Influence of Dopamine on Fluorescent Advanced Glycation End Products Formation Using Drosophila melanogaster


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Standard Calibrator fAGEs-BSA Synthesis and Characterization
2.1.1. Chemicals and Reagents
2.1.2. Standard Calibrator fAGEs-BSA Hydrothermal Synthesis and Fluorescence Characterization
2.1.3. fAGEs-BSA Calibration Curve
2.1.4. Influence of Monoamines, Psychostimulants and Antioxidant on fAGEs-BSA In Vitro Formation
2.2. In Vivo Experiments
2.2.1. Chemicals and Reagents
2.2.2. Fly Husbandry and Strains
2.2.3. Oral Administration of Substrates
2.2.4. Sample Preparation for fAGEs Determination
2.3. Statistical Analysis
3. Results
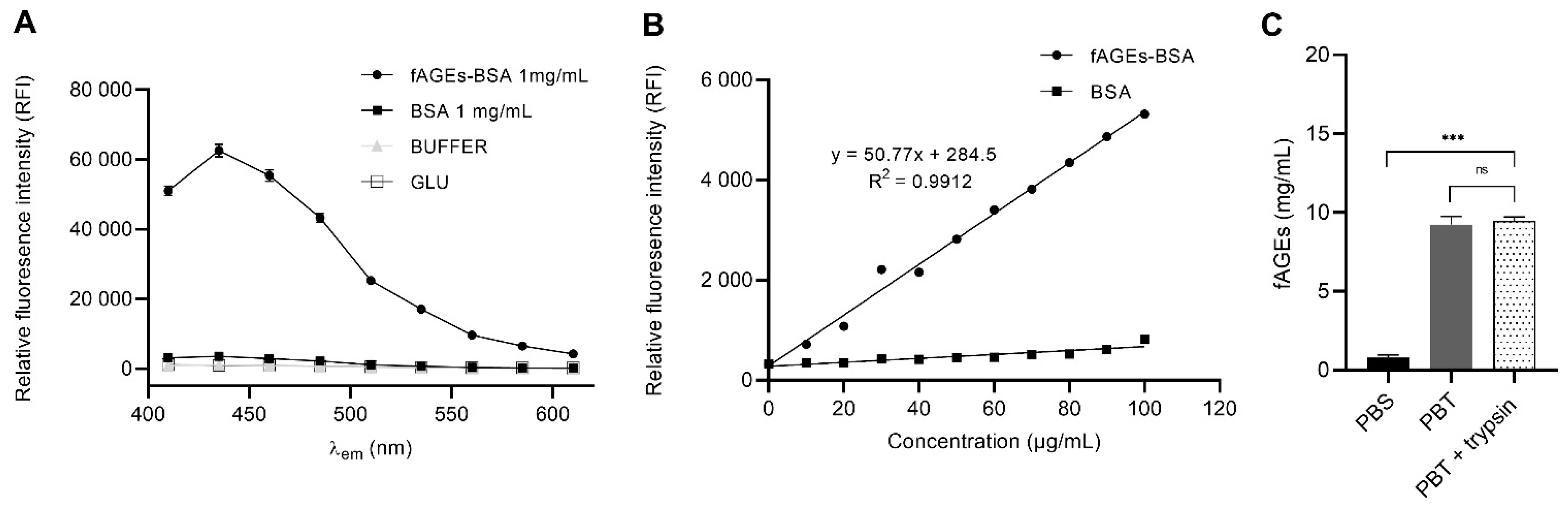
3.1. In Vitro fAGEs-BSA Hydrothermal Synthesis as Standard Calibrator for In Vivo fAGEs Quantification
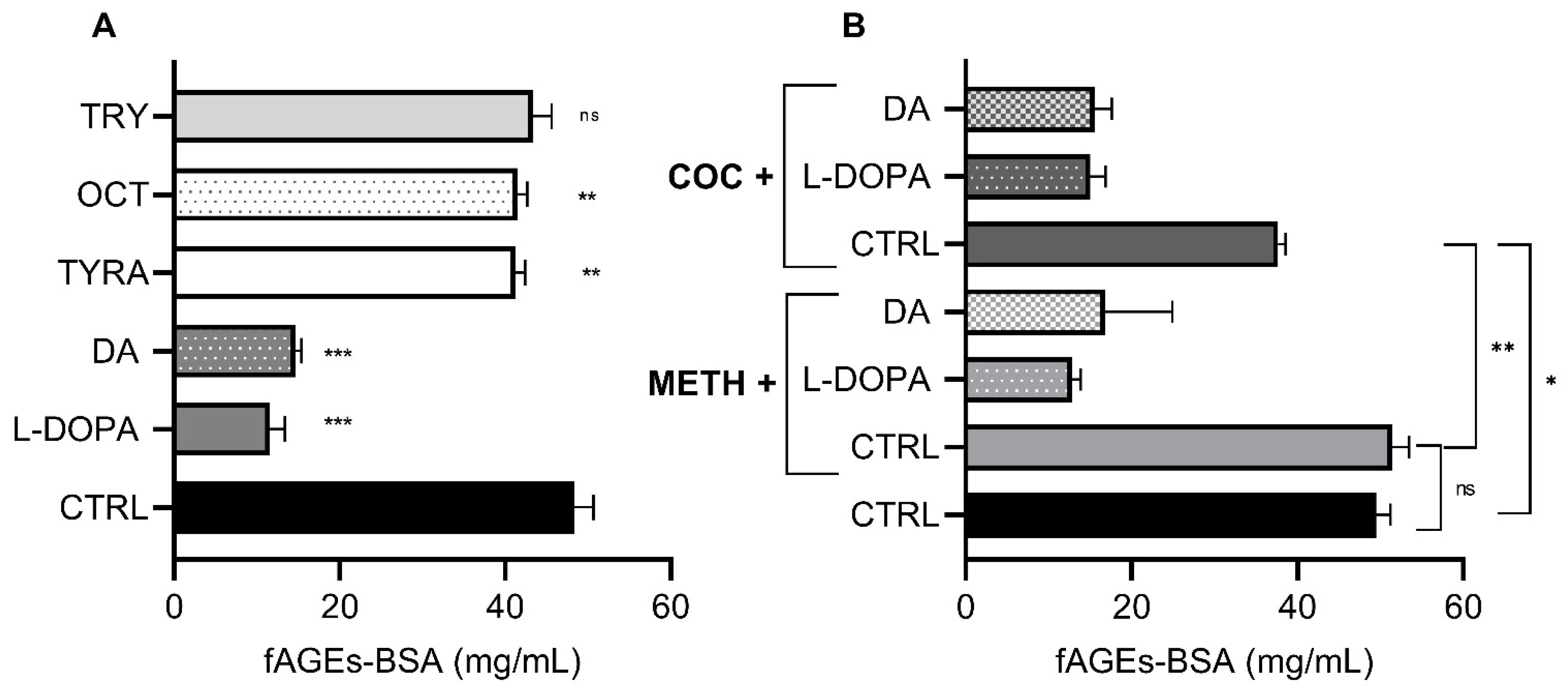
3.2. Effect of Monoamines and Psychostimulants on In Vitro fAGEs-BSA Formation
3.3. Effect of DA Synthesis and Signaling on In Vivo fAGEs Formation
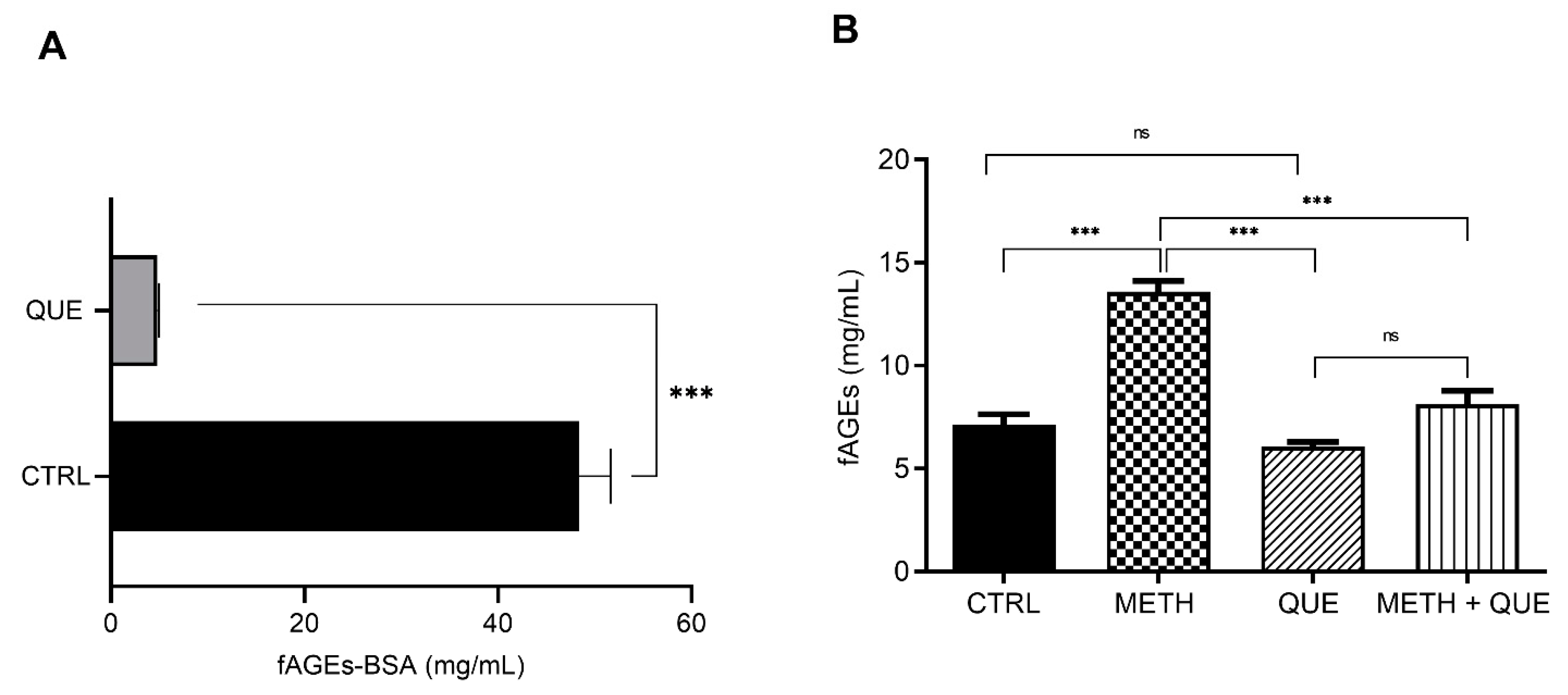
3.4. Influence of Quercetin on In Vivo fAGEs Formation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jacobson, J.; Lambert, A.J.; Portero-Otín, M.; Pamplona, R.; Magwere, T.; Miwa, S.; Driege, Y.; Brand, M.D.; Partridge, L. Biomarkers of aging in Drosophila. Aging Cell 2010, 9, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Oudes, A.J.; Herr, C.M.; Olsen, Y.; Fleming, J.E. Age-dependent accumulation of advanced glycation end-products in adult Drosophila melanogaster. Mech. Ageing Dev. 1998, 100, 221–229. [Google Scholar] [CrossRef]
- Rowan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of advanced glycation end-products in age-related diseases. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef]
- Sharma, A.; Weber, D.; Raupbach, J.; Dakal, T.C.; Fließbach, K.; Ramirez, A.; Grune, T.; Wüllner, U. Advanced glycation end products and protein carbonyl levels in plasma reveal sex-specific differences in Parkinson’s and Alzheimer’s disease. Redox Biol. 2020, 34, 101546. [Google Scholar] [CrossRef]
- Traverso, N.; Menini, S.; Cottalasso, D.; Odetti, P.; Marinari, U.M.; Pronzato, M.A. Mutual interaction between glycation and oxidation during non-enzymatic protein modification. Biochim. Biophys. Acta 1997, 1336, 409–418. [Google Scholar] [CrossRef]
- Münch, G.; Gerlach, M.; Sian, J.; Wong, A.; Riederer, P. Advanced glycation end products in neurodegeneration: More than early markers of oxidative stress? Ann. Neurol. 1998, 44, S85–S88. [Google Scholar] [CrossRef]
- Yamashita, H.; Fukushima, E.; Shimomura, K.; Hirose, H.; Nakayama, K.; Orimo, N.; Mao, W.; Katsuta, N.; Nishimon, S.; Ohnuma, T. Use of skin advanced glycation end product levels measured using a simple noninvasive method as a biological marker for the diagnosis of neuropsychiatric diseases. Int. J. Methods Psychiatr. Res. 2020, 29, e1824. [Google Scholar] [CrossRef]
- Gkogkolou, P.; Böhm, M. Advanced glycation end products. Dermatoendocrinology 2012, 4, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [Green Version]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxidative Med. Cell. Longev. 2020, 2020, 3818196. Available online: https://www.hindawi.com/journals/omcl/2020/3818196/?msclkid=6d11cf3fcf021a95c0d5902d02afe0f7 (accessed on 6 October 2020). [CrossRef] [Green Version]
- Ravichandran, G.; Lakshmanan, D.K.; Raju, K.; Elangovan, A.; Nambirajan, G.; Devanesan, A.A.; Thilagar, S. Food advanced glycation end products as potential endocrine disruptors: An emerging threat to contemporary and future generation. Environ. Int. 2019, 123, 486–500. [Google Scholar] [CrossRef]
- Chaudhuri, J.; Bains, Y.; Guha, S.; Kahn, A.; Hall, D.; Bose, N.; Gugliucci, A.; Kapahi, P. The role of advanced glycation end products in aging and metabolic diseases: Bridging association and causality. Cell Metab. 2018, 28, 337–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, M.; Favilla, R.; Strömberg, T. Assessment of advanced glycated end product accumulation in skin using auto fluorescence multispectral imaging. Comput. Biol. Med. 2017, 85, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Aragno, M.; Mastrocola, R. Dietary Sugars and Endogenous Formation of Advanced Glycation Endproducts: Emerging Mechanisms of Disease. Nutrients 2017, 9, 385. [Google Scholar] [CrossRef] [Green Version]
- Snelson, M.; Coughlan, M.T. Dietary Advanced Glycation End Products: Digestion, Metabolism and Modulation of Gut Microbial Ecology. Nutrients 2019, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, K.M.; Qais, F.A.; Hasan, H.; Naseem, I. Anti-diabetic study of vitamin B6 on hyperglycaemia induced protein carbonylation, DNA damage and ROS production in alloxan induced diabetic rats. Toxicol Res. (Camb) 2019, 8, 568–579. [Google Scholar] [CrossRef]
- Alam, M.M.; Ahmad, I.; Naseem, I. Inhibitory effect of quercetin in the formation of advance glycation end products of human serum albumin: An in vitro and molecular interaction study. Int. J. Biol. Macromol. 2015, 79, 336–343. [Google Scholar] [CrossRef]
- Gupta, A.; Birhman, K.; Raheja, I.; Sharma, S.K.; Kar, H.K. Quercetin: A wonder bioflavonoid with therapeutic potential in disease management. Asian Pac. J. Trop. Dis. 2016, 6, 248–252. [Google Scholar] [CrossRef]
- Huttunen, H.J.; Kuja-Panula, J.; Rauvala, H. Receptor for Advanced Glycation End Products (RAGE) Signaling Induces CREB-dependent Chromogranin Expression during Neuronal Differentiation. J. Biol. Chem. 2002, 277, 38635–38646. [Google Scholar] [CrossRef] [Green Version]
- Krautwald, M.; Münch, G. Advanced glycation end products as biomarkers and gerontotoxins—A basis to explore methylglyoxal-lowering agents for Alzheimer’s disease? Exp. Gerontol. 2010, 45, 744–751. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 3085756. Available online: https://www.hindawi.com/journals/omcl/2019/3085756/ (accessed on 16 November 2020). [CrossRef] [Green Version]
- Teismann, P.; Sathe, K.; Bierhaus, A.; Leng, L.; Martin, H.L.; Bucala, R.; Weigle, B.; Nawroth, P.P.; Schulz, J.B. Receptor for advanced glycation endproducts (RAGE) deficiency protects against MPTP toxicity. Neurobiol. Aging 2012, 33, 2478–2490. [Google Scholar] [CrossRef] [Green Version]
- Viana, S.D.; Valero, J.; Rodrigues-Santos, P.; Couceiro, P.; Silva, A.M.; Carvalho, F.; Ali, S.F.; Fontes-Ribeiro, C.A.; Pereira, F.C. Regulation of striatal astrocytic receptor for advanced glycation end-products variants in an early stage of experimental Parkinson’s disease. J. Neurochem. 2016, 138, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Cassar, M.; Issa, A.R.; Riemensperger, T.; Petitgas, C.; Rival, T.; Coulom, H.; Iché-Torres, M.; Han, K.; Birman, S. A dopamine receptor contributes to paraquat-induced neurotoxicity in Drosophila. Hum. Mol. Genet. 2015, 24, 197–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Oliveira, T.; Rego, A.; Oliveira, C. Oxidative Stress and Drugs of Abuse: An Update. MROC 2013, 10, 321–334. [Google Scholar] [CrossRef]
- Riemensperger, T.; Isabel, G.; Coulom, H.; Neuser, K.; Seugnet, L.; Kume, K.; Iché-Torres, M.; Cassar, M.; Strauss, R.; Preat, T.; et al. Behavioral consequences of dopamine deficiency in the Drosophila central nervous system. Proc. Natl. Acad. Sci. USA 2011, 108, 834–839. [Google Scholar] [CrossRef] [Green Version]
- Wise, R.A.; Robble, M.A. Dopamine and Addiction. Annu. Rev. Psychol. 2020, 71, 79–106. [Google Scholar] [CrossRef] [PubMed]
- Treweek, J.B.; Dickerson, T.J.; Janda, K.D. Drugs of Abuse That Mediate Advanced Glycation End Product Formation: A Chemical Link to Disease Pathology. Acc. Chem. Res. 2009, 42, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Kasture, A.; Hummel, T.; Sucic, S.; Freissmuth, M. Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems. IJMS 2018, 19, 1788. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Ho, M.C.W.; Liu, Q.; Horiuchi, W.; Lin, C.; Task, D.; Luan, H.; White, B.H.; Potter, C.J.; Wu, M.N. A Genetic Toolkit for Dissecting Dopamine Circuit Function in Drosophila. Cell Rep. 2018, 23, 652–665. [Google Scholar] [CrossRef] [Green Version]
- Lüscher, C.; Malenka, R.C. Drug-Evoked Synaptic Plasticity in Addiction: From Molecular Changes to Circuit Remodeling. Neuron 2011, 69, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Seto, E.S. Dopamine dynamics and signaling in Drosophila: An overview of genes, drugs and behavioral paradigms. Exp. Anim. 2014, 63, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, S.; Kodama, L.; Driscoll, M.R.; Wu, M.N. Two Dopaminergic Neurons Signal to the Dorsal Fan-Shaped Body to Promote Wakefulness in Drosophila. Curr. Biol. 2012, 22, 2114–2123. [Google Scholar] [CrossRef] [Green Version]
- Bhatwadekar, A.D.; Ghole, V.S. Rapid method for the preparation of an AGE-BSA standard calibrator using thermal glycation. J. Clin. Lab. Anal. 2005, 19, 11–15. [Google Scholar] [CrossRef]
- Sharma, S.D.; Pandey, B.N.; Mishra, K.P.; Sivakami, S. Amadori product and age formation during nonenzymatic glycosylation of bovine serum albumin in vitro. J. Biochem. Mol. Biol. Biophys. Off. J. Fed. Asian Ocean. Biochem. Mol. Biol. (FAOBMB) 2002, 6, 233–242. [Google Scholar] [CrossRef]
- Kume, K. Dopamine Is a Regulator of Arousal in the Fruit Fly. J. Neurosci. 2005, 25, 7377–7384. [Google Scholar] [CrossRef] [Green Version]
- Faville, R.; Kottler, B.; Goodhill, G.J.; Shaw, P.J.; van Swinderen, B. How deeply does your mutant sleep? Probing arousal to better understand sleep defects in Drosophila. Sci. Rep. 2015, 5, 8454. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wang, Y.; Li, Q.; Zhong, Y.; Chen, L.; Du, Y.; He, J.; Liao, L.; Xiong, K.; Yi, C.; et al. The Main Molecular Mechanisms Underlying Methamphetamine- Induced Neurotoxicity and Implications for Pharmacological Treatment. Front. Mol. Neurosci. 2018, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Huenchuguala, S.; Paris, I.; Segura-Aguilar, J. Dopamine Oxidation and Autophagy. Parkinson’s Dis. 2012, 2012, 920953. Available online: https://www.hindawi.com/journals/pd/2012/920953/ (accessed on 20 January 2021). [CrossRef]
- Bainton, R.J.; Tsai, L.T.-Y.; Singh, C.M.; Moore, M.S.; Neckameyer, W.S.; Heberlein, U. Dopamine modulates acute responses to cocaine, nicotine and ethanol in Drosophila. Curr. Biol. 2000, 10, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Yen, G.-C.; Hsieh, C.-L. Antioxidant Effects of Dopamine and Related Compounds. Biosci. Biotechnol. Biochem. 1997, 61, 1646–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnier, V.; Cerami, A. Nonenzymatic browning in vivo: Possible process for aging of long-lived proteins. Science 1981, 211, 491–493. [Google Scholar] [CrossRef]
- Kannari, K.; Tanaka, H.; Maeda, T.; Tomiyama, M.; Suda, T.; Matsunaga, M. Reserpine pretreatment prevents increases in extracellular striatal dopamine following L-DOPA administration in rats with nigrostriatal denervation. J. Neurochem. 2000, 74, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Omiatek, D.M.; Bressler, A.J.; Cans, A.-S.; Andrews, A.M.; Heien, M.L.; Ewing, A.G. The real catecholamine content of secretory vesicles in the CNS revealed by electrochemical cytometry. Sci. Rep. 2013, 3, 1447. [Google Scholar] [CrossRef] [Green Version]
- Sulzer, D.; Cragg, S.J.; Rice, M.E. Striatal dopamine neurotransmission: Regulation of release and uptake. Basal Ganglia 2016, 6, 123–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulvihill, K.G. Presynaptic regulation of dopamine release: Role of the DAT and VMAT2 transporters. Neurochem. Int. 2019, 122, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Paris, G.; Kraszewski, S.; Ramseyer, C.; Enescu, M. About the structural role of disulfide bridges in serum albumins: Evidence from protein simulated unfolding. Biopolymers 2012, 97, 889–898. [Google Scholar] [CrossRef]
- Hull, H.H.; Chang, R.; Kaplan, L.J. On the location of the sulfhydryl group in bovine plasma albumin. Biochim. Biophys. Acta 1975, 400, 132–136. [Google Scholar] [CrossRef]
- Abha, K.; Sumithra, I.S.; Suji, S.; Anjana, R.R.; Devi, J.S.A.; Nebu, J.; Lekha, G.M.; Aparna, R.S.; George, S. Dopamine-induced photoluminescence quenching of bovine serum albumin–capped manganese-doped zinc sulphide quantum dots. Anal. Bioanal. Chem. 2020, 412, 5671–5681. [Google Scholar] [CrossRef]
- LaVoie, M.J.; Hastings, T.G. Dopamine Quinone Formation and Protein Modification Associated with the Striatal Neurotoxicity of Methamphetamine: Evidence against a Role for Extracellular Dopamine. J. Neurosci. 1999, 19, 1484–1491. [Google Scholar] [CrossRef]
- Pereira, S.P.; Cunha-Oliveira, T. Role of Mitochondria on the Neurological Effects of Cocaine. In The Neuroscience of Cocaine; Elsevier: Amsterdam, The Netherlands, 2017; pp. 205–218. [Google Scholar]
- Deng, S.-X.; Bharat, N.; Fischman, M.C.; Landry, D.W. Covalent modification of proteins by cocaine. Proc. Natl. Acad. Sci. USA 2002, 99, 3412–3416. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Ka, H.; Ml, H.; Ag, E. Using in vivo Electrochemistry to Study the Physiological Effects of Cocaine and Other Stimulants on the Drosophila melanogaster Dopamine Transporter. ACS Chem. Neurosci. 2010, 1, 74–83. [Google Scholar] [CrossRef]
- Makos, M.A.; Kim, Y.-C.; Han, K.-A.; Heien, M.L.; Ewing, A.G. in vivo Electrochemical Measurements of Exogenously Applied Dopamine in Drosophila melanogaster. Anal. Chem. 2009, 81, 1848–1854. [Google Scholar] [CrossRef] [Green Version]
- Hanna, M.E.; Bednářová, A.; Rakshit, K.; Chaudhuri, A.; O’Donnell, J.M.; Krishnan, N. Perturbations in dopamine synthesis lead to discrete physiological effects and impact oxidative stress response in Drosophila. J. Insect Physiol. 2015, 73, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Ma, J.; Chen, F.; Wang, M. Naturally occurring inhibitors against the formation of advanced glycation end-products. Food Funct. 2011, 2, 289. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filošević Vujnović, A.; Jović, K.; Pištan, E.; Andretić Waldowski, R. Influence of Dopamine on Fluorescent Advanced Glycation End Products Formation Using Drosophila melanogaster. Biomolecules 2021, 11, 453. https://doi.org/10.3390/biom11030453
Filošević Vujnović A, Jović K, Pištan E, Andretić Waldowski R. Influence of Dopamine on Fluorescent Advanced Glycation End Products Formation Using Drosophila melanogaster. Biomolecules. 2021; 11(3):453. https://doi.org/10.3390/biom11030453
Chicago/Turabian StyleFilošević Vujnović, Ana, Katarina Jović, Emanuel Pištan, and Rozi Andretić Waldowski. 2021. "Influence of Dopamine on Fluorescent Advanced Glycation End Products Formation Using Drosophila melanogaster" Biomolecules 11, no. 3: 453. https://doi.org/10.3390/biom11030453
APA StyleFilošević Vujnović, A., Jović, K., Pištan, E., & Andretić Waldowski, R. (2021). Influence of Dopamine on Fluorescent Advanced Glycation End Products Formation Using Drosophila melanogaster. Biomolecules, 11(3), 453. https://doi.org/10.3390/biom11030453