
RNA-seq Characterization of Melanoma Phenotype Switch in 3D Collagen after p38 MAPK Inhibitor Treatment

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells Culture, Treatments and Morphological Analysis
2.2. RNA Extraction and Sequencing
2.3. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
2.4. Statistical Analysis
3. Results
3.1. SB202190 and BIRB796 Induce a Phenotype Switch in A375M2 Cells Cultured in 3D Collagen
3.2. Transcriptomic Profiling of the Phenotype Switch-Associated Changes with RNA-seq
3.3. Validation and Reproducibility of the RNA-seq Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rambow, F.; Marine, J.-C.; Goding, C.R. Melanoma plasticity and phenotypic diversity: Therapeutic barriers and opportunities. Genes Dev. 2019, 33, 1295–1318. [Google Scholar] [CrossRef] [Green Version]
- Goding, C.R.; Arnheiter, H. Mitf—the First 25 Years. Genes Dev. 2019, 33, 983–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, M.L.; Czyz, M. MITF in melanoma: Mechanisms behind its expression and activity. Cell. Mol. Life Sci. 2015, 72, 1249–1260. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Hong, A.-R.; Kim, Y.-H.; Yoo, H.; Kang, S.-W.; Chang, S.E.; Song, Y. JNK suppresses melanogenesis by interfering with CREB-regulated transcription coactivator 3-dependent MITF expression. Theranostics 2020, 10, 4017–4029. [Google Scholar] [CrossRef]
- Falletta, P.; Sanchez-Del-Campo, L.; Chauhan, J.; Effern, M.; Kenyon, A.; Kershaw, C.J.; Siddaway, R.; Lisle, R.J.; Freter, R.; Daniels, M.J.; et al. Translation reprogramming is an evolutionarily conserved driver of phenotypic plasticity and therapeutic resistance in melanoma. Genes Dev. 2017, 31, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Buscà, R.; Bertolotto, C.; Abbe, P.; Englaro, W.; Ishizaki, T.; Narumiya, S.; Boquet, P.; Ortonne, J.P.; Ballotti, R. Inhibition of rho is required for CAMP-induced melanoma cell differentiation. Mol. Biol. Cell 1998, 9, 1367–1378. [Google Scholar] [CrossRef] [Green Version]
- Hoek, K.S.; Schlegel, N.C.; Brafford, P.; Sucker, A.; Ugurel, S.; Kumar, R.; Weber, B.L.; Nathanson, K.L.; Phillips, D.J.; Herlyn, M.; et al. Metastatic potential of melanomas defined by specific gene expression profiles with no BRAF signature. Pigment. Cell Res. 2006, 19, 290–302. [Google Scholar] [CrossRef]
- Vivas-García, Y.; Falletta, P.; Liebing, J.; Louphrasitthiphol, P.; Feng, Y.; Chauhan, J.; Scott, D.A.; Glodde, N.; Chocarro-Calvo, A.; Bonham, S.; et al. Lineage-restricted regulation of SCD and fatty acid saturation by MITF controls melanoma phenotypic plasticity. Mol. Cell 2020, 77, 120–137.e9. [Google Scholar] [CrossRef]
- Misek, S.A.; Appleton, K.M.; Dexheimer, T.S.; Lisabeth, E.M.; Lo, R.S.; Larsen, S.D.; Gallo, K.A.; Neubig, R.R. Rho-mediated signaling promotes BRAF inhibitor resistance in de-differentiated melanoma cells. Oncogene 2020, 39, 1466–1483. [Google Scholar] [CrossRef] [PubMed]
- Konieczkowski, D.J.; Johannessen, C.M.; Abudayyeh, O.; Kim, J.W.; Cooper, Z.A.; Piris, A.; Frederick, D.T.; Barzily-Rokni, M.; Straussman, R.; Haq, R.; et al. A melanoma cell state distinction influences sensitivity to MAPK pathway inhibitors. Cancer Discov. 2014, 4, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verfaillie, A.; Imrichova, H.; Atak, Z.K.; Dewaele, M.; Rambow, F.; Hulselmans, G.; Christiaens, V.; Svetlichnyy, D.; Luciani, F.; Van Der Mooter, L.L.; et al. Decoding the regulatory landscape of melanoma reveals TEADS as regulators of the invasive cell state. Nat. Commun. 2015, 6, 6683. [Google Scholar] [CrossRef] [Green Version]
- Kholmanskikh, O.; Van Baren, N.; Brasseur, F.; Ottaviani, S.; Vanacker, J.; Arts, N.; Van Der Bruggen, P.; Coulie, P.; De Plaen, E. Interleukins 1α and 1β secreted by some melanoma cell lines strongly reduce expression of MITF-M and melanocyte differentiation antigens. Int. J. Cancer 2010, 127, 1625–1636. [Google Scholar] [CrossRef]
- Miskolczi, Z.; Smith, M.P.; Rowling, E.J.; Ferguson, J.; Barriuso, J.; Wellbrock, C. Collagen abundance controls melanoma phenotypes through lineage-specific microenvironment sensing. Oncogene 2018, 37, 3166–3182. [Google Scholar] [CrossRef] [Green Version]
- Strub, T.; Kobi, D.; Koludrovic, D.; Davidson, I. Research on Melanoma—A Glimpse into Current Directions and Future Trends; Murph, M., Ed.; IntechOpen: London, UK, 2011. [Google Scholar]
- Thurber, A.; Douglas, G.; Sturm, E.; Zabierowski, S.; Smit, D.; Ramakrishnan, S.; Hacker, E.; Leonard, J.; Herlyn, M.; Sturm, R. Inverse expression states of the BRN2 and MITF transcription factors in melanoma spheres and tumour xenografts regulate the NOTCH pathway. Oncogene 2011, 30, 3036–3048. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, M.P.; Marchbank, K.; Webster, M.R.; Valiga, A.A.; Kaur, A.; Vultur, A.; Li, L.; Herlyn, M.; Villanueva, J.; Liu, Q.; et al. Hypoxia induces phenotypic plasticity and therapy resistance in melanoma via the tyrosine kinase receptors ROR1 and ROR2. Cancer Discov. 2013, 3, 1378–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landsberg, J.; Kohlmeyer, J.; Renn, M.; Bald, T.; Rogava, M.; Cron, M.; Fatho, M.; Lennerz, V.; Wölfel, T.; Hölzel, M.; et al. Melanomas resist T-cell therapy through inflammation-induced reversible dedifferentiation. Nature 2012, 490, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Kim, Y.J.; Robert, L.; Tsoi, J.; Comin-Anduix, B.; Berent-Maoz, B.; Cochran, A.J.; Economou, J.S.; Tumeh, P.C.; Puig-Saus, C.; et al. Immunotherapy resistance by inflammation-induced dedifferentiation. Cancer Discov. 2018, 8, 935–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.; Krijgsman, O.; Tsoi, J.; Robert, L.; Hugo, W.; Song, C.; Kong, X.; Possik, P.A.; Cornelissen-Steijger, P.D.M.; Foppen, M.H.G.; et al. Low MITF/AXL ratio predicts early resistance to multiple targeted drugs in melanoma. Nat. Commun. 2014, 5, 5712. [Google Scholar] [CrossRef]
- Kozar, I.; Margue, C.; Rothengatter, S.; Haan, C.; Kreis, S. Many ways to resistance: How melanoma cells evade targeted therapies. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 313–322. [Google Scholar] [CrossRef]
- Arozarena, I.; Wellbrock, C. Phenotype plasticity as enabler of melanoma progression and therapy resistance. Nat. Rev. Cancer 2019, 19, 377–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustakas, A.; Heldin, C.H. Non-smad TGF-β signals. J. Cell Sci. 2005, 118, 3573–3584. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, A.; Heldin, C.-H. Dynamic control of TGF-β signaling and its links to the cytoskeleton. FEBS Lett. 2008, 582, 2051–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-G.; Islam, R.; Cho, J.Y.; Jeong, H.; Cap, K.-C.; Park, Y.; Hossain, A.J.; Park, J.-B. Regulation of RhoA GTPase and various transcription factors in the RhoA pathway. J. Cell. Physiol. 2018, 233, 6381–6392. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Yao, E.; Zhang, K.; Jiang, X.; Croll, S.; Thompson-Peer, K.; Chuang, P.-T. YAP is essential for mechanical force production and epithelial cell proliferation during lung branching morphogenesis. eLife 2017, 6, e21130. [Google Scholar] [CrossRef]
- Edwards, D.N.; Ngwa, V.M.; Wang, S.; Shiuan, E.; Brantley-Sieders, D.M.; Kim, L.C.; Reynolds, A.B.; Chen, J. The receptor tyrosine kinase EphA2 promotes glutamine metabolism in tumors by activating the transcriptional coactivators YAP and TAZ. Sci. Signal. 2017, 10, eaan4667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Pang, K.M.; Iida, M.; Nelson, M.S.; Liu, J.; Nam, A.; Wang, J.; Mambetsariev, I.; Pillai, R.; Mohanty, A.; et al. Activation of EPHA2-ROBO1 heterodimer by SLIT2 attenuates non-canonical signaling and proliferation in squamous cell carcinomas. iScience 2020, 23, 101692. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Kanneganti, T.-D. Function and regulation of IL-1α in inflammatory diseases and cancer. Immunol. Rev. 2018, 281, 124–137. [Google Scholar] [CrossRef]
- Paňková, D.; Jobe, N.; Kratochvílová, M.; Buccione, R.; Brábek, J.; Rösel, D. NG2-mediated Rho activation promotes amoeboid invasiveness of cancer cells. Eur. J. Cell Biol. 2012, 91, 969–977. [Google Scholar] [CrossRef]
- Vaškovičová, K.; Szabadosová, E.; Čermák, V.; Gandalovičová, A.; Kasalová, L.; Rösel, D.; Brábek, J. PKCα promotes the mesenchymal to amoeboid transition and increases cancer cell invasiveness. BMC Cancer 2015, 15, 326. [Google Scholar] [CrossRef] [Green Version]
- Dovas, A.; Yoneda, A.; Couchman, J.R. PKC-α-dependent activation of RhoA by syndecan-4 during focal adhension formation. J. Cell Sci. 2006, 119, 2837–2846. [Google Scholar] [CrossRef] [Green Version]
- Riesenberg, S.; Groetchen, A.; Siddaway, R.; Bald, T.; Reinhardt, J.; Smorra, D.; Kohlmeyer, J.; Renn, M.; Phung, B.; Aymans, P.; et al. MITF and c-Jun antagonism interconnects melanoma dedifferentiation with pro-inflammatory cytokine responsiveness and myeloid cell recruitment. Nat. Commun. 2015, 6, 8755. [Google Scholar] [CrossRef]
- Arts, N.; Cané, S.; Hennequart, M.; Lamy, J.; Bommer, G.; Van Den Eynde, B.; De Plaen, E. microRNA-155, Induced by interleukin-1ß, represses the expression of microphthalmia-associated transcription factor (MITF-M) in melanoma cells. PLoS ONE 2015, 10, e0122517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soustek, M.S.; Balsa, E.; Barrow, J.J.; Jedrychowski, M.; Vogel, R.; Gygi, S.P.; Puigserver, P. Inhibition of the ER stress IRE1α inflammatory pathway protects against cell death in mitochondrial complex I mutant cells. Cell Death Dis. 2018, 9, 658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhari, N.; Talwar, P.; Parimisetty, A.; d’Hellencourt, C.L.; Ravanan, P. A molecular web: Endoplasmic reticulum stress, inflammation, and oxidative stress. Front. Cell. Neurosci. 2014, 8, 213. [Google Scholar] [CrossRef]
- Ju, R.J.; Stehbens, S.J.; Haass, N.K. The role of melanoma cell-stroma interaction in cell motility, invasion, and metastasis. Front. Med. 2018, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paňková, K.; Rösel, D.; Novotný, M.; Brábek, J. The molecular mechanisms of transition between mesenchymal and amoeboid invasiveness in tumor cells. Cell. Mol. Life Sci. 2009, 67, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosla, J.; Paňková, D.; Plachý, J.; Tolde, O.; Bicanová, K.; Dvořák, M.; Rösel, D.; Brábek, J. Metastasis of aggressive amoeboid sarcoma cells is dependent on Rho/ROCK/MLC signaling. Cell Commun. Signal. 2013, 11, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micuda, S.; Rösel, D.; Ryska, A.; Brábek, J. ROCK inhibitors as emerging therapeutic candidates for sarcomas. Curr. Cancer Drug Targets 2010, 10, 127–134. [Google Scholar] [CrossRef]
- Voller, J.; Zahajská, L.; Plíhalová, L.; Jeřábková, J.; Burget, D.; Pataki, A.C.; Kryštof, V.; Zatloukal, M.; Brábek, J.; Rösel, D.; et al. 6-substituted purines as ROCK inhibitors with anti-metastatic activity. Bioorg. Chem. 2019, 90, 103005. [Google Scholar] [CrossRef]
- Cantelli, G.; Orgaz, J.L.; Rodriguez-Hernandez, I.; Karagiannis, P.; Maiques, O.; Matias-Guiu, X.; Nestle, F.O.; Marti, R.M.; Karagiannis, S.N.; Sanz-Moreno, V. TGF-β-induced transcription sustains amoeboid melanoma migration and dissemination. Curr. Biol. 2015, 25, 2899–2914. [Google Scholar] [CrossRef] [Green Version]
- Georgouli, M.; Herraiz, C.; Crosas-Molist, E.; Fanshawe, B.; Maiques, O.; Perdrix, A.; Pandya, P.; Rodriguez-Hernandez, I.; Ilieva, K.M.; Cantelli, G.; et al. Regional activation of myosin II in cancer cells drives tumor progression via a secretory cross-talk with the immune microenvironment. Cell 2019, 176, 757–774.e23. [Google Scholar] [CrossRef] [PubMed]
- Merta, L.; Gandalovičová, A.; Čermák, V.; Dibus, M.; Gutschner, T.; Diederichs, S.; Rösel, D.; Brábek, J. Increased level of long non-coding RNA MALAT1 is a common feature of amoeboid invasion. Cancers 2020, 12, 1136. [Google Scholar] [CrossRef]
- Sanz-Moreno, V.; Gaggioli, C.; Yeo, M.; Albrengues, J.; Wallberg, F.; Viros, A.; Hooper, S.; Mitter, R.; Féral, C.C.; Cook, M.; et al. ROCK and JAK1 signaling cooperate to control actomyosin contractility in tumor cells and stroma. Cancer Cell 2011, 20, 229–245. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Moreno, V.; Gadea, G.; Ahn, J.; Paterson, H.; Marra, P.; Pinner, S.; Sahai, E.; Marshall, C.J. Rac activation and inactivation control plasticity of tumor cell movement. Cell 2008, 135, 510–523. [Google Scholar] [CrossRef] [Green Version]
- Karaman, M.W.; Herrgard, S.; Treiber, D.K.; Gallant, P.; Atteridge, C.E.; Campbell, B.T.; Chan, K.W.; Ciceri, P.; Davis, M.I.; Edeen, P.T.; et al. A quantitative analysis of kinase inhibitor selectivity. Nat. Biotechnol. 2008, 26, 127–132. [Google Scholar] [CrossRef]
- Bain, J.; Plater, L.; Elliott, M.; Shpiro, N.; Hastie, C.J.; Mclauchlan, H.; Klevernic, I.; Arthur, J.S.C.; Alessi, D.R.; Cohen, P. The selectivity of protein kinase inhibitors: A further update. Biochem. J. 2007, 408, 297–315. [Google Scholar] [CrossRef] [Green Version]
- Palušová, V.; Renzová, T.; Verlande, A.; Vaclová, T.; Medková, M.; Cetlová, L.; Sedláčková, M.; Hříbková, H.; Slaninová, I.; Krutá, M.; et al. Dual targeting of BRAF and mTOR signaling in melanoma cells with pyridinyl imidazole compounds. Cancers 2020, 12, 1516. [Google Scholar] [CrossRef]
- Yang, C.; Zhu, Z.; Tong, B.C.-K.; Iyaswamy, A.; Xie, W.-J.; Zhu, Y.; Sreenivasmurthy, S.G.; Senthilkumar, K.; Cheung, K.-H.; Song, J.-X.; et al. A stress response P38 MAP kinase inhibitor SB202190 promoted TFEB/TFE3-dependent autophagy and lysosomal biogenesis independent of P38. Redox Biol. 2020, 32, 101445. [Google Scholar] [CrossRef]
- Čermák, V.; Škarková, A.; Merta, L.; Rösel, D.; Brábek, J. Melanoma Phenotype Switch in 3D Collagen After P38 MAPK Inhibitor Treatment. Available online: https://doi.org/10.6084/m9.figshare.c.5215253 (accessed on 7 March 2021).
- Hemesath, T.J.; Steingrímsson, E.; McGill, G.; Hansen, M.J.; Vaught, J.; Hodgkinson, C.A.; Arnheiter, H.; Copeland, N.G.; Jenkins, N.A.; Fisher, D.E. Microphthalmia, a critical factor in melanocyte development, defines a discrete transcription factor family. Genes Dev. 1994, 8, 2770–2780. [Google Scholar] [CrossRef] [Green Version]
- Martina, J.A.; Puertollano, R. Rag GTPases mediate amino acid–dependent recruitment of TFEB and MITF to lysosomes. J. Cell Biol. 2013, 200, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Amae, S.; Fuse, N.; Yasumoto, K.I.; Sato, S.; Yajima, I.; Yamamoto, H.; Udono, T.; Durlu, Y.K.; Tamai, M.; Takahashi, K.; et al. Identification of a novel isoform of microphthalmia-associated transcription factor that is enriched in retinal pigment epithelium. Biochem. Biophys. Res. Commun. 1998, 247, 710–715. [Google Scholar] [CrossRef]
- Yasumoto, K.; Takeda, K.; Saito, H.; Watanabe, K.; Takahashi, K.; Shibahara, S. Microphthalmia-associated transcription factor interacts with LEF-1, a mediator of Wnt signaling. EMBO J. 2002, 21, 2703–2714. [Google Scholar] [CrossRef] [Green Version]
- Čermák, V.; Gandalovičová, A.; Merta, L.; Harant, K.; Rösel, D.; Brábek, J. High-throughput transcriptomic and proteomic profiling of mesenchymal-amoeboid transition in 3D collagen. Sci. Data 2020, 7, 1–11. [Google Scholar] [CrossRef]
- Clark, E.A.; Golub, T.R.; Lander, E.S.; Hynes, R.O. Genomic analysis of metastasis reveals an essential role for RhoC. Nature 2000, 406, 532–535. [Google Scholar] [CrossRef]
- Kozlowski, J.M.; Fidler, I.J.; Hanna, N.; Hart, I.R. A human melanoma line heterogeneous with respect to metastatic capacity in athymic nude mice. J. Natl. Cancer Inst. 1984, 72, 913–917. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Čermák, V.; Gandalovičová, A.; Merta, L.; Rösel, D.; Brábek, J. RNA-Seq Of Human Melanoma Cell Line A375m2 Treated with SB202190 or BIRB796 against DMSO-Treated Controls. Available online: https://identifiers.org/arrayexpress:E-MTAB-9273 (accessed on 28 November 2020).
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034. [Google Scholar] [CrossRef] [Green Version]
- Perkins, J.R.; Dawes, J.M.; McMahon, S.B.; Bennett, D.L.H.; Orengo, C.; Kohl, M. ReadqPCR and NormqPCR: R packages for the reading, quality checking and normalisation of RT-QPCR quantification cycle (Cq) data. BMC Genom. 2012, 13, 296. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Čermák, V.; Gandalovičová, A.; Merta, L.; Fučíková, J.; Špíšek, R.; Rösel, D.; Brábek, J. RNA-seq of macrophages of amoeboid or mesenchymal migratory phenotype due to specific structure of environment. Sci. Data 2018, 5, 180198. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Folberg, R.; Arbieva, Z.; Moses, J.; Hayee, A.; Sandal, T.; Kadkol, S.H.; Lin, A.Y.; Valyi-Nagy, K.; Setty, S.; Leach, L.; et al. Tumor cell plasticity in uveal melanoma: Microenvironment directed dampening of the invasive and metastatic genotype and phenotype accompanies the generation of vasculogenic mimicry patterns. Am. J. Pathol. 2006, 169, 1376–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molbiotools. Multiple List Comparator. Available online: http://www.molbiotools.com/listcompare.html (accessed on 4 March 2021).
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 21 November 2020).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widmer, D.S.; Cheng, P.F.; Eichhoff, O.M.; Belloni, B.C.; Zipser, M.C.; Schlegel, N.C.; Javelaud, D.; Mauviel, A.; Dummer, R.; Hoek, K.S. Systematic classification of melanoma cells by phenotype-specific gene expression mapping. Pigment. Cell Melanoma Res. 2012, 25, 343–353. [Google Scholar] [CrossRef]
- Han, H.; Cho, J.-W.; Lee, S.-Y.; Yun, A.; Kim, H.; Bae, D.; Yang, S.; Kim, C.Y.; Lee, M.; Kim, E.; et al. TRRUST v2: An expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018, 46, D380–D386. [Google Scholar] [CrossRef]
- Tirosh, I.; Izar, B.; Prakadan, S.M.; Wadsworth, M.H.; Treacy, D.; Trombetta, J.J.; Rotem, A.; Rodman, C.; Lian, C.; Murphy, G.; et al. Dissecting the multicellular ecosystem of metastatic melanoma by single-cell RNA-Seq. Science 2016, 352, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Jeffs, A.R.; Glover, A.C.; Slobbe, L.J.; Wang, L.; He, S.; Hazlett, J.A.; Awasthi, A.; Woolley, A.G.; Marshall, E.S.; Joseph, W.R.; et al. A gene expression signature of invasive potential in metastatic melanoma cells. PLoS ONE 2009, 4, e8461. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Hernandez, I.; Maiques, O.; Kohlhammer, L.; Cantelli, G.; Perdrix-Rosell, A.; Monger, J.; Fanshawe, B.; Bridgeman, V.L.; Karagiannis, S.N.; Penin, R.M.; et al. WNT11-FZD7-DAAM1 signalling supports tumour initiating abilities and melanoma amoeboid invasion. Nat. Commun. 2020, 11, 1–20. [Google Scholar] [CrossRef]
- Arozarena, I.; Bischof, H.; Gilby, D.; Belloni, B.; Dummer, R.; Wellbrock, C. In melanoma, beta-catenin is a suppressor of invasion. Oncogene 2011, 30, 4531–4543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parri, M.; Taddei, M.L.; Bianchini, F.; Calorini, L.; Chiarugi, P. EphA2 reexpression prompts invasion of melanoma cells shifting from mesenchymal to amoeboid-like motility style. Cancer Res. 2009, 69, 2072–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]





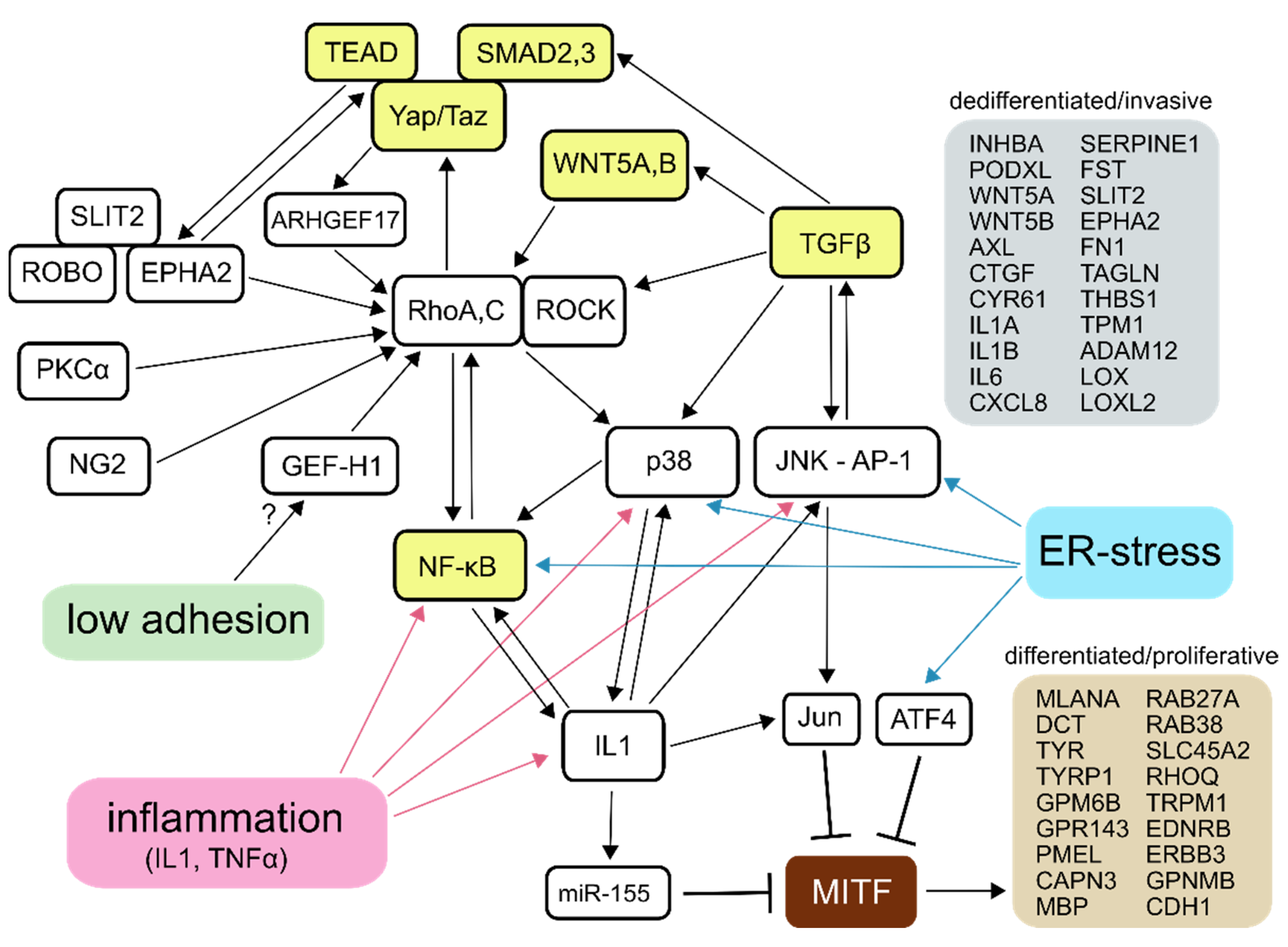{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinase | SB202190 | BIRB796 |
|---|---|---|
| p38alpha | xx/+++ | xx/++++ |
| p38beta | xx/++ | x/ |
| p38gamma | x/+ | |
| p38delta | x | |
| JNK2 | x/+ | xx/++ |
| JNK3 | x/++ | |
| NLK | x/++ | |
| RIPK2/RIP2 | xx/+ | |
| GAK | xx/++ | |
| CK1delta | x/++ | |
| BRAF | + | |
| GSK3beta | x | |
| CK1epsilon | + | |
| Lck | x | |
| ACVR1B | + | |
| CIT | + | |
| CDC42BPG | + | |
| EGFR | + | |
| PRKACB | + | |
| RPS6KA1 | + | |
| RPS6KA6 | + | |
| STK36 | + | |
| DDR1 | ++ | |
| TIE1 | ++ | |
| MAP4K4 | + | |
| STK10 | + | |
| SLK | + | |
| ABL1 | + | |
| DDR2 | + | |
| TIE2 | + | |
| RSK1 | x | |
| RSK2 | x | |
| BRSK2 | x |
| Gene | SB202190 | BIRB796 |
|---|---|---|
| TRPM1 | 4.00 ± 0.67 | |
| DCT | 3.38 ± 0.09 | 1.37 ± 0.09 |
| MLANA | 2.47 ± 0.18 | 0.94 ± 0.19 |
| GPM6B | 2.07 ± 0.08 | 1.00 ± 0.08 |
| PMEL | 1.58 ± 0.27 | |
| TYR | 1.39 ± 0.08 | |
| MBP | 1.17 ± 0.34 | 1.52 ± 0.34 |
| RAB27A | 0.95 ± 0.11 | |
| CAPN3 | 0.88 ± 0.13 | |
| GPNMB | 0.80 ± 0.07 | |
| IL1B | −4.48 ± 0.33 | −3.06 ± 0.29 |
| IL1A | −3.80 ± 0.18 | −1.95 ± 0.15 |
| CXCL8 | −3.60 ± 0.18 | −2.01 ± 0.15 |
| SERPINE1 | −3.07 ± 0.58 | |
| PODXL | −2.83 ± 0.16 | −1.36 ± 0.15 |
| AXL | −2.70 ± 0.41 | −1.11 ± 0.34 |
| INHBA | −2.24 ± 0.12 | −0.88 ± 0.12 |
| FN1 | −1.91 ± 0.09 | −1.23 ± 0.09 |
| LOXL2 | −1.81 ± 0.10 | −0.78 ± 0.10 |
| FST | −1.61 ± 0.11 | −1.13 ± 0.11 |
| ADAM12 | −1.34 ± 0.10 | −0.60 ± 0.10 |
| WNT5B | −0.82 ± 0.35 | |
| WNT5A | −0.80 ± 0.11 | |
| THBS1 | −0.73 ± 0.11 |
| Database | Data | SB202190 | BIRB796 |
|---|---|---|---|
| NCI-60 cancer cell line panel vs. upregulated genes | UACC257 | 1.83 × 10−10 | 1.33 × 10−4 |
| SKMEL5 | 1.83 × 10−10 | 7.06 × 10−2 | |
| SKMEL28 | 1.46 × 10−6 | 2.49 × 10−3 | |
| MALME 3M | 4.46 × 10−5 | 1.85 × 10−2 | |
| M14 | 5.64 × 10−3 | 1.07 × 10−1 | |
| GO—Biological Process vs. downregulated genes | extracellular matrix organization (GO:0030198) | 3.44 × 10−20 | 2.27 × 10−11 |
| regulation of cell proliferation (GO:0042127) | 2.79 × 10−11 | 2.09 × 10−5 | |
| regulation of apoptotic process (GO:0042981) | 4.35 × 10−11 | 1.38 × 10−9 | |
| regulation of cell migration (GO:0030334) | 6.14 × 10−11 | 4.86 × 10−7 | |
| regulation of angiogenesis (GO:0045765) | 8.39 × 10−9 | 3.34 × 10−5 | |
| negative regulation of programmed cell death (GO:0043069) | 9.99 × 10−9 | 3.34 × 10−5 | |
| cellular response to cytokine stimulus (GO:0071345) | 1.86 × 10−8 | 2.11 × 10−4 | |
| positive regulation of angiogenesis (GO:0045766) | 3.98 × 10−8 | 6.02 × 10−5 | |
| regulation of signal transduction (GO:0009966) | 3.98 × 10−8 | 6.02 × 10−5 | |
| negative regulation of apoptotic process (GO:0043066) | 7.51 × 10−8 | 2.06 × 10−5 | |
| positive regulation of cell migration (GO:0030335) | 7.51 × 10−8 | 7.08 × 10−5 | |
| regulation of MAPK cascade (GO:0043408) | 2.39 × 10−7 | 6.17 × 10−5 | |
| TRRUST vs. downregulated genes | NFKB1 human | 8.76 × 10−9 | 1.85 × 10−6 |
| RELA human | 3.55 × 10−8 | 1.08 × 10−5 | |
| NFKB1 mouse | 7.76 × 10−8 | 1.01 × 10−11 | |
| VHL human | 5.70 × 10−7 | 5.01 × 10−4 | |
| STAT3 mouse | 7.85 × 10−7 | 1.08 × 10−5 | |
| SP1 mouse | 8.33 × 10−7 | 1.07 × 10−11 | |
| EGR1 mouse | 2.54 × 10−6 | 3.75 × 10−6 | |
| ETS1 human | 2.72 × 10−6 | 4.92 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čermák, V.; Škarková, A.; Merta, L.; Kolomazníková, V.; Palušová, V.; Uldrijan, S.; Rösel, D.; Brábek, J. RNA-seq Characterization of Melanoma Phenotype Switch in 3D Collagen after p38 MAPK Inhibitor Treatment. Biomolecules 2021, 11, 449. https://doi.org/10.3390/biom11030449
Čermák V, Škarková A, Merta L, Kolomazníková V, Palušová V, Uldrijan S, Rösel D, Brábek J. RNA-seq Characterization of Melanoma Phenotype Switch in 3D Collagen after p38 MAPK Inhibitor Treatment. Biomolecules. 2021; 11(3):449. https://doi.org/10.3390/biom11030449
Chicago/Turabian StyleČermák, Vladimír, Aneta Škarková, Ladislav Merta, Veronika Kolomazníková, Veronika Palušová, Stjepan Uldrijan, Daniel Rösel, and Jan Brábek. 2021. "RNA-seq Characterization of Melanoma Phenotype Switch in 3D Collagen after p38 MAPK Inhibitor Treatment" Biomolecules 11, no. 3: 449. https://doi.org/10.3390/biom11030449
APA StyleČermák, V., Škarková, A., Merta, L., Kolomazníková, V., Palušová, V., Uldrijan, S., Rösel, D., & Brábek, J. (2021). RNA-seq Characterization of Melanoma Phenotype Switch in 3D Collagen after p38 MAPK Inhibitor Treatment. Biomolecules, 11(3), 449. https://doi.org/10.3390/biom11030449