef]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef]
- Skowron, K.; Wałecka-Zacharska, E.; Grudlewska, K.; Gajewski, P.; Wiktorczyk, N.; Wietlicka-Piszcz, M.; Dudek, A.; Skowron, K.J.; Gospodarek-Komkowska, E. Disinfectant susceptibility of biofilm formed by Listeria monocytogenes under selected environmental conditions. Microorganisms 2019, 7, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.A.; Chandry, P.S.; Kaur, M.; Kocharunchitt, C.; Bowman, J.P.; Fox, E.M. Novel biocontrol methods for Listeria monocytogenes biofilms in food production facilities. Front. Microbiol. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diana, M.; Felipe-Sotelo, M.; Bond, T. Disinfection byproducts potentially responsible for the association between chlorinated drinking water and bladder cancer: A review. Water Res. 2019, 162, 492–504. [Google Scholar] [CrossRef]
- Ferk, F.; Misík, M.; Hoelzl, C.; Uhl, M.; Fuerhacker, M.; Grillitsch, B.; Parzefall, W.; Nersesyan, A.; Micieta, K.; Grummt, T.; et al. Benzalkonium chloride (BAC) and dimethyldioctadecyl-ammonium bromide (DDAB), two common quaternary ammonium compounds, cause genotoxic effects in mammalian and plant cells at environmentally relevant concentrations. Mutagenesis 2007, 22, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Abreu, A.C.; Dias, C.; Saavedra, M.J.; Borges, F.; Simões, M. New perspectives on the use of phytochemicals as an emergent strategy to control bacterial infections including biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and thyme essential oil–new insights into selected therapeutic applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef]
- Cusimano, M.G.; Di Stefano, V.; La Giglia, M.; Di Marco Lo Presti, V.; Schillaci, D.; Pomilio, F.; Vitale, M. Control of growth and persistence of Listeria monocytogenes and β–lactam-resistant Escherichia coli by thymol in food processing settings. Molecules 2020, 25, 383. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, S.; Morán, A.; Martínez-Blanco, H.; Ferrero, M.A.; Rodríguez-Aparicio, L.B. The usefulness of non-toxic plant metabolites in the control of bacterial proliferation. Probiotics Antimicrob. Proteins. 2017, 9, 323–333. [Google Scholar] [CrossRef]
- Kerekes, E.B.; Vidács, A.; Takó, M.; Petkovits, T.; Vágvölgyi, C.; Horváth, G.; Balázs, V.L.; Krisch, J. Anti-biofilm effect of selected essential oils and main components on mono–and polymicrobic bacterial cultures. Microorganisms 2019, 7, 345. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Niu, H.; Zhang, W.; Mu, H.; Sun, C.; Duan, J. Synergy among thymol, eugenol, berberine, cinnamaldehyde and streptomycin against planktonic and biofilm-associated food-borne pathogens. Lett. Appl. Microbiol. 2015, 60, 421–430. [Google Scholar] [CrossRef]
- Upadhyay, A.; Upadhyaya, I.; Kollanoor-Johny, A.; Venkitanarayanan, K. Antibiofilm effect of plant derived antimicrobials on Listeria monocytogenes. Food Microbiol. 2013, 36, 79–89. [Google Scholar] [CrossRef]
- Karampoula, F.; Giaouris, E.; Deschamps, J.; Doulgeraki, A.I.; Nychas, G.J.; Dubois-Brissonnet, F. Hydrosol of Thymbra capitata is a highly efficient biocide against Salmonella enterica serovar Typhimurium biofilms. Appl. Environ. Microbiol. 2016, 82, 5309–5319. [Google Scholar] [CrossRef] [Green Version]
- Kostoglou, D.; Protopappas, I.; Giaouris, E. Common plant-derived terpenoids present increased anti-biofilm potential against Staphylococcus bacteria compared to a quaternary ammonium biocide. Foods 2020, 9, 697. [Google Scholar] [CrossRef]
- Lebert, I.; Leroy, S.; Talon, R. Effect of industrial and natural biocides on spoilage, pathogenic and technological strains grown in biofilm. Food Microbiol. 2007, 24, 281–287. [Google Scholar] [CrossRef]
- Merchel Piovesan Pereira, B.; Tagkopoulos, I. Benzalkonium chlorides: Uses, regulatory status, and microbial resistance. Appl. Environ. Microbiol. 2019, 85, 00377-19. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.I.; Macagnan, K.L.; Piecha, C.R.; Torres, M.M.; Perez, I.A.; Kesserlingh, S.M.; Rodrigues, R.D.S.; De Oliveira, P.D.; Moreira, A.D.S. Optimization of Ralstonia solanacearum cell growth using a central composite rotational design for the P(3HB) production: Effect of agitation and aeration. PLoS ONE 2019, 14, e0211211. [Google Scholar] [CrossRef] [Green Version]
- Hadjilouka, A.; Andritsos, N.D.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Listeria monocytogenes serotype prevalence and biodiversity in diverse food products. J. Food Prot. 2014, 77, 2115–2120. [Google Scholar] [CrossRef]
- Iliadis, I.; Daskalopoulou, A.; Simões, M.; Giaouris, E. Integrated combined effects of temperature, pH and sodium chloride concentration on biofilm formation by Salmonella enterica ser. Enteritidis and Typhimurium under low nutrient food-related conditions. Food Res. Int. 2018, 107, 10–18. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Paramithiotis, S.; Nychas, G.J. Characterization of the Enterobacteriaceae community that developed during storage of minced beef under aerobic or modified atmosphere packaging conditions. Int. J. Food Microbiol. 2011, 145, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Schmid, F.-X. Biological macromolecules: UV-visible spectrophotometry. In Encyclopedia of Life Sciences; Willey-Blackwell: Hoboken, NJ, USA, 2001. [Google Scholar]
- Ross, T. Indices for performance evaluation of predictive models in food microbiology. J. Appl. Bacteriol. 1996, 81, 501–508. [Google Scholar] [CrossRef]
- Jang, Y.S.; Mosolygó, T. Inhibition of bacterial biofilm formation by phytotherapeutics with focus on overcoming antimicrobial resistance. Curr. Pharm. Des. 2020, 26, 2807–2816. [Google Scholar] [CrossRef] [PubMed]
- Sakarikou, C.; Kostoglou, D.; Simões, M.; Giaouris, E. Exploitation of plant extracts and phytochemicals against resistant Salmonella spp. in biofilms. Food Res. Int. 2020, 128, 108806. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.R.; Burall, L.S. Serotype to genotype: The changing landscape of listeriosis outbreak investigations. Food Microbiol. 2018, 75, 18–27. [Google Scholar] [CrossRef]
- Maćkiw, E.; Stasiak, M.; Kowalska, J.; Kucharek, K.; Korsak, D.; Postupolski, J. Occurrence and characteristics of Listeria monocytogenes in ready-to-eat meat products in Poland. J. Food Prot. 2020, 83, 1002–1009. [Google Scholar] [CrossRef]
- Zhang, Y.; Yeh, E.; Hall, G.; Cripe, J.; Bhagwat, A.A.; Meng, J. Characterization of Listeria monocytogenes isolated from retail foods. Int. J. Food Microbiol. 2007, 113, 47–53. [Google Scholar] [CrossRef]
- Candioti, L.V.; De Zan, M.M.; Cámara, M.S.; Goicoechea, H.C. Experimental design and multiple response optimization. Using the desirability function in analytical methods development. Talanta 2014, 124, 123–138. [Google Scholar] [CrossRef]
- Bezerra, M.A.; Santelli, R.E.; Oliveira, E.P.; Villar, L.S.; Escaleira, L.A. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta 2008, 76, 965–977. [Google Scholar] [CrossRef]
- Giaouris, E.; Chorianopoulos, N.; Nychas, G.J. Impact of acid adaptation on attachment of Listeria monocytogenes to stainless steel during long-term incubation under low or moderate temperature conditions and on subsequent recalcitrance of attached cells to lethal acid treatments. Int. J. Food Microbiol. 2014, 171, 1–7. [Google Scholar] [CrossRef]
- Ding, T.; Dong, Q.-L.; Rahman, S.M.E.; Oh, D.-H. Response surface modeling of Listeria monocytogenes inactivation on lettuce treated with electrolyzed oxidizing water. J. Food Process. Eng. 2011, 34, 1729–1745. [Google Scholar] [CrossRef]
- Rubio, B.; Possas, A.; Rincón, F.; García-Gímeno, R.M.; Martínez, B. Model for Listeria monocytogenes inactivation by high hydrostatic pressure processing in Spanish chorizo sausage. Food Microbiol. 2018, 69, 18–24. [Google Scholar] [CrossRef]
- Ganjali Dashti, M.; Abdeshahian, P.; Sudesh, K.; Phua, K.K. Optimization of Salmonella Typhi biofilm assay on polypropylene microtiter plates using response surface methodology. Biofouling 2016, 32, 477–487. [Google Scholar] [CrossRef]
- Kim, H.W.; Rhee, M.S. Response surface modeling of reductions in uropathogenic Escherichia coli biofilms on silicone by cranberry extract, caprylic acid, and thymol. Biofouling 2018, 34, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Rosado de Castro, M.; Da Silva Fernandes, M.; Kabuki, D.Y.; Kuaye, A.Y. Biofilm formation on stainless steel as a function of time and temperature and control through sanitizers. Int. Dairy J. 2017, 68, 9–16. [Google Scholar] [CrossRef]
- Allende, A.; Martínez, B.; Selma, V.; Gil, M.; Suárez, J.E.; Rodríguez, A. Growth and bacteriocin production by lactic acid bacteria in vegetable broth and their effectiveness at reducing Listeria monocytogenes in vitro and in fresh-cut lettuce. Food Microbiol. 2007, 24, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Crozier, L.; Hedley, P.E.; Morris, J.; Wagstaff, C.; Andrews, S.C.; Toth, I.; Jackson, R.W.; Holden, N.J. Whole-transcriptome analysis of verocytotoxigenic Escherichia coli O157:H7 (Sakai) suggests plant-species-specific metabolic responses on exposure to spinach and lettuce extracts. Front. Microbiol. 2016, 7, 1088. [Google Scholar] [CrossRef] [Green Version]
- De Abrew Abeysundara, P.; Dhowlaghar, N.; Nannapaneni, R.; Schilling, M.W.; Chang, S.; Mahmoud, B.; Sharma, C.S.; Ma, D.-P. Growth and biofilm formation by Listeria monocytogenes in cantaloupe flesh and peel extracts on four food-contact surfaces at 22 °C and 10 °C. Food Control 2017, 80, 131–142. [Google Scholar] [CrossRef]
- Hood, S.K.; Zottola, E.A. Adherence to stainless steel by foodborne microorganisms during growth in model food systems. Int. J. Food Microbiol. 1997, 37, 145–153. [Google Scholar] [CrossRef]
- Papaioannou, E.; Giaouris, E.D.; Berillis, P.; Boziaris, I.S. Dynamics of biofilm formation by Listeria monocytogenes on stainless steel under mono-species and mixed-culture simulated fish processing conditions and chemical disinfection challenges. Int. J. Food Microbiol. 2018, 267, 9–19. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hébraud, M.; Møretrø, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.J.; Kačániová, M.; Czaczyk, K.; et al. Intra- and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 841. [Google Scholar] [CrossRef]
- Khan, F.; Javaid, A.; Kim, Y.M. Functional diversity of quorum sensing receptors in pathogenic bacteria: Interspecies, intraspecies and interkingdom level. Curr. Drug Targets 2019, 20, 655–667. [Google Scholar] [CrossRef]
- Bezek, K.; Nipič, D.; Torkar, K.G.; Oder, M.; Dražić, G.; Abram, A.; Žibert, J.; Raspor, P.; Bohinc, K. Biofouling of stainless steel surfaces by four common pathogens: The effects of glucose concentration, temperature and surface roughness. Biofouling 2019, 35, 273–283. [Google Scholar] [CrossRef]
- Nowak, J.; Cruz, C.D.; Tempelaars, M.; Abee, T.; Van Vliet, A.H.M.; Fletcher, G.C.; Hedderley, D.; Palmer, J.; Flint, S. Persistent Listeria monocytogenes strains isolated from mussel production facilities form more biofilm but are not linked to specific genetic markers. Int. J. Food Microbiol. 2017, 256, 45–53. [Google Scholar] [CrossRef]
- Lee, H.Y.; Chai, L.C.; Pui, C.F.; Mustafa, S.; Cheah, Y.K.; Nishibuchi, M.; Radu, S. Formation of biofilm by Listeria monocytogenes ATCC 19112 at different incubation temperatures and concentrations of sodium chloride. Braz. J. Microbiol. 2013, 44, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Mendez, E.; Walker, D.K.; Vipham, J.; Trinetta, V. The use of a CDC biofilm reactor to grow multi-strain Listeria monocytogenes biofilm. Food Microbiol. 2020, 92, 103592. [Google Scholar] [CrossRef]
- Ochiai, Y.; Yamada, F.; Mochizuki, M.; Takano, T.; Hondo, R.; Ueda, F. Biofilm formation under different temperature conditions by a single genotype of persistent Listeria monocytogenes strains. J. Food Prot. 2014, 77, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Kim, H.; Beuchat, L.R.; Ryu, J.H. Synergistic activities of gaseous oregano and thyme thymol essential oils against Listeria monocytogenes on surfaces of a laboratory medium and radish sprouts. Food Microbiol. 2020, 86, 103357. [Google Scholar] [CrossRef]
- Barroso, I.; Maia, V.; Cabrita, P.; Martínez-Suárez, J.V.; Brito, L. The benzalkonium chloride resistant or sensitive phenotype of Listeria monocytogenes planktonic cells did not dictate the susceptibility of its biofilm counterparts. Food Res. Int. 2019, 123, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Heir, E.; Lindstedt, B.A.; Røtterud, O.J.; Vardund, T.; Kapperud, G.; Nesbakken, T. Molecular epidemiology and disinfectant susceptibility of Listeria monocytogenes from meat processing plants and human infections. Int. J. Food Microbiol. 2004, 96, 85–96. [Google Scholar] [CrossRef]
- Minarovičová, J.; Véghová, A.; Mikulášová, M.; Chovanová, R.; Šoltýs, K.; Drahovská, H.; Kaclíková, E. Benzalkonium chloride tolerance of Listeria monocytogenes strains isolated from a meat processing facility is related to presence of plasmid-borne bcrABC cassette. Antonie Van Leeuwenhoek 2018, 111, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Brauge, T.; Faille, C.; Leleu, G.; Denis, C.; Hanin, A.; Midelet, G. Treatment with disinfectants may induce an increase in viable but non culturable populations of Listeria monocytogenes in biofilms formed in smoked salmon processing environments. Food Microbiol. 2020, 92, 103548. [Google Scholar] [CrossRef] [PubMed]
- Noll, M.; Trunzer, K.; Vondran, A.; Vincze, S.; Dieckmann, R.; Al Dahouk, S.; Gold, C. Benzalkonium chloride induces a VBNC state in Listeria monocytogenes. Microorganisms 2020, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- De Medeiros Barbosa, I.; Da Cruz Almeida, É.T.; Gomes, A.C.A.; De Souza, E.L. Evidence on the induction of viable but non-culturable state in Listeria monocytogenes by Origanum vulgare L. and Rosmarinus officinalis L. essential oils in a meat-based broth. Innov. Food Sci. Emerg Technol. 2020, 62, 102351. [Google Scholar] [CrossRef]
- Baldy-Chudzik, K. Rep-PCR–a variant to RAPD or an independent technique of bacteria genotyping? A comparison of the typing properties of rep-PCR with other recognised methods of genotyping of microorganisms. Acta Microbiol. Pol. 2001, 50, 189–204. [Google Scholar]
- Lianou, A.; Nychas, G.E.; Koutsoumanis, K.P. Strain variability in biofilm formation: A food safety and quality perspective. Food Res. Int. 2020, 137, 109424. [Google Scholar] [CrossRef]
- Giaouris, E.; Chorianopoulos, N.; Doulgeraki, A.; Nychas, G.J. Co-culture with Listeria monocytogenes within a dual-species biofilm community strongly increases resistance of Pseudomonas putida to benzalkonium chloride. PLoS ONE 2013, 8, 77276. [Google Scholar] [CrossRef] [Green Version]
- Heir, E.; Møretrø, T.; Simensen, A.; Langsrud, S. Listeria monocytogenes strains show large variations in competitive growth in mixed culture biofilms and suspensions with bacteria from food processing environments. Int. J. Food Microbiol. 2018, 275, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.J.; Giaouris, E. Differential biofilm formation and chemical disinfection resistance of sessile cells of Listeria monocytogenes strains under monospecies and dual-species (with Salmonella enterica) conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Independent Factors a | Response b | ||
|---|---|---|---|---|
| Incubation Time (h) (Χ1) | Incubation Temperature (°C) (Χ2) | Attached Population (Log10 CFU/cm2) | ||
| Measured | Predicted | |||
| 1 | 14.1 (−1.414) | 20 (0) | 2.12 ± 0.22 | 2.61 ± 0.25 |
| 2 | 72 (1) | 8 (−1) | 2.26 ± 0.32 | 2.50 ± 0.25 |
| 3 | 48 (0) | 37 (1.414) | 5.01 ± 0.20 | 5.41 ± 0.25 |
| 4 | 48 (0) | 37 (1.414) | 4.74 ± 0.50 | 5.41 ± 0.25 |
| 5 | 72 (1) | 8 (−1) | 2.14 ± 0.34 | 2.50 ± 0.25 |
| 6 | 24 (−1) | 8 (−1) | −0.05c ± 0.00 | −0.20 ± 0.25 |
| 7 | 48 (0) | 3 (−1.414) | −0.05c ± 0.00 | −0.23 ± 0.25 |
| 8 | 24 (−1) | 32 (1) | 5.69 ± 0.07 | 4.96 ± 0.25 |
| 9 | 24 (−1) | 32 (1) | 5.54 ± 0.16 | 4.96 ± 0.25 |
| 10 | 48 (0) | 3 (−1.414) | −0.05c ± 0.00 | −0.23 ± 0.25 |
| 11 | 48 (0) | 20 (0) | 4.94 ± 0.09 | 4.68 ± 0.22 |
| 12 | 81.9 (1.414) | 20 (0) | 4.90 ± 0.12 | 4.78 ± 0.25 |
| 13 | 14.1 (−1.414) | 20 (0) | 2.10 ± 0.30 | 2.61 ± 0.25 |
| 14 | 72 (1) | 32 (1) | 5.62 ± 0.02 | 5.31 ± 0.25 |
| 15 | 48 (0) | 20 (0) | 4.90 ± 0.38 | 4.68 ± 0.22 |
| 16 | 24 (−1) | 8 (−1) | −0.05c ± 0.00 | −0.20 ± 0.25 |
| 17 | 48 (0) | 20 (0) | 4.33 ± 1.04 | 4.68 ± 0.22 |
| 18 | 81.9 (1.414) | 20 (0) | 4.94 ± 0.20 | 4.78 ± 0.25 |
| 19 | 48 (0) | 20 (0) | 4.53 ± 0.75 | 4.68 ± 0.22 |
| 20 | 72 (1) | 32 (1) | 5.41 ± 0.04 | 5.31 ± 0.25 |
| Response | Polynomial Equation | R2 | R2adj | p |
|---|---|---|---|---|
| Log10 CFU/cm2 | 4.68 + 0.77 × t + 1.99 × T − 0.59(t × T) − 0.49 × t2 − 1.04 × T2 | 0.97 | 0.96 | <0.0001 |
| Parameter | Estimated Value a | 95% Confidence Limits | p-Value | t Ratio | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
| Intercept | 4.68 ± 0.22 | 4.20 | 5.15 | <0.0001 | 21.32 |
| t | 0.77 ± 0.11 | 0.53 | 1.00 | <0.0001 | 6.98 |
| T | 1.99 ± 0.11 | 1.76 | 2.23 | <0.0001 | 18.18 |
| t * T | −0.59 ± 0.16 | −0.92 | −0.25 | 0.002 | −3.79 |
| t2 | −0.49 ± 0.15 | −0.80 | −0.18 | 0.004 | −3.38 |
| T2 | −1.04 ± 0.15 | −1.35 | −0.73 | <0.0001 | −7.18 |
| Experiment | Independent Factors a | Response b | ||
|---|---|---|---|---|
| Incubation Time (h) (Χ1) | Incubation Temperature (°C) (Χ2) | Attached Population (Log10 CFU/cm2) | ||
| Measured | Predicted | |||
| 1 | 36 (−0.5) | 26 (0.5) | 5.39 ± 0.14 | 5.05 ± 0.40 |
| 2 | 60 (0.5) | 26 (0.5) | 5.40 ± 0.09 | 5.52 ± 0.40 |
| 3 | 66 (0.75) | 11 (−0.75) | 2.96 ± 0.37 | 3.22 ± 0.40 |
| 4 | 66 (0.75) | 29 (0.75) | 5.58 ± 0.05 | 5.55 ± 0.40 |
| 5 (max) | 54 (0.25) | 30.6 (0.89) | 5.46 ± 0.31 | 5.65 ± 0.40 |
| Factors | ||||
| Bias | 1.01 | |||
| Accuracy | 1.04 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostoglou, D.; Tsaklidou, P.; Iliadis, I.; Garoufallidou, N.; Skarmoutsou, G.; Koulouris, I.; Giaouris, E. Advanced Killing Potential of Thymol against a Time and Temperature Optimized Attached Listeria monocytogenes Population in Lettuce Broth. Biomolecules 2021, 11, 397. https://doi.org/10.3390/biom11030397
Kostoglou D, Tsaklidou P, Iliadis I, Garoufallidou N, Skarmoutsou G, Koulouris I, Giaouris E. Advanced Killing Potential of Thymol against a Time and Temperature Optimized Attached Listeria monocytogenes Population in Lettuce Broth. Biomolecules. 2021; 11(3):397. https://doi.org/10.3390/biom11030397
Chicago/Turabian StyleKostoglou, Dimitra, Parthena Tsaklidou, Ioannis Iliadis, Nikoletta Garoufallidou, Georgia Skarmoutsou, Ioannis Koulouris, and Efstathios Giaouris. 2021. "Advanced Killing Potential of Thymol against a Time and Temperature Optimized Attached Listeria monocytogenes Population in Lettuce Broth" Biomolecules 11, no. 3: 397. https://doi.org/10.3390/biom11030397
APA StyleKostoglou, D., Tsaklidou, P., Iliadis, I., Garoufallidou, N., Skarmoutsou, G., Koulouris, I., & Giaouris, E. (2021). Advanced Killing Potential of Thymol against a Time and Temperature Optimized Attached Listeria monocytogenes Population in Lettuce Broth. Biomolecules, 11(3), 397. https://doi.org/10.3390/biom11030397