
Protein–Protein Connections—Oligomer, Amyloid and Protein Complex—By Wide Line 1H NMR


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
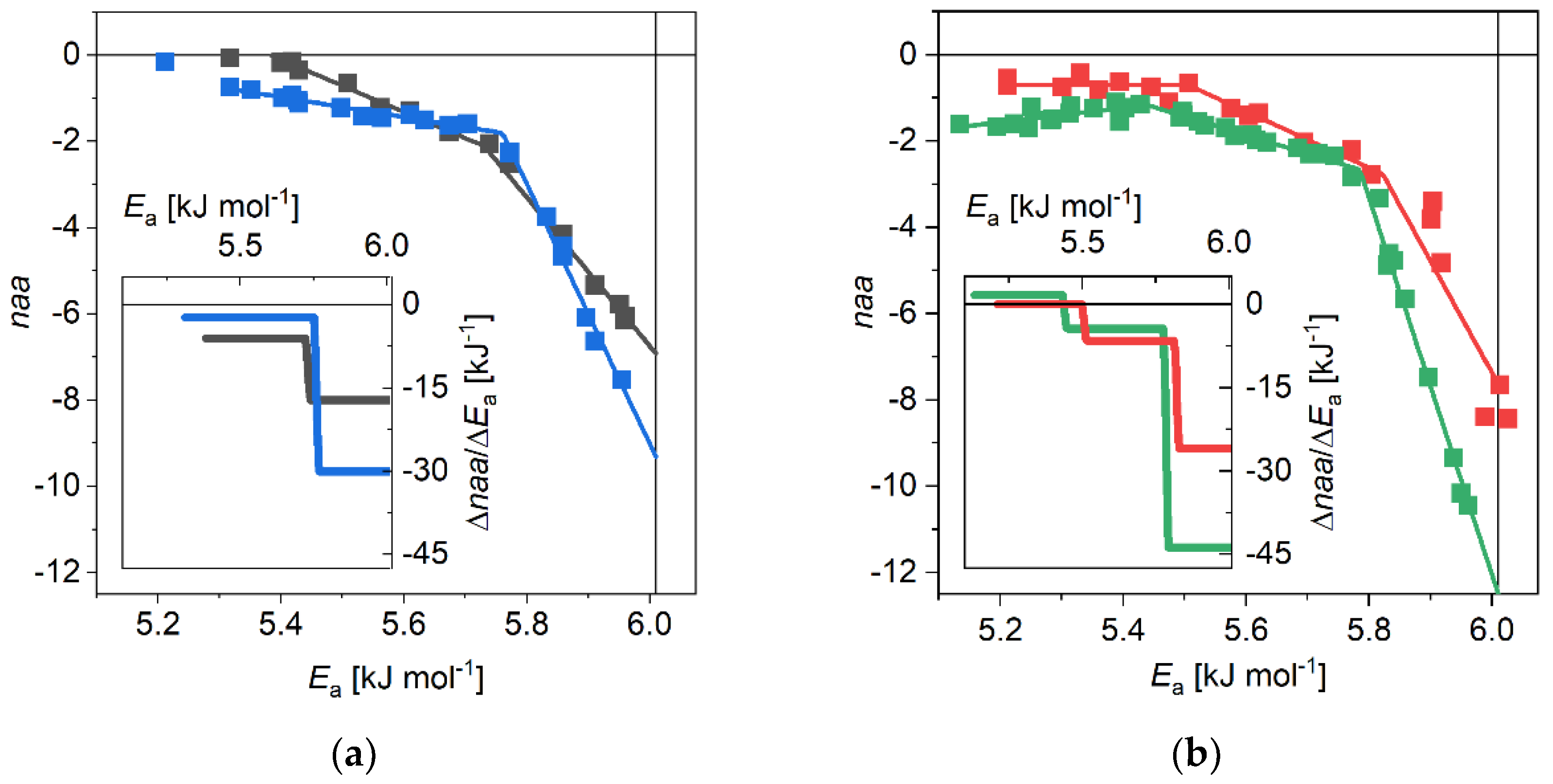
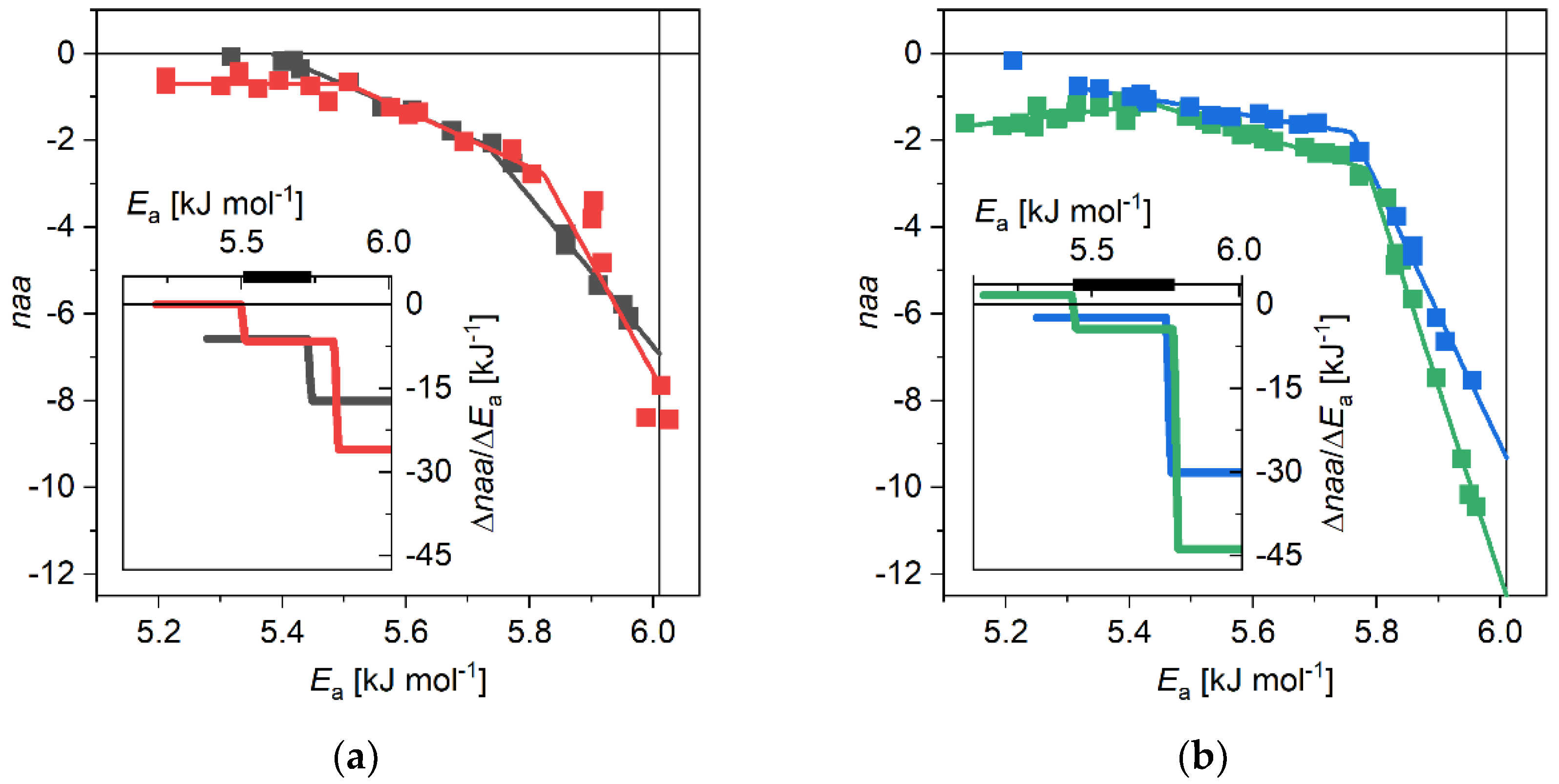
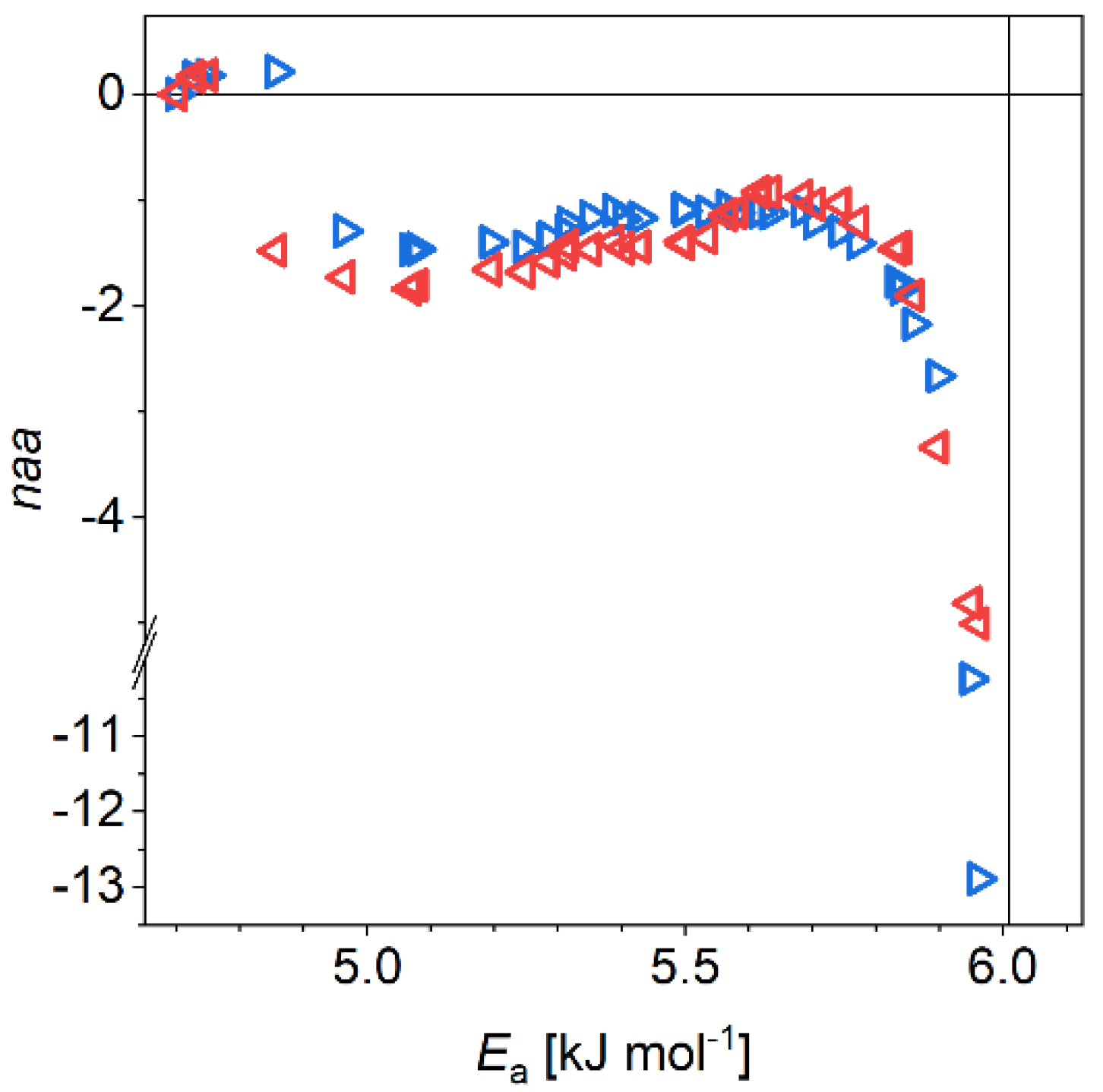
3.1. The α-Synuclein System
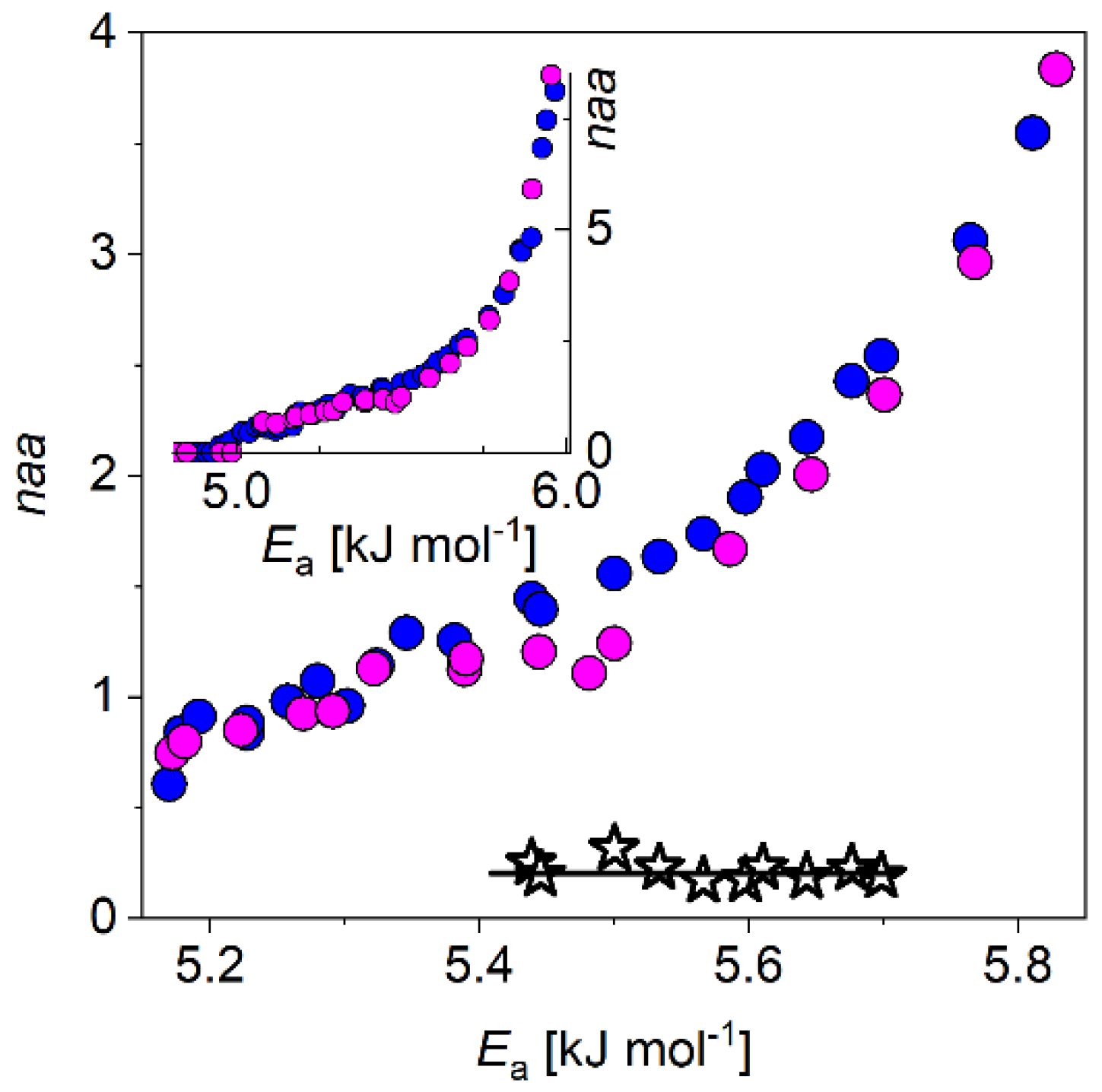
3.2. The Stabilin CTD-Tβ4 System
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A
Wide-Line 1H NMR-Spectrometry
References
- Tompa, P.; Schad, E.; Tantos, A.; Kalmar, L. Intrinsically disordered proteins: Emerging interaction specialists. Curr. Opin. Struct. Biol. 2015, 35, 49–59. [Google Scholar] [CrossRef]
- Hazy, E.; Bokor, M.; Kalmar, L.; Gelencser, A.; Kamasa, P.; Han, K.-H.; Tompa, K.; Tompa, P. Distinct Hydration Properties of Wild-Type and Familial Point Mutant A53T of α-Synuclein Associated with Parkinson’s Disease. Biophys. J. 2011, 101, 2260–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokor, M.; Tantos, Á.; Tompa, P.; Han, K.-H.; Tompa, K. WT and A53T α-Synuclein Systems: Melting Diagram and Its New Interpretation. Int. J. Mol. Sci. 2020, 21, 3997. [Google Scholar] [CrossRef] [PubMed]
- Tantos, A.; Szabó, B.; Láng, A.; Varga, Z.; Tsylonok, M.; Bokor, M.; Verebelyi, T.; Kamasa, P.; Tompa, K.; Perczel, A.; et al. Multiple fuzzy interactions in the moonlighting function of thymosin-β4. Intrinsically Disord. Proteins 2013, 1, e26204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokor, M.; Tantos, Á.; Mészáros, A.; Jenei, B.; Haminda, R.; Tompa, P.; Tompa, K. Molecular Motions and Interactions in Aqueous Solutions of Thymosin-β 4, Stabilin CTD and Their 1: 1 Complex, Studied by 1 H-NMR Spectroscopy. ChemPhysChem 2020, 21, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M. Alpha-synuclein and neurodegenerative diseases. Nat. Rev. Neurosci. 2001, 2, 492–501. [Google Scholar] [CrossRef]
- Scott, D.; Roy, S. -Synuclein Inhibits Intersynaptic Vesicle Mobility and Maintains Recycling-Pool Homeostasis. J. Neurosci. 2012, 32, 10129–10135. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Wang, L.; Bao, H.; Premi, S.; Das, U.; Chapman, E.R.; Roy, S. Functional cooperation of α-synuclein and VAMP2 in synaptic vesicle recycling. Proc. Natl. Acad. Sci. USA 2019, 116, 11113–11115. [Google Scholar] [CrossRef] [Green Version]
- Jo, E.; Fuller, N.; Rand, R.; George-Hyslop, P.S.; Fraser, P.E. Defective membrane interactions of familial Parkinson’s disease mutant A30P α-synuclein. J. Mol. Biol. 2002, 315, 799–807. [Google Scholar] [CrossRef]
- Spira, P.J.; Sharpe, D.M.; Halliday, G.; Cavanagh, J.; Nicholson, G.A. Clinical and pathological features of a parkinsonian syndrome in a family with an Ala53Thr ?-synuclein mutation. Ann. Neurol. 2001, 49, 313–319. [Google Scholar] [CrossRef]
- Marchalonis, J. Lymphocyte surface immunoglobulins. Science 1975, 190, 20–29. [Google Scholar] [CrossRef]
- Burré, J.; Sharma, M.; Südhof, T.C. Definition of a Molecular Pathway Mediating -Synuclein Neurotoxicity. J. Neurosci. 2015, 35, 5221–5232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outeiro, T.F.; Putcha, P.; Tetzlaff, J.E.; Spoelgen, R.; Koker, M.; Carvalho, F.; Hyman, B.T.; McLean, P.J. Formation of Toxic Oligomeric α-Synuclein Species in Living Cells. PLoS ONE 2008, 3, e1867. [Google Scholar] [CrossRef]
- Winner, B.; Jappelli, R.; Maji, S.K.; Desplats, P.A.; Boyer, L.; Aigner, S.; Hetzer, C.; Loher, T.; Vilar, M.; Campioni, S.; et al. In vivo demonstration that -synuclein oligomers are toxic. Proc. Natl. Acad. Sci. USA 2011, 108, 4194–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremades, N.; Cohen, S.I.; Deas, E.; Abramov, A.Y.; Chen, A.Y.; Orte, A.; Sandal, M.; Clarke, R.W.; Dunne, P.; Aprile, F.A.; et al. Direct Observation of the Interconversion of Normal and Toxic Forms of α-Synuclein. Cell 2012, 149, 1048–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prots, I.; Veber, V.; Brey, S.; Campioni, S.; Buder, K.; Riek, R.; Böhm, K.J.; Winner, B. α-Synuclein Oligomers Impair Neuronal Microtubule-Kinesin Interplay. J. Biol. Chem. 2013, 288, 21742–21754. [Google Scholar] [CrossRef] [Green Version]
- Dunker, A.K.; Silman, I.; Uversky, V.N.; Sussman, J.L. Function and structure of inherently disordered proteins. Curr. Opin. Struct. Biol. 2008, 18, 756–764. [Google Scholar] [CrossRef]
- Mor, D.E.; Ugras, S.E.; Daniels, M.J.; Ischiropoulos, H. Dynamic structural flexibility of α-synuclein. Neurobiol. Dis. 2016, 88, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Minde, D.-P.; Dunker, A.K.; Lilley, K.S. Time, space, and disorder in the expanding proteome universe. Proteome 2017, 17, 1600399. [Google Scholar] [CrossRef]
- Chen, J.; Kriwacki, R.W. Intrinsically Disordered Proteins: Structure, Function and Therapeutics. J. Mol. Biol. 2018, 430, 2275–2277. [Google Scholar] [CrossRef]
- Li, J.; Uversky, V.N.; Fink, A.L. Conformational Behavior of Human α-Synuclein is Modulated by Familial Parkinson’s Disease Point Mutations A30P and A53T. NeuroToxicology 2002, 23, 553–567. [Google Scholar] [CrossRef]
- Conway, K.A.; Harper, J.D.; Lansbury, P.T. Accelerated in vitro fibril formation by a mutant α-synuclein linked to early-onset Parkinson disease. Nat. Med. 1998, 4, 1318–1320. [Google Scholar] [CrossRef] [PubMed]
- Heise, H.; Celej, M.S.; Becker, S.; Riedel, D.; Pelah, A.; Kumar, A.; Jovin, T.M.; Baldus, M. Solid-State NMR Reveals Structural Differences between Fibrils of Wild-Type and Disease-Related A53T Mutant α-Synuclein. J. Mol. Biol. 2008, 380, 444–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brucale, M.; Sandal, M.; Di Maio, S.; Rampioni, A.; Tessari, I.; Tosatto, L.; Bisaglia, M.; Bubacco, L.; Samorì, B. Pathogenic Mutations Shift the Equilibria of α-Synuclein Single Molecules towards Structured Conformers. ChemBioChem 2009, 10, 176–183. [Google Scholar] [CrossRef]
- Giasson, B.I.; Uryu, K.; Trojanowski, J.Q.; Lee, V.M.-Y. Mutant and Wild Type Human α-Synucleins Assemble into Elongated Filaments with Distinct Morphologies in Vitro. J. Biol. Chem. 1999, 274, 7619–7622. [Google Scholar] [CrossRef] [Green Version]
- Bertoncini, C.W.; Jung, Y.-S.; Fernandez, C.O.; Hoyer, W.; Griesinger, C.; Jovin, T.M.; Zweckstetter, M. From the Cover: Release of long-range tertiary interactions potentiates aggregation of natively unstructured -synuclein. Proc. Natl. Acad. Sci. USA 2005, 102, 1430–1435. [Google Scholar] [CrossRef] [Green Version]
- Narhi, L.; Wood, S.J.; Steavenson, S.; Jiang, Y.; Wu, G.M.; Anafi, D.; Kaufman, S.A.; Martin, F.; Sitney, K.; Denis, P.; et al. Both Familial Parkinson’s Disease Mutations Accelerate α-Synuclein Aggregation. J. Biol. Chem. 1999, 274, 9843–9846. [Google Scholar] [CrossRef] [Green Version]
- Coskuner, O.; Wise-Scira, O. Structures and Free Energy Landscapes of the A53T Mutant-Type α-Synuclein Protein and Impact of A53T Mutation on the Structures of the Wild-Type α-Synuclein Protein with Dynamics. ACS Chem. Neurosci. 2013, 4, 1101–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serpell, L.C.; Berriman, J.; Jakes, R.; Goedert, M.; Crowther, R.A. Fiber diffraction of synthetic alpha -synuclein filaments shows amyloid-like cross-beta conformation. Proc. Natl. Acad. Sci. USA 2000, 97, 4897–4902. [Google Scholar] [CrossRef] [Green Version]
- Kamiyoshihara, T.; Kojima, M.; Ueda, K.; Tashiro, M.; Shimotakahara, S. Observation of multiple intermediates in α-synuclein fibril formation by singular value decomposition analysis. Biochem. Biophys. Res. Commun. 2007, 355, 398–403. [Google Scholar] [CrossRef]
- Lashuel, H.A.; Petre, B.M.; Wall, J.; Simon, M.; Nowak, R.J.; Walz, T.; Lansbury, P.T. α-Synuclein, Especially the Parkinson’s Disease-associated Mutants, Forms Pore-like Annular and Tubular Protofibrils. J. Mol. Biol. 2002, 322, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Conway, K.A.; Lee, S.-J.; Rochet, J.-C.; Ding, T.T.; Williamson, R.E.; Lansbury, P.T. Acceleration of oligomerization, not fibrillization, is a shared property of both alpha -synuclein mutations linked to early-onset Parkinson’s disease: Implications for pathogenesis and therapy. Proc. Natl. Acad. Sci. USA 2000, 97, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Ikeda, T.; Takasaki, J.-I.; Yamada, M. Familial Parkinson disease mutations influence α-synuclein assembly. Neurobiol. Dis. 2011, 43, 715–724. [Google Scholar] [CrossRef]
- Gallardo, R.; Ranson, N.A.; Radford, S.E. Amyloid structures: Much more than just a cross-β fold. Curr. Opin. Struct. Biol. 2020, 60, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, M. Fibril formation from the amyloid-β peptide is governed by a dynamic equilibrium involving association and dissociation of the monomer. Biophys. Rev. 2017, 9, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Tipping, K.W.; Karamanos, T.K.; Jakhria, T.; Iadanza, M.G.; Goodchild, S.C.; Tuma, R.; Ranson, N.A.; Hewitt, E.W.; Radford, S.E. pH-induced molecular shedding drives the formation of amyloid fibril-derived oligomers. Proc. Natl. Acad. Sci. USA 2015, 112, 5691–5696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safer, D.; Elzinga, M.; Nachmias, V.T. Thymosin β4 and Fx, an actin-sequestering peptide, are indistinguishable. J. Biol. Chem. 1991, 266, 4029–4032. [Google Scholar] [CrossRef]
- Yarmola, E.G.; Klimenko, E.S.; Fujita, G.; Bubb, M.R. Thymosin beta4: Actin Regulation and More. Ann. N. Y. Acad. Sci. 2007, 1112, 76–85. [Google Scholar] [CrossRef]
- Grant, D.S.; Rose, W.; Yaen, C.; Goldstein, A.; Martinez, J.; Kleinman, H. Thymosin β4 enhances endothelial cell differentiation and angiogenesis. Angiogenesis 1999, 3, 125–135. [Google Scholar] [CrossRef]
- Smart, N.; Rossdeutsch, A.; Riley, P.R. Thymosin β4 and angiogenesis: Modes of action and therapeutic potential. Angiogenesis 2007, 10, 229–241. [Google Scholar] [CrossRef]
- Qiu, P.; Sosne, G.; Kurpakus-Wheater, M. Matrix metalloproteinase activity is necessary for thymosin beta 4 promotion of epithelial cell migration. J. Cell. Physiol. 2007, 212, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Domanski, M.; Hertzog, M.; Coutant, J.; Gutsche-Perelroizen, I.; Bontems, F.; Carlier, M.-F.; Guittet, E.; van Heijenoort, C. Coupling of Folding and Binding of Thymosin β4 upon Interaction with Monomeric Actin Monitored by Nuclear Magnetic Resonance. J. Biol. Chem. 2004, 279, 23637–23645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertzog, M.; van Heijenoort, C.; Didry, D.; Gaudier, M.; Coutant, J.; Gigant, B.; Didelot, G.; Préat, T.; Knossow, M.; Guittet, E.; et al. The β-Thymosin/WH2 Domain: Structural Basis for the Switch from Inhibition to Promotion of Actin Assembly. Cell 2004, 117, 611–623. [Google Scholar] [CrossRef]
- Ho, J.H.-C.; Tseng, K.-C.; Ma, W.-H.; Chen, K.-H.; Lee, O.K.-S.; Su, Y. Thymosin beta-4 upregulates anti-oxidative enzymes and protects human cornea epithelial cells against oxidative damage. Br. J. Ophthalmol. 2008, 92, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; So, I.-S.; Park, S.-Y.; Kim, I.-S. Thymosin β4 is involved in stabilin-2-mediated apoptotic cell engulfment. FEBS Lett. 2008, 582, 2161–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podoly, E.; Hanin, G.; Soreq, H. Alanine-to-threonine substitutions and amyloid diseases: Butyrylcholinesterase as a case study. Chem. Interac. 2010, 187, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, D. Extreme Fuzziness: Direct Interactions between Two IDPs. Biomolecules 2019, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CRC. Handbook of Chemistry and Physics, 101st ed.; CRC Press: Boca Raton, FL, USA, 2020; ISBN 9780367417246. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bokor, M.; Tantos, Á. Protein–Protein Connections—Oligomer, Amyloid and Protein Complex—By Wide Line 1H NMR. Biomolecules 2021, 11, 757. https://doi.org/10.3390/biom11050757
Bokor M, Tantos Á. Protein–Protein Connections—Oligomer, Amyloid and Protein Complex—By Wide Line 1H NMR. Biomolecules. 2021; 11(5):757. https://doi.org/10.3390/biom11050757
Chicago/Turabian StyleBokor, Mónika, and Ágnes Tantos. 2021. "Protein–Protein Connections—Oligomer, Amyloid and Protein Complex—By Wide Line 1H NMR" Biomolecules 11, no. 5: 757. https://doi.org/10.3390/biom11050757
APA StyleBokor, M., & Tantos, Á. (2021). Protein–Protein Connections—Oligomer, Amyloid and Protein Complex—By Wide Line 1H NMR. Biomolecules, 11(5), 757. https://doi.org/10.3390/biom11050757