
Viscoelastic Response of Neurofilaments: An Atomistic Simulation Approach


Abstract
:1. Introduction
2. Method
2.1. MD Simulation Scheme
2.2. Viscoelastic Modeling of NF
- (a)
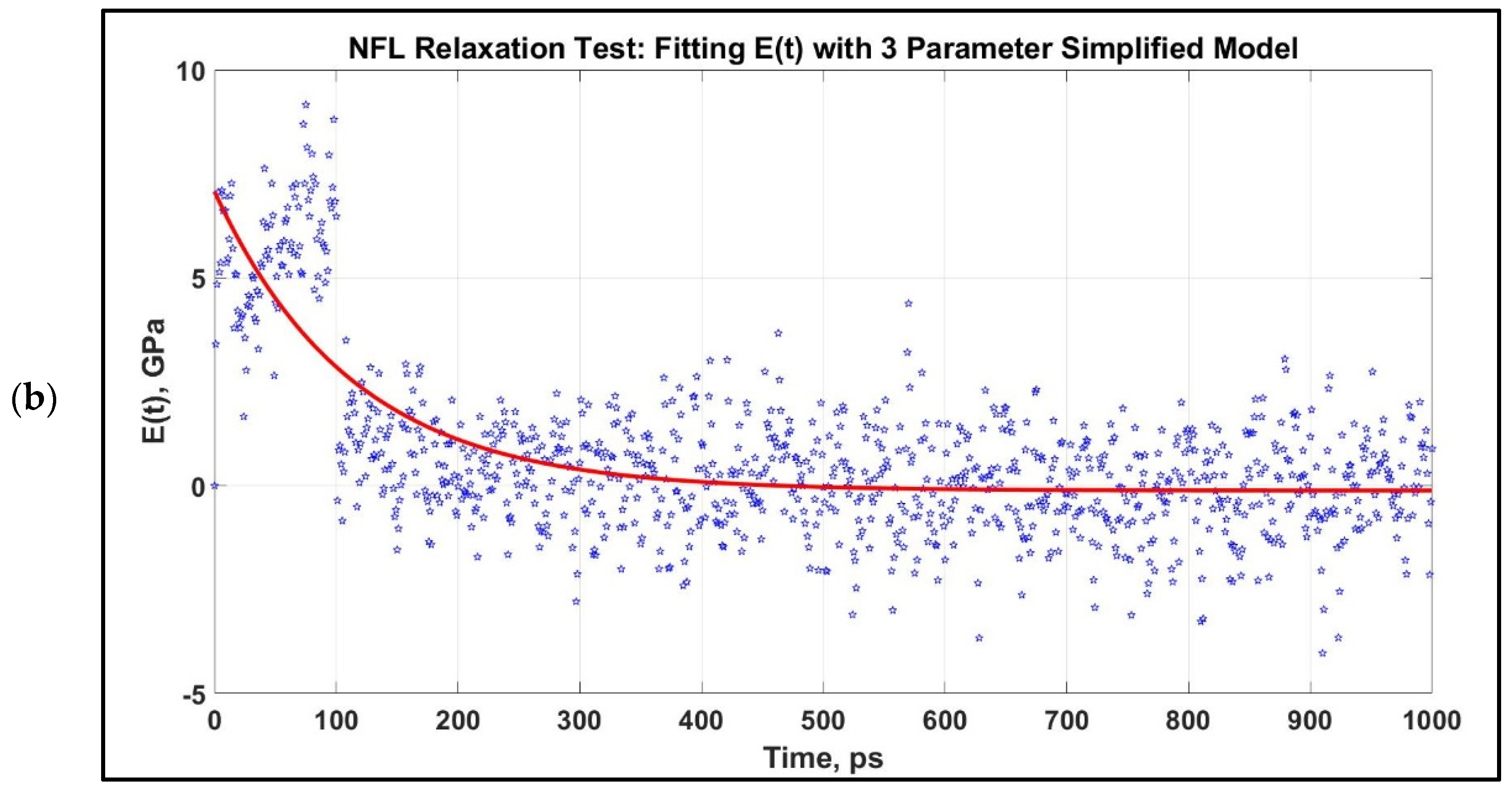
- Obtain E(t),
- (b)
- Obtain E(s)*, which is the expression of E at the Laplacian domain,
- (c)
- Calculate G(s)* and K(s)* using Equations (3) and (4) (these are the expression of G and K at the Laplacian domain, respectively),
- (d)
- Use inverse Laplace calculation to obtain G(t) and K(t).
- Determining the NF-L and NF-H structures from i-TASSER (pdb files),
- Using CHARMM-GUI (quick MD simulator or solution builder module) for creating the simulation box, solvating the system along with neutralization by the necessary number of ions, and creating LAMMPS readable data file for simulation,
- Creating input scripts to run simulation in LAMMPS (equilibration and tensile test),
- Running equilibration for energy minimization in LAMMPS,
- Running tensile test in LAMMPS,
- Post-processing (calculating stress and strain from the log file, plotting the data in MATLAB, fitting the stress vs. strain data to required expression by using fminsearch function of MATLAB, creating snapshots of the simulation by using OVITO modifiers).
3. Results
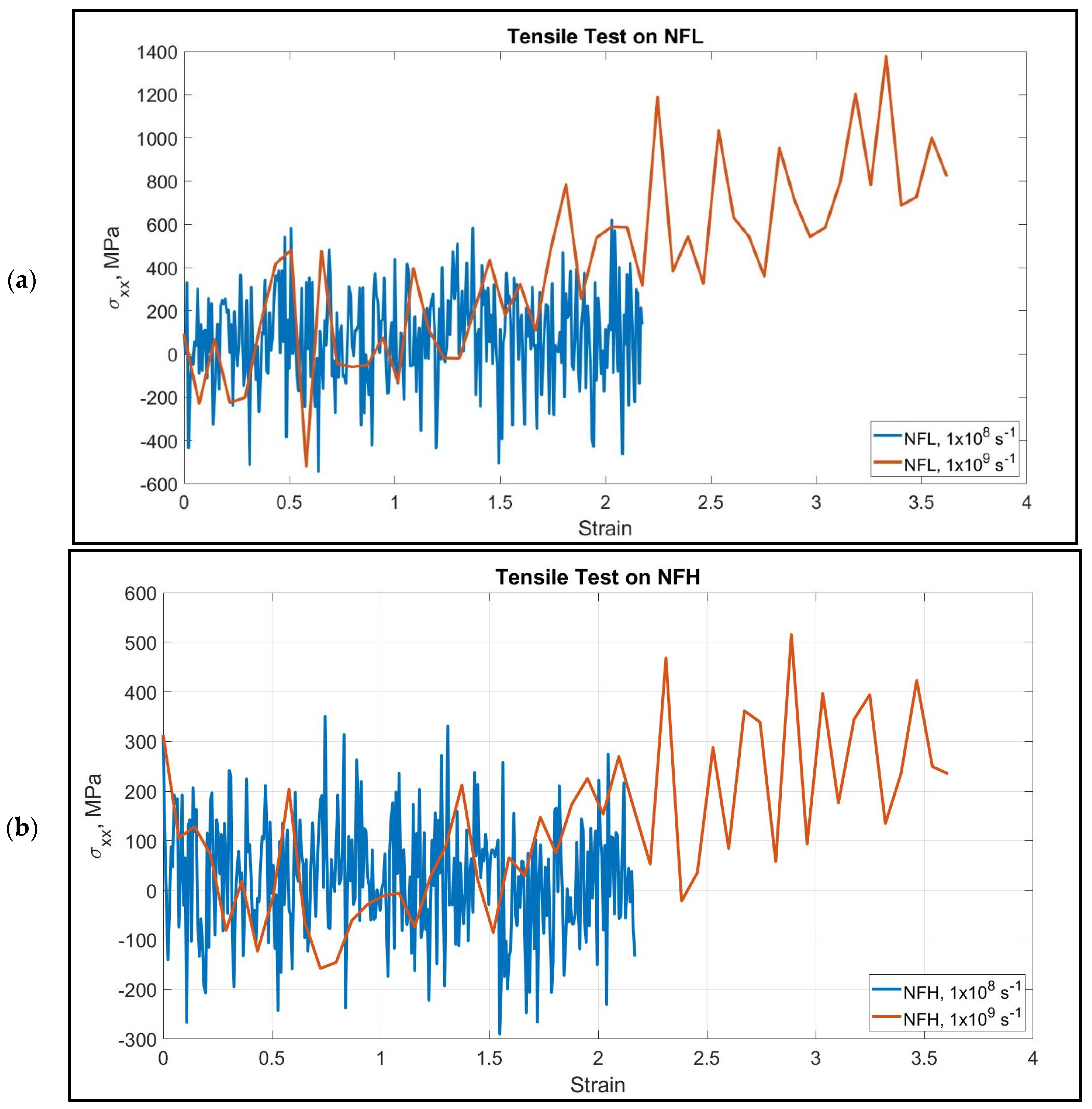
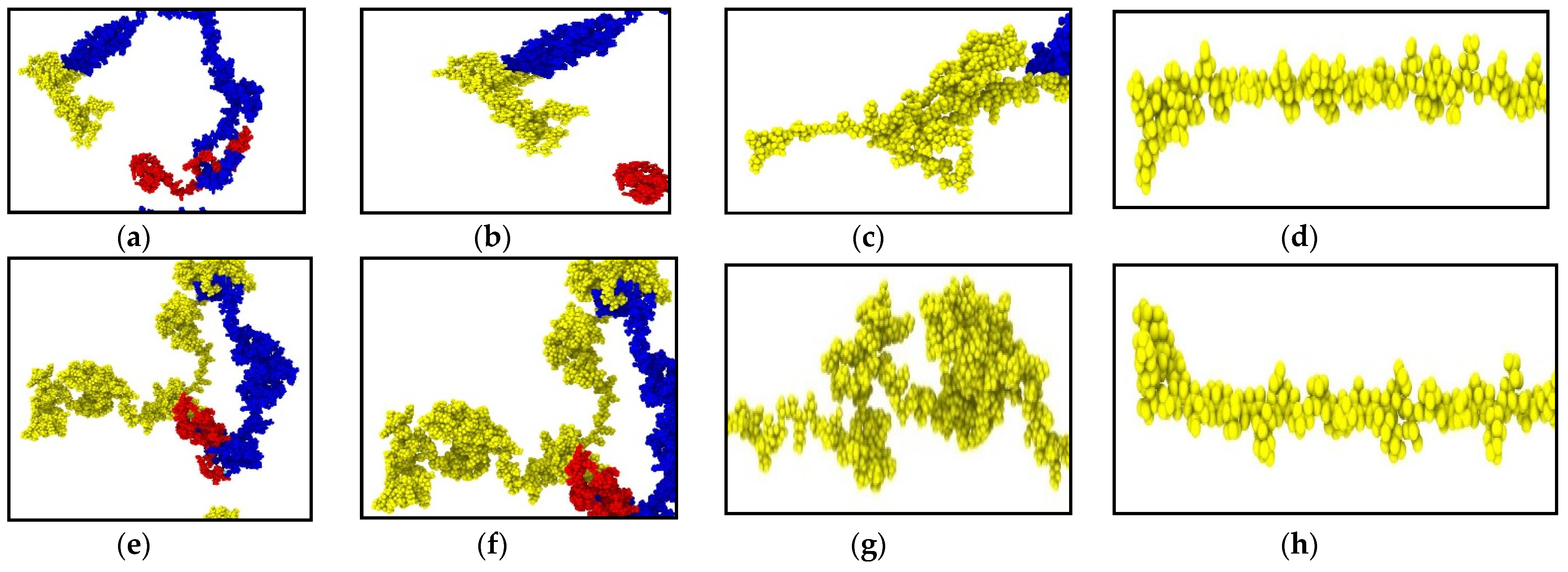
3.1. Neurofilament Deformation
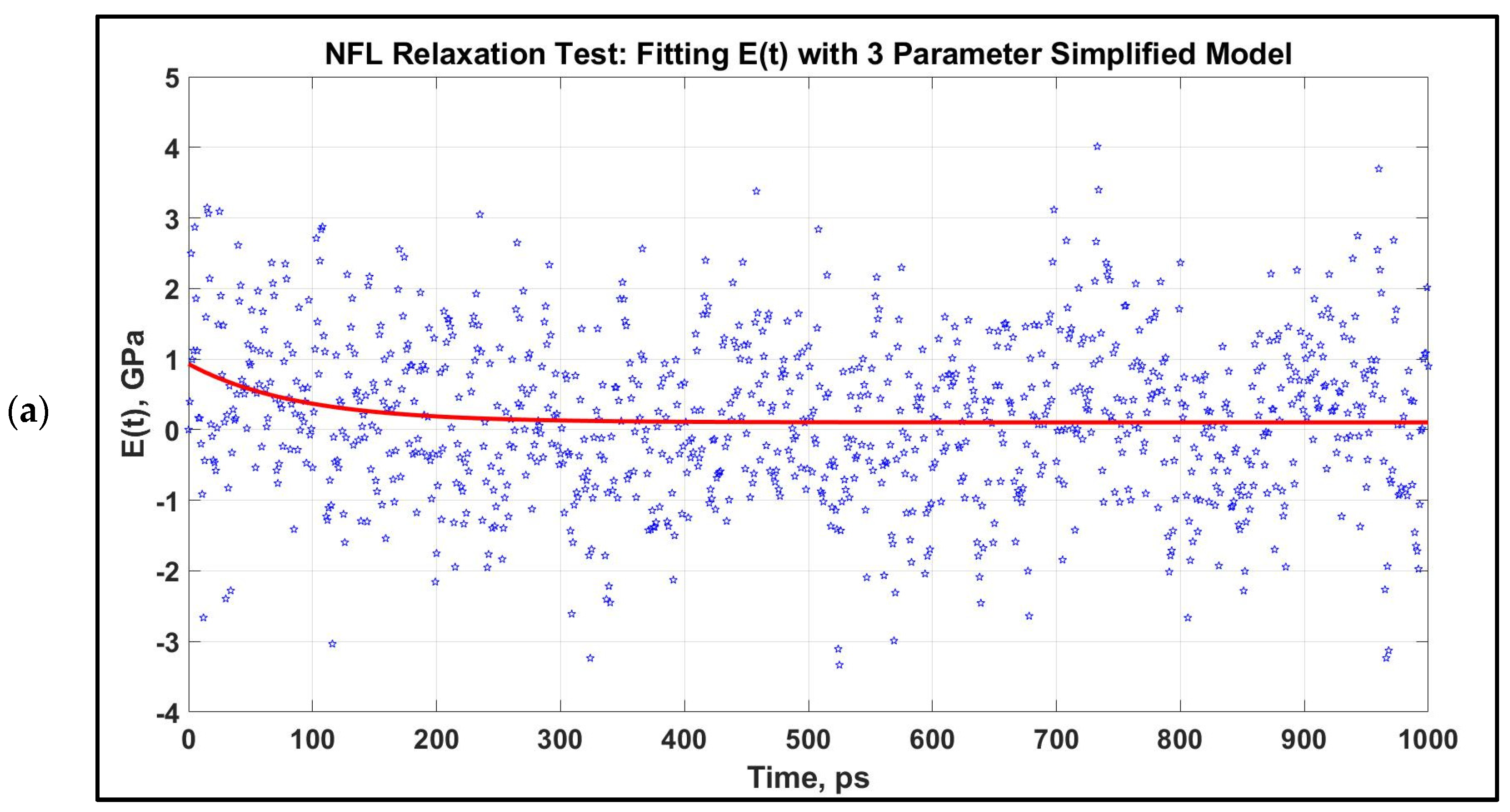
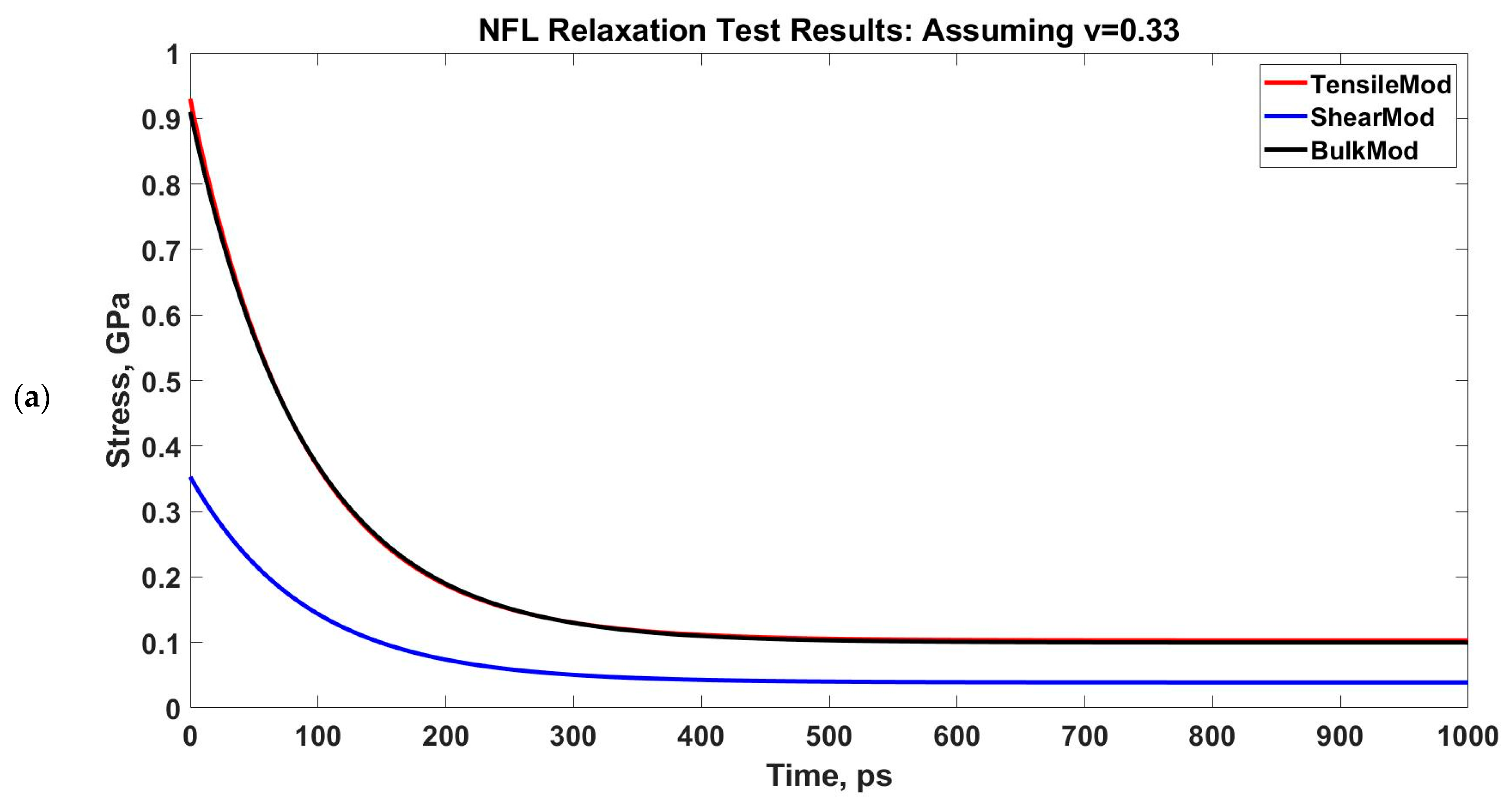
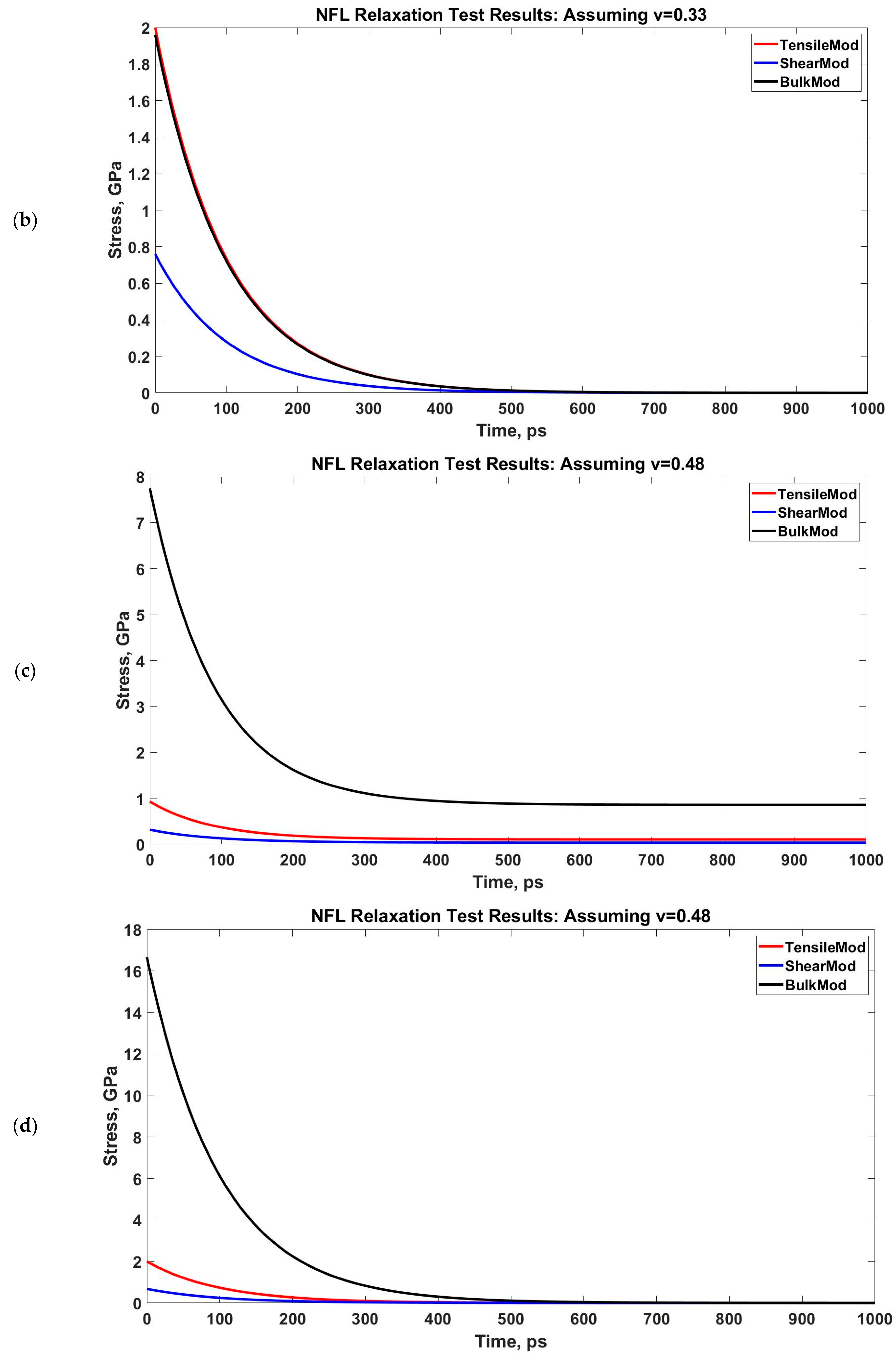
3.2. Viscoelastic Modeling of Neurofilaments
4. Discussion
4.1. Strain Rate Dependence of Neurofilaments
4.2. Viscoelastic Behavior of Neurofilaments
5. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
MSSFSYEPYYSTSYKRRYVETPRVHISSVRSGYSTARSAYSSYSAPVSSSLSVRRSYSSS SGSLMPSLENLDLSQVAAISNDLKSIRTQEKAQLQDLNDRFASFIERVHELEQQNKVLEA ELLVLRQKHSEPSRFRALYEQEIRDLRLAAEDATNEKQALQGEREGLEETLRNLQARYEE EVLSREDAEGRLMEARKGADEAALARAELEKRIDSLMDEISFLKKVHEEEIAELQAQIQY AQISVEMDVTKPDLSAALKDIRAQYEKLAAKNMQNAEEWFKSRFTVLTESAAKNTDAVRA AKDEVSESRRLLKAKTLEIEACRGMNEALEKQLQELEDKQNADISAMQDTINKLENELRT TKSEMARYLKEYQDLLNVKMALDIEIAAYRKLLEGEETRLSFTSVGSITSGYSQSSQVFG RSAYGGLQTSSYLMSTRSFPSYYTSHVQEEQIEVEETIEAAKAEEAKDEPPSEGEAEEEE KDKEEAEEEEAAEEEEAAKEESEEAKEEEEGGEGEEGEETKEAEEEEKKVEGAGEEQAAK KKD
MSYTLDSLGNPSAYRRVTETRSSFSRVSGSPSSGFRSQSWSRGSPSTVSSSYKRSMLAPR LAYSSAMLSSAESSLDFSQSSSLLNGGSGPGGDYKLSRSNEKEQLQGLNDRFAGYIEKVH YLEQQNKEIEAEIQALRQKQASHAQLGDAYDQEIRELRATLEMVNHEKAQVQLDSDHLEE DIHRLKERFEEEARLRDDTEAAIRALRKDIEEASLVKVELDKKVQSLQDEVAFLRSNHEE EVADLLAQIQASHITVERKDYLKTDISTALKEIRSQLESHSDQNMHQAEEWFKCRYAKLT EAAEQNKEAIRSAKEEIAEYRRQLQSKSIELESVRGTKESLERQLSDIEERHNHDLSSYQ DTIQQLENELRGTKWEMARHLREYQDLLNVKMALDIEIAAYRKLLEGEETRFSTFAGSIT GPLYTHRPPITISSKIQKPKVEAPKLKVQHKFVEEIIEETKVEDEKSEMEEALTAITEEL AVSMKEEKKEAAEEKEEEPEAEEEEVAAKKSPVKATAPEVKEEEGEKEEEEGQEEEEEED EGAKSDQAEEGGSEKEGSSEKEEGEQEEGETEAEAEGEEAEAKEEKKVEEKSEEVATKEE LVADAKVEKPEKAKSPVPKSPVEEKGKSPVPKSPVEEKGKSPVPKSPVEEKGKSPVPKSP VEEKGKSPVSKSPVEEKAKSPVPKSPVEEAKSKAEVGKGEQKEEEEKEVKEAPKEEKVEK KEEKPKDVPEKKKAESPVKEEAVAEVVTITKSVKVHLEKETKEEGKPLQQEKEKEKAGGE GGSEEEGSDKGAKGSRKEDIAVNGEVEGKEEVEQETKEKGSGREEEKGVVTNGLDLSPAD EKKGGDKSEEKVVVTKTVEKITSEGGDGATKYITKSVTVTQKVEEHEETFEEKLVSTKKV EKVTSHAIVKEVTQSD
MMSFGGADALLGAPFAPLHGGGSLHYALARKGGAGGTRSAAGSSSGFHSWTRTSVSSVSA SPSRFRGAGAASSTDSLDTLSNGPEGCMVAVATSRSEKEQLQALNDRFAGYIDKVRQLEA HNRSLEGEAAALRQQQAGRSAMGELYEREVREMRGAVLRLGAARGQLRLEQEHLLEDIAH VRQRLDDEARQREEAEAAARALARFAQEAEAARVDLQKKAQALQEECGYLRRHHQEEVGE LLGQIQGSGAAQAQMQAETRDALKCDVTSALREIRAQLEGHAVQSTLQSEEWFRVRLDRL SEAAKVNTDAMRSAQEEITEYRRQLQARTTELEALKSTKDSLERQRSELEDRHQADIASY QEAIQQLDAELRNTKWEMAAQLREYQDLLNVKMALDIEIAAYRKLLEGEECRIGFGPIPF SLPEGLPKIPSVSTHIKVKSEEKIKVVEKSEKETVIVEEQTEETQVTEEVTEEEEKEAKE EEGKEEEGGEEEEAEGGEEETKSPPAEEAASPEKEAKSPVKEEAKSPAEAKSPEKEEAKS PAEVKSPEKAKSPAKEEAKSPPEAKSPEKEEAKSPAEVKSPEKAKSPAKEEAKSPAEAKS PEKAKSPVKEEAKSPAEAKSPVKEEAKSPAEVKSPEKAKSPTKEEAKSPEKAKSPEKAKS PEKEEAKSPEKAKSPVKAEAKSPEKAKSPVKAEAKSPEKAKSPVKEEAKSPEKAKSPVKE EAKSPEKAKSPVKEEAKTPEKAKSPVKEEAKSPEKAKSPEKAKTLDVKSPEAKTPAKEEA RSPADKFPEKAKSPVKEEVKSPEKAKSPLKEDAKAPEKEIPKKEEVKSPVKEEEKPQEVK VKEPPKKAEEEKAPATPKTEEKKDSKKEEAPKKEAPKPKVEEKKEPAVEKPKESKVEAKK EEAEDKKKVPTPEKEAPAKVEVKEDAKPKEKTEVAKKEPDDAKAKEPSKPAEKKEAAPEK KDTKEEKAKKPEEKPKTEAKAKEDDKTLSKEPSKPKAEKAEKSSSTDQKDSKPPEKATED KAAKGK
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NF-L | NF-H | |
|---|---|---|
| Number of atoms in NF | 8486 | 15,882 |
| Number of atoms in the simulation box (including water and ions) | 567,706 | 757,130 |
| Simulation box size (nm × nm × nm) | 40 × 18 × 10 | 40 × 20 × 10 |
| Approximate filament length in nm (N.B. the end-to-end distance of the filament, which is not in a straight conformation, is being mentioned here as “length” to provide an idea of the protein placement in the simulation box) | 17.3 | 17.3 |
References
- Aamodt, E.J.; Williams, R.C., Jr. Microtubule-associated proteins connect microtubules and neurofilaments in vitro. Biochemistry 1984, 23, 6023–6031. [Google Scholar] [CrossRef]
- Aamodt, E.J.; Williams, R.C., Jr. Association of microtubules and neurofilaments in vitro is not mediated by ATP. Biochemistry 1984, 23, 6031–6035. [Google Scholar] [CrossRef]
- Ahmadzadeh, H.; Smith, D.H.; Shenoy, V.B. Mechanical effects of dynamic binding between tau proteins on microtubules during axonal injury. Biophys. J. 2015, 109, 2328–2337. [Google Scholar] [CrossRef] [Green Version]
- Ahmadzadeh, H.; Smith, D.H.; Shenoy, V.B. Viscoelasticity of tau proteins leads to strain rate-dependent breaking of microtubules during axonal stretch injury: Predictions from a mathematical model. Biophys. J. 2014, 106, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranda-Espinoza, H.; Carl, P.; Leterrier, J.-F.; Janmey, P.; Discher, D.E. Domain unfolding in neurofilament sidearms: Effects of phosphorylation and ATP. FEBS Lett. 2002, 531, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Battisti, A.; Ciasca, G.; Grottesi, A.; Bianconi, A.; Tenenbaum, A. Temporary secondary structures in tau, an intrinsically disordered protein. Mol. Simul. 2012, 38, 525–533. [Google Scholar] [CrossRef]
- Battisti, A.; Tenenbaum, A. Molecular dynamics simulation of intrinsically disordered proteins. Mol. Simul. 2012, 38, 139–143. [Google Scholar] [CrossRef]
- Beck, R.; Deek, J.; Safinya, C.R. Structures and interactions in ‘bottlebrush’ neurofilaments: The role of charged disordered proteins in forming hydrogel networks. Biochem. Soc. Trans. 2012, 40, 1027–1031. [Google Scholar] [CrossRef]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; MacKerell, A.D. Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone ϕ, ψ and side-chain χ1 and χ2 dihedral angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocquet, A.; Berges, R.; Frank, R.; Robert, P.; Peterson, A.C.; Eyer, J. Neurofilaments bind tubulin and modulate its polymerization. J. Neurosci. 2009, 29, 11043–11054. [Google Scholar] [CrossRef]
- Brinson, H.F.; Brinson, L.C. Polymer Engineering Science and Viscoelasticity; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Brooks, B.R.; Brooks, C.L., III; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef]
- Brureau, A.; Blanchard-Bregeon, V.; Pech, C.; Hamon, S.; Chaillou, P.; Guillemot, J.-C.; Barneoud, P.; Bertrand, P.; Pradier, L.; Rooney, T. NF-L in cerebrospinal fluid and serum is a biomarker of neuronal damage in an inducible mouse model of neurodegeneration. Neurobiol. Dis. 2017, 104, 73–84. [Google Scholar] [CrossRef]
- Castro, T.G.; Munteanu, F.-D.; Cavaco-Paulo, A. Electrostatics of tau protein by molecular dynamics. Biomolecules 2019, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, R.; Kwak, Y.; Gebremichael, Y. Structural properties of neurofilament sidearms: Sequence-based modeling of neurofilament architecture. J. Mol. Biol. 2009, 391, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Chen, T. Determining a Prony Series for a Viscoelastic Material from Time Strain Data; US Army Res. Lab.: Fort Belvoir, VA, USA, 2000. [Google Scholar]
- Christensen, R.M. Theory of Viscoelasticity: An Introduction; Elsevier: Amsterdam, The Netherlands, 1982. [Google Scholar]
- Cifuentes-Diaz, C.; Nicole, S.; Velasco, M.E.; Borra-Cebrian, C.; Panozzo, C.; Frugier, T.; Millet, G.; Roblot, N.; Joshi, V.; Melki, J. Neurofilament accumulation at the motor endplate and lack of axonal sprouting in a spinal muscular atrophy mouse model. Hum. Mol. Genet. 2002, 11, 1439–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waegh, S.M.D.; Lee, V.M.-Y.; Brady, S.T. Local modulation of neurofilament phosphorylation, axonal caliber, and slow axonal transport by myelinating Schwann cells. Cell 1992, 68, 451–463. [Google Scholar] [CrossRef]
- Elder, G.A.; Friedrich, V.L.; Bosco, P.; Kang, C.; Gourov, A.; Tu, P.-H.; Lee, V.M.-Y.; Lazzarini, R.A. Absence of the mid-sized neurofilament subunit decreases axonal calibers, levels of light neurofilament (NF-L), and neurofilament content. J. Cell Biol. 1998, 141, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Eyer, J.; Leterrier, J.-F. Influence of the phosphorylation state of neurofilament proteins on the interactions between purified filaments in vitro. Biochem. J. 1988, 252, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Flügge, W. Viscoelasticity; Blaisdell Publ. Co.: Waltham, MA, USA, 1975. [Google Scholar]
- Fliegner, K.H.; Liem, R.K.H. Cellular and Molecular Biology of Neuronal Intermediate Filaments. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 1991; pp. 109–167. [Google Scholar]
- Garcia, M.L.; Rao, M.V.; Fujimoto, J.; Garcia, V.B.; Shah, S.B.; Crum, J.; Gotow, T.; Uchiyama, Y.; Ellisman, M.; Calcutt, N.A. Phosphorylation of highly conserved neurofilament medium KSP repeats is not required for myelin-dependent radial axonal growth. J. Neurosci. 2009, 29, 1277–1284. [Google Scholar] [CrossRef]
- Herrmann, H.; Aebi, U. Intermediate filaments: Molecular structure, assembly mechanism, and integration into functionally distinct intracellular scaffolds. Annu. Rev. Biochem. 2004, 73, 749–789. [Google Scholar] [CrossRef]
- Higham, D.J.; Higham, N.J. MATLAB Guide; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 2016. [Google Scholar]
- Hisanaga, S.; Gonda, Y.; Inagaki, M.; Ikai, A.; Hirokawa, N. Effects of phosphorylation of the neurofilament L protein on filamentous structures. Cell Regul. 1990, 1, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisanaga, S.; Hirokawa, N. The effects of dephosphorylation on the structure of the projections of neurofilament. J. Neurosci. 1989, 9, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisanaga, S.; Hirokawa, N. Structure of the peripheral domains of neurofilaments revealed by low angle rotary shadowing. J. Mol. Biol. 1988, 202, 297–305. [Google Scholar] [CrossRef]
- Hoffman, P.N.; Lasek, R.J. The slow component of axonal transport. Identification of major structural polypeptides of the axon and their generality among mammalian neurons. J. Cell Biol. 1975, 66, 351–366. [Google Scholar] [CrossRef]
- Hsieh, S.-T.; Crawford, T.O.; Griffin, J.W. Neurofilament distribution and organization in the myelinated axons of the peripheral nervous system. Brain Res. 1994, 642, 316–326. [Google Scholar] [CrossRef]
- Janmey, P.A.; Leterrier, J.-F.; Herrmann, H. Assembly and structure of neurofilaments. Curr. Opin. Colloid Interface Sci. 2003, 8, 40–47. [Google Scholar] [CrossRef]
- Jayanthi, L. Computational Investigation on the Structural Properties of Neurofilaments and Their Sidearms; Wayne State University: Detroit, MI, USA, 2014. [Google Scholar]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Zetterberg, H.; Van Straaten, E.; Lind, K.; Syversen, S.; Edman, Å.; Blennow, K.; Rosengren, L.; Pantoni, L.; Inzitari, D. Cerebrospinal fluid biomarkers of white matter lesions–cross-sectional results from the LADIS study. Eur. J. Neurol. 2010, 17, 377–382. [Google Scholar] [CrossRef]
- Kaji, T.; Boland, B.; Odrljin, T.; Mohan, P.; Basavarajappa, B.S.; Peterhoff, C.; Cataldo, A.; Rudnicki, A.; Amin, N.; Li, B.S. Calpain mediates calcium-induced activation of the erk1, 2 MAPK pathway and cytoskeletal phosphorylation in neurons: Relevance to Alzheimer’s disease. Am. J. Pathol. 2004, 165, 795–805. [Google Scholar]
- Khan, M.I.; Hasan, F.; Al Mahmud, K.A.H.; Adnan, A. Recent computational approaches on mechanical behavior of axonal cytoskeletal components of neuron: A Brief Review. Multiscale Sci. Eng. 2020, 1–15. [Google Scholar] [CrossRef]
- Khan, M.I.; Hasan, F.; Hasan Al Mahmud, K.A.; Adnan, A. Domain focused and residue focused phosphorylation effect on tau protein: A molecular dynamics simulation study. J. Mech. Behav. Biomed. Mater. 2020, 113, 104149. [Google Scholar] [CrossRef]
- Kim, B.; Lee, S.B.; Lee, J.; Cho, S.; Park, H.; Yeom, S.; Park, S.H. A comparison among Neo-Hookean model, Mooney-Rivlin model, and Ogden model for chloroprene rubber. Int. J. Precis. Eng. Manuf. 2012, 13, 759–764. [Google Scholar] [CrossRef]
- Kim, S.; Chang, R.; Teunissen, C.; Gebremichael, Y.; Petzold, A. Neurofilament stoichiometry simulations during neurodegeneration suggest a remarkable self-sufficient and stable in vivo protein structure. J. Neurol. Sci. 2011, 307, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Yin, X.; Trapp, B.D.; Hoh, J.H.; Paulaitis, M.E. Relating interactions between neurofilaments to the structure of axonal neurofilament distributions through polymer brush models. Biophys. J. 2002, 82, 2360–2372. [Google Scholar] [CrossRef] [Green Version]
- Kushkuley, J.; Metkar, S.; Chan, W.K.H.; Lee, S.; Shea, T.B. Aluminum induces neurofilament aggregation by stabilizing cross-bridging of phosphorylated c-terminal sidearms. Brain Res. 2010, 1322, 118–123. [Google Scholar] [CrossRef]
- Laser-Azogui, A.; Kornreich, M.; Malka-Gibor, E.; Beck, R. Neurofilament assembly and function during neuronal development. Curr. Opin. Cell Biol. 2010, 32, 92–101. [Google Scholar] [CrossRef]
- Leterrier, J.-F.; Eyer, J. Properties of highly viscous gels formed by neurofilaments in vitro. A possible consequence of a specific inter-filament cross-bridging. Biochem. J. 1987, 245, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Leterrier, J.F.; Käs, J.; Hartwig, J.; Vegners, R.; Janmey, P.A. Mechanical effects of neurofilament cross-bridges modulation by phosphorylation, lipids, and interactions with f-actin. J. Biol. Chem. 1996, 271, 15687–15694. [Google Scholar] [CrossRef] [Green Version]
- MacKerell, A.D.; Bashford, D.; Bellott, M.; Dunbrack, R.L.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-atom empirical potential for molecular modeling and dynamics studies of proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef]
- Minami, Y.; Sakai, H. Effects of microtubule-associated proteins on network formation by neurofilament-induced polymerization of tubulin. FEBS Lett. 1986, 195, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Nixon, R.A.; Shea, T.B. Dynamics of neuronal intermediate filaments: A developmental perspective. Cell Motil. Cytoskelet. 1992, 22, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Pant, H.C. Dephosphorylation of neurofilament proteins enhances their susceptibility to degradation by calpain. Biochem. J. 1988, 256, 665–668. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, V.; Gal, N.; Fournier, P.; Kilfoil, M.L. Microrheology of microtubule solutions and actin-microtubule composite networks. Phys. Rev. Lett. 2009, 102, 188303. [Google Scholar] [CrossRef] [Green Version]
- Perrot, R.; Eyer, J. Neuronal intermediate filaments and neurodegenerative disorders. Brain Res. Bull. 2009, 80, 282–295. [Google Scholar] [CrossRef]
- Peter, S.J.; Mofrad, M.R.K. Computational modeling of axonal microtubule bundles under tension. Biophys. J. 2012, 102, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plimpton, S. Fast parallel algorithms for short-range molecular dynamics. J. Comput. Phys. 1995, 117, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Rammensee, S.; Janmey, P.A.; Bausch, A.R. Mechanical and structural properties of in vitro neurofilament hydrogels. Eur. Biophys. J. 2007, 36, 661–668. [Google Scholar] [CrossRef]
- Rao, M.V.; Houseweart, M.K.; Williamson, T.L.; Crawford, T.O.; Folmer, J.; Cleveland, D.W. Neurofilament-dependent radial growth of motor axons and axonal organization of neurofilaments does not require the neurofilament heavy subunit (NF-H) or its phosphorylation. J. Cell Biol. 1998, 143, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.V.; Mohan, P.S.; Kumar, A.; Yuan, A.; Montagna, L.; Campbell, J.; Espreafico, E.M.; Julien, J.P.; Nixon, R.A. The myosin Va head domain binds to the neurofilament-L rod and modulates endoplasmic reticulum (ER) content and distribution within axons. PLoS ONE 2011, 6, e17087. [Google Scholar] [CrossRef]
- Rosengren, L.E.; Karlsson, J.E.; Karlsson, J.O.; Persson, L.I.; Wikkelsø, C. Patients with amyotrophic lateral sclerosis and other neurodegenerative diseases have increased levels of neurofilament protein in CSF. J. Neurochem. 1996, 67, 2013–2018. [Google Scholar] [CrossRef]
- Runge, M.S.; Laue, T.M.; Yphantis, D.A.; Lifsics, M.R.; Saito, A.; Altin, M.; Reinke, K.; Williams, R.C. ATP-induced formation of an associated complex between microtubules and neurofilaments. Proc. Natl. Acad. Sci. USA 1981, 78, 1431–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rycroft, C. Voro++: A Three-Dimensional Voronoi Cell Library in C++; Lawrence Berkeley National Lab (LBNL): Berkeley, CA, USA, 2009.
- Stevens, M.J.; Hoh, J.H. Conformational dynamics of neurofilament side-arms. J. Phys. Chem. B 2010, 114, 8879–8886. [Google Scholar] [CrossRef] [PubMed]
- Stukowski, A. Visualization and analysis of atomistic simulation data with OVITO–the open visualization tool. Model. Simul. Mater. Sci. Eng. 2010, 18, 015012. [Google Scholar] [CrossRef]
- Szaro, B.G.; Strong, M.J. Post-transcriptional control of neurofilaments: New roles in development, regeneration and neurodegenerative disease. Trends Neurosci. 2010, 33, 27–37. [Google Scholar] [CrossRef]
- Van den Bedem, H.; Kuhl, E. Tau-ism: The Yin and Yang of microtubule sliding, detachment, and rupture. Biophys. J. 2015, 109, 2215. [Google Scholar] [CrossRef] [Green Version]
- Wagner, O.I.; Rammensee, S.; Korde, N.; Wen, Q.; Leterrier, J.-F.; Janmey, P.A. Softness, strength and self-repair in intermediate filament networks. Exp. Cell Res. 2007, 313, 2228–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.Y.; Ru, C.Q.; Mioduchowski, A. Orthotropic elastic shell model for buckling of microtubules. Phys. Rev. E 2006, 74, 52901. [Google Scholar] [CrossRef]
- Wang, H.; Wu, M.; Zhan, C.; Ma, E.; Yang, M.; Yang, X.; Li, Y. Neurofilament proteins in axonal regeneration and neurodegenerative diseases. Neural Regen. Res. 2012, 7, 620. [Google Scholar]
- Wen, G.Y.; Wisniewski, H.M. Substructures of neurofilaments. Acta Neuropathol. 1984, 64, 339–343. [Google Scholar] [CrossRef]
- Wu, Y.-T.; Adnan, A. Damage and failure of axonal microtubule under extreme high strain rate: An in-silico molecular dynamics study. Sci. Rep. 2018, 8, 12260. [Google Scholar] [CrossRef]
- Yao, N.Y.; Broedersz, C.P.; Lin, Y.-C.; Kasza, K.E.; MacKintosh, F.C.; Weitz, D.A. Elasticity in ionically cross-linked neurofilament networks. Biophys. J. 2010, 98, 2147–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, A.; Rao, M.V.; Nixon, R.A. Neurofilaments at a glance. J. Cell Sci. 2012, 125, 3257–3263. [Google Scholar] [CrossRef] [PubMed]
- Yuan, A.; Rao, M.V.; Sasaki, T.; Chen, Y.; Kumar, A.; Liem, R.K.H.; Eyer, J.; Peterson, A.C.; Julien, J.-P.; Nixon, R.A. α-internexin is structurally and functionally associated with the neurofilament triplet proteins in the mature CNS. J. Neurosci. 2006, 26, 10006–10019. [Google Scholar] [CrossRef]
- Yuan, A.; Sasaki, T.; Kumar, A.; Peterhoff, C.M.; Rao, M.V.; Liem, R.K.; Julien, J.-P.; Nixon, R.A. Peripherin is a subunit of peripheral nerve neurofilaments: Implications for differential vulnerability of CNS and peripheral nervous system axons. J. Neurosci. 2012, 32, 8501–8508. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Raina, A.K.; Rottkamp, C.A.; Aliev, G.; Perry, G.; Boux, H.; Smith, M.A. Activation and redistribution of c-jun N-terminal kinase/stress activated protein kinase in degenerating neurons in Alzheimer’s disease. J. Neurochem. 2001, 76, 435–441. [Google Scholar] [CrossRef]
- Zhulina, E.B.; Leermakers, F.A.M. The polymer brush model of neurofilament projections: Effect of protein composition. Biophys. J. 2010, 98, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Zhulina, E.B.; Leermakers, F.A.M. Effect of the ionic strength and pH on the equilibrium structure of a neurofilament brush. Biophys. J. 2007, 93, 1452–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhulina, E.B.; Leermakers, F.A.M. A self-consistent field analysis of the neurofilament brush with amino-acid resolution. Biophys. J. 2007, 93, 1421–1430. [Google Scholar] [CrossRef] [Green Version]








| Strain Rate, 1/s | 1 × 108 | 1 × 109 |
|---|---|---|
| E1, GPa | 0.103 | 0 |
| E2, GPa | 0.827 | 2 |
| τ, ps | 87.866 | 100 |
| Poisson’s Ratio, ν | 0.33 | 0.48 | ||
|---|---|---|---|---|
| Strain Rate, 1/s | 1 × 108 | 1 × 109 | 1 × 108 | 1 × 109 |
| G1, GPa | 0.039 | 0 | 0.035 | 0 |
| G2, GPa | 0.314 | 0.76 | 0.281 | 0.68 |
| τ (for G), ps | 90.909 | 100 | 90.909 | 100 |
| K1, GPa | 0.1 | 0 | 0.858 | 0 |
| K2, GPa | 0.81 | 1.96 | 6.889 | 16.66 |
| τ (for K), ps | 90.909 | 100 | 90.909 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.I.; Hasan, F.; Mahmud, K.A.H.A.; Adnan, A. Viscoelastic Response of Neurofilaments: An Atomistic Simulation Approach. Biomolecules 2021, 11, 540. https://doi.org/10.3390/biom11040540
Khan MI, Hasan F, Mahmud KAHA, Adnan A. Viscoelastic Response of Neurofilaments: An Atomistic Simulation Approach. Biomolecules. 2021; 11(4):540. https://doi.org/10.3390/biom11040540
Chicago/Turabian StyleKhan, Md Ishak, Fuad Hasan, Khandakar Abu Hasan Al Mahmud, and Ashfaq Adnan. 2021. "Viscoelastic Response of Neurofilaments: An Atomistic Simulation Approach" Biomolecules 11, no. 4: 540. https://doi.org/10.3390/biom11040540
APA StyleKhan, M. I., Hasan, F., Mahmud, K. A. H. A., & Adnan, A. (2021). Viscoelastic Response of Neurofilaments: An Atomistic Simulation Approach. Biomolecules, 11(4), 540. https://doi.org/10.3390/biom11040540