
Engineering the Vasculature of Stem-Cell-Derived Liver Organoids


Abstract
:1. Introduction
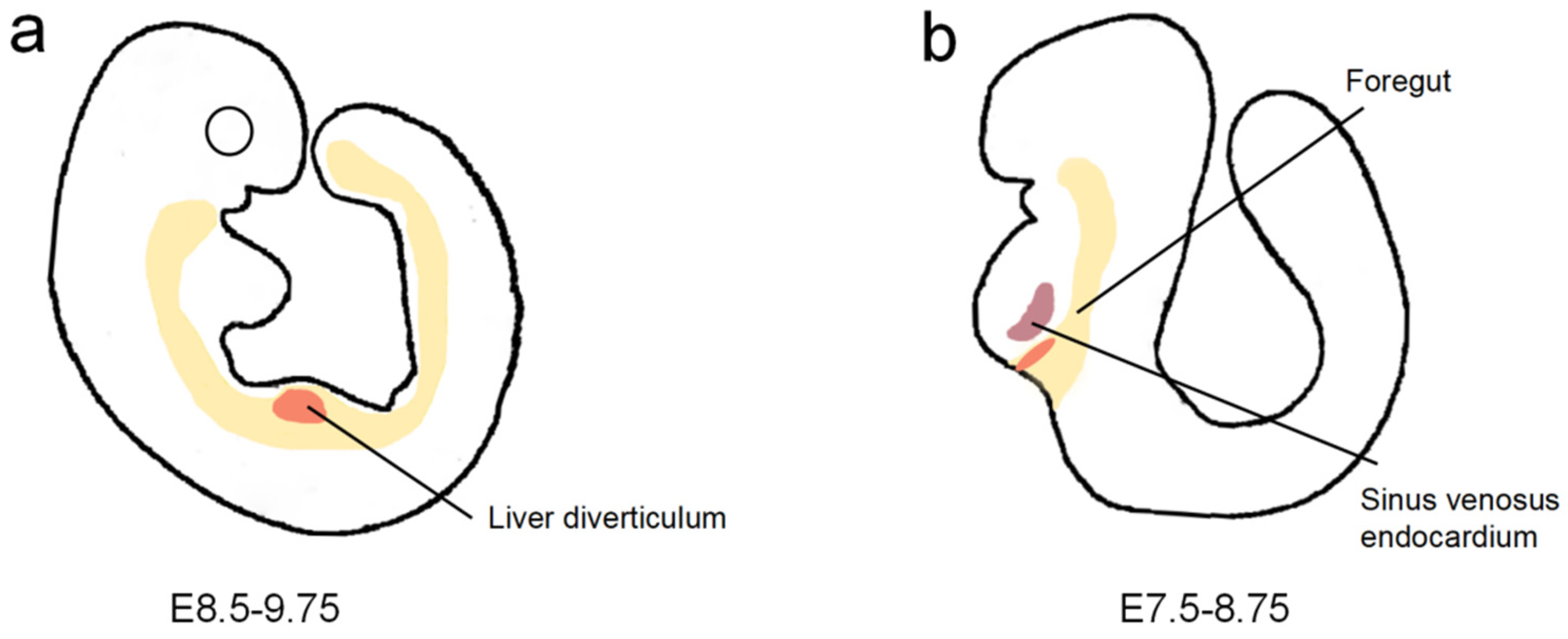
2. Liver Vascularization in Embryonic Development
2.1. Origin of Hepatic Endothelial Cells
2.2. Morphogenesis of Veins, Arteries, and Sinusoids
3. Vascularization and Maturation of Liver Organoids
3.1. Self-Organization in Well Arrays
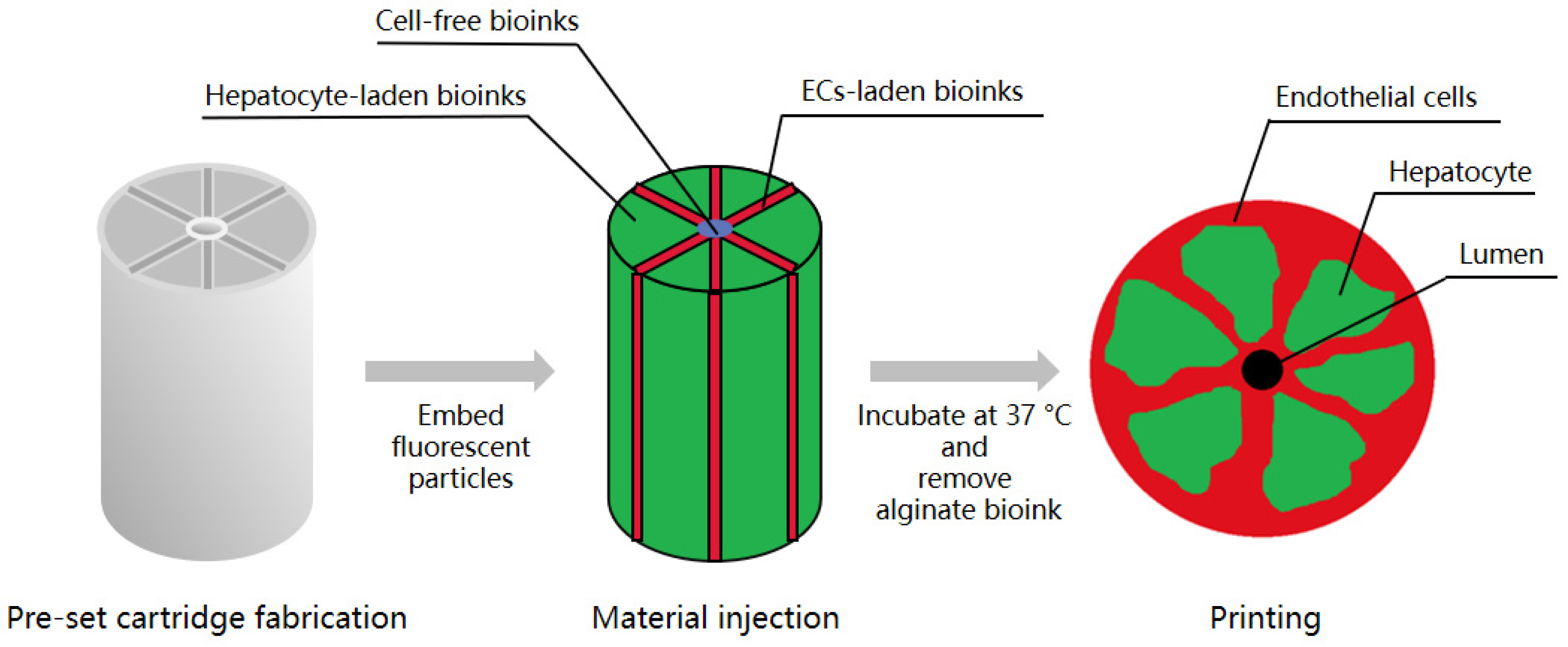
3.2. Three-Dimensional Bioprinting
3.3. Decellularization
3.4. In Vivo Transplantation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fisher, R.A. Living donor liver transplantation: Eliminating the wait for death in end-stage liver disease? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 373–382. [Google Scholar] [CrossRef]
- Bernal, W.; Hyyrylainen, A.; Gera, A.; Audimoolam, V.K.; McPhail, M.J.; Auzinger, G.; Rela, M.; Heaton, N.; O’Grady, J.G.; Wendon, J.; et al. Lessons from look-back in acute liver failure? A single centre experience of 3300 patients. J. Hepatol. 2013, 59, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Kuse, Y.; Taniguchi, H. Present and Future Perspectives of Using Human-Induced Pluripotent Stem Cells and Organoid Against Liver Failure. Cell Transplant. 2019, 28, 160S–165S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef]
- Hu, H.; Gehart, H.; Artegiani, B.; Löpez-Iglesias, C.; Dekkers, F.; Basak, O.; van Es, J.; de Sousa Lopes, S.M.C.; Begthel, H.; Korving, J.; et al. Long-Term Expansion of Functional Mouse and Human Hepatocytes as 3D Organoids. Cell 2018, 175, 1591–1606.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huch, M.; Dorrell, C.; Boj, S.F.; Van Es, J.H.; Li, V.S.; Van De Wetering, M.; Sato, T.; Hamer, K.; Sasaki, N.; Finegold, M.J.; et al. In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature 2013, 494, 247–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mun, S.J.; Ryu, J.-S.; Lee, M.-O.; Son, Y.S.; Oh, S.J.; Cho, H.-S.; Son, M.-Y.; Kim, D.-S.; Kim, S.J.; Yoo, H.J.; et al. Generation of expandable human pluripotent stem cell-derived hepatocyte-like liver organoids. J. Hepatol. 2019, 71, 970–985. [Google Scholar] [CrossRef]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N.; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Grebenyuk, S.; Ranga, A. Engineering Organoid Vascularization. Front. Bioeng. Biotechnol. 2019, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSesso, J.M. Vascular ontogeny within selected thoracoabdominal organs and the limbs. Reprod. Toxicol. 2017, 70, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Cast, A.E.; Walter, T.J.; Huppert, S.S. Vascular patterning sets the stage for macro and micro hepatic architecture. Dev. Dyn. 2015, 244, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Si-Tayeb, K.; Lemaigre, F.; Duncan, S.A. Organogenesis and Development of the Liver. Dev. Cell 2010, 18, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, Y.; Takabe, Y.; Koike, T.; Nakakura, T.; Tanaka, S.; Shiojiri, N. Sinusoid development and morphogenesis may be stimulated by VEGF-Flk-1 signaling during fetal mouse liver development. Dev. Dyn. 2009, 239, 386–397. [Google Scholar] [CrossRef]
- Zhang, H.; Pu, W.; Tian, X.; Huang, X.; He, L.; Liu, Q.; Lingjuan, H.; Zhang, L.; He, L.; Liu, K.; et al. Genetic lineage tracing identifies endocardial origin of liver vasculature. Nat. Genet. 2016, 48, 537–543. [Google Scholar] [CrossRef]
- Goldman, O.; Han, S.; Hamou, W.; de Villeroche, V.J.; Uzan, G.; Lickert, H.; Gouon-Evans, V. Endoderm Generates Endothelial Cells during Liver Development. Stem Cell Rep. 2014, 3, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Boström, K.I.; Yao, J.; Wu, X.; Yao, Y. Endothelial Cells May Have Tissue-Specific Origins. J. Cell Biol. Histol. 2018, 1, 1. [Google Scholar]
- Ober, E.A.; Lemaigre, F.P. Development of the liver: Insights into organ and tissue morphogenesis. J. Hepatol. 2018, 68, 1049–1062. [Google Scholar] [CrossRef] [Green Version]
- Fabris, L.; Cadamuro, M.; Libbrecht, L.; Raynaud, P.; Spirli, C.; Fiorotto, R.; Okolicsanyi, L.; Lemaigre, F.; Strazzabosco, M.; Roskams, T. Epithelial expression of angiogenic growth factors modulate arterial vasculogenesis in human liver development. Hepatology 2007, 47, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Koch, P.-S.; Lee, K.H.; Goerdt, S.; Augustin, H.G. Angiodiversity and organotypic functions of sinusoidal endothelial cells. Angiogenesis 2021, 24, 289–310. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.; Demory, A.; Peyre, F.; Kroll, J.; Augustin, H.G.; Helfrich, W.; Kzhyshkowska, J.; Schledzewski, K.; Arnold, B.; Goerdt, S. Wnt2 acts as a cell type-specific, autocrine growth factor in rat hepatic sinusoidal endothelial cells cross-stimulating the VEGF pathway. Hepatology 2008, 47, 1018–1031. [Google Scholar] [CrossRef]
- LeCouter, J.; Moritz, D.R.; Li, B.; Phillips, G.L.; Liang, X.H.; Gerber, H.-P.; Hillan, K.J.; Ferrara, N. Angiogenesis-Independent Endothelial Protection of Liver: Role of VEGFR-1. Science 2003, 299, 890–893. [Google Scholar] [CrossRef]
- Géraud, C.; Koch, P.-S.; Zierow, J.; Klapproth, K.; Busch, K.; Olsavszky, V.; Leibing, T.; Demory, A.; Ulbrich, F.; Diett, M.; et al. GATA4-dependent organ-specific endothelial differentiation controls liver development and embryonic hematopoiesis. J. Clin. Investig. 2017, 127, 1099–1114. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Kimura, M.; Yoshizawa, E.; Ayano, S.; Koido, M.; Funayama, S.; Nakanishi, N.; Hisai, T.; Kobayashi, T.; et al. Massive and Reproducible Production of Liver Buds Entirely from Human Pluripotent Stem Cells. Cell Rep. 2017, 21, 2661–2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettinato, G.; Lehoux, S.; Ramanathan, R.; Salem, M.M.; He, L.-X.; Muse, O.; Flaumenhaft, R.; Thompson, M.T.; Rouse, E.A.; Cummings, R.D.; et al. Generation of fully functional hepatocyte-like organoids from human induced pluripotent stem cells mixed with Endothelial Cells. Sci. Rep. 2019, 9, 8920. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xing, F.; Chen, F.; He, L.; So, K.-F.; Liu, Y.; Xiao, J. Functional 3D Human Liver Bud Assembled from MSC-Derived Multiple Liver Cell Lineages. Cell Transplant. 2019, 28, 510–521. [Google Scholar] [CrossRef]
- Yap, K.K.; Gerrand, Y.-W.; Dingle, A.M.; Yeoh, G.C.; Morrison, W.A.; Mitchell, G.M. Liver sinusoidal endothelial cells promote the differentiation and survival of mouse vascularised hepatobiliary organoids. Biomaterials 2020, 251, 120091. [Google Scholar] [CrossRef] [PubMed]
- Guye, P.; Ebrahimkhani, M.R.; Kipniss, N.; Velazquez, J.J.; Schoenfeld, E.; Kiani, S.; Griffith, L.G.; Weiss, R. Genetically engineering self-organization of human pluripotent stem cells into a liver bud-like tissue using Gata6. Nat. Commun. 2016, 7, 10243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velazquez, J.J.; LeGraw, R.; Moghadam, F.; Tan, Y.; Kilbourne, J.; Maggiore, J.C.; Hislop, J.; Liu, S.; Cats, D.; de Sousa Lopes, S.M.C.; et al. Gene Regulatory Network Analysis and Engineering Directs Development and Vascularization of Multilineage Human Liver Organoids. Cell Syst. 2021, 12, 41–55.e11. [Google Scholar] [CrossRef]
- Cui, H.; Nowicki, M.; Fisher, J.P.; Zhang, L.G. 3D Bioprinting for Organ Regeneration. Adv. Healthcare Mater. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.; Hong, G.; An, S.; Jang, I.; Yun, W.S.; Shim, J.H.; Jin, S. Bioprinting of Multiscaled Hepatic Lobules within a Highly Vascularized Construct. Small 2020, 16, e1905505. [Google Scholar] [CrossRef]
- Yanagi, Y.; Nakayama, K.; Taguchi, T.; Enosawa, S.; Tamura, T.; Yoshimaru, K.; Matsuura, T.; Hayashida, M.; Kohashi, K.; Oda, Y.; et al. In vivo and ex vivo methods of growing a liver bud through tissue connection. Sci. Rep. 2017, 7, 1408. [Google Scholar] [CrossRef]
- Yang, H.; Sun, L.; Pang, Y.; Hu, D.; Xu, H.; Mao, S.; Peng, W.; Wang, Y.; Xu, Y.; Zheng, Y.-C.; et al. Three-dimensional bioprinted hepatorganoids prolong survival of mice with liver failure. Gut 2021, 70, 567–574. [Google Scholar] [CrossRef]
- Derakhshanfar, S.; Mbeleck, R.; Xu, K.; Zhang, X.; Zhong, W.; Xing, M. 3D bioprinting for biomedical devices and tissue engineering: A review of recent trends and advances. Bioact. Mater. 2018, 3, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, Y.; Doi, Y.; Watanabe, H.; Tokairin, T.; Omori, Y.; Su, M.; Yoshioka, T.; Enomoto, K. Transdifferentiation of Mature Rat Hepatocytes into Bile Duct-Like Cells in Vitro. Am. J. Pathol. 2005, 166, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.-X.; Sui, B.-D.; Hu, C.-H.; Qiu, X.-Y.; Zhao, P.; Jin, Y. Reconstruction of structure and function in tissue engineering of solid organs: Toward simulation of natural development based on decellularization. J. Tissue Eng. Regen. Med. 2018, 12, 1432–1447. [Google Scholar] [CrossRef]
- Baptista, P.M.; Siddiqui, M.M.; Lozier, G.; Rodriguez, S.R.; Atala, A.; Soker, S. The use of whole organ decellularization for the generation of a vascularized liver organoid. Hepatology 2011, 53, 604–617. [Google Scholar] [CrossRef]
- Orlando, G.; Farney, A.C.; Iskandar, S.S.; Mirmalek-Sani, S.-H.; Sullivan, D.C.; Moran, E.; AbouShwareb, T.; De Coppi, P.; Wood, K.J.; Stratta, R.J.; et al. Production and Implantation of Renal Extracellular Matrix Scaffolds from Porcine Kidneys as a Platform for Renal Bioengineering Investigations. Ann. Surg. 2012, 256, 363–370. [Google Scholar] [CrossRef]
- Ko, I.K.; Peng, L.; Peloso, A.; Smith, C.J.; Dhal, A.; Deegan, D.B.; Zimmerman, C.; Clouse, C.; Zhao, W.; Shupe, T.D.; et al. Bioengineered transplantable porcine livers with re-endothelialized vasculature. Biomaterials 2015, 40, 72–79. [Google Scholar] [CrossRef]
- Hussein, K.H.; Park, K.-M.; Kang, K.-S.; Woo, H.-M. Heparin-gelatin mixture improves vascular reconstruction efficiency and hepatic function in bioengineered livers. Acta Biomater. 2016, 38, 82–93. [Google Scholar] [CrossRef]
- Takebe, T.; Zhang, R.-R.; Koike, H.; Kimura, M.; Yoshizawa, E.; Enomura, M.; Koike, N.; Sekine, K.; Taniguchi, H. Generation of a vascularized and functional human liver from an iPSC-derived organ bud transplant. Nat. Protoc. 2014, 9, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Mavila, N.; Trecartin, A.; Spurrier, R.; Xiao, Y.; Hou, X.; James, D.; Fu, X.; Truong, B.; Wang, C.; Lipshutz, G.S.; et al. Functional Human and Murine Tissue-Engineered Liver Is Generated from Adult Stem/Progenitor Cells. Stem Cells Transl. Med. 2017, 6, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Method | Material | Vascularization Strategy | References |
|---|---|---|---|
| Self-organization in well arrays | iPSC-HE cells, HUVECs, and BMSCs | Coculture of HUVEC, MSCs, and hPSC-derived hepatic progenitors. MSC-driven condensation on Matrigel | [8] |
| Feeder-free human iPSCs | Coculture of iEC, iMSCs, and hiPSC-derived hepatic progenitors iMSC-driven condensation on Matrigel | [23] | |
| HAMECs and hiPSCs | EB differentiation of hepatic cells in the presence of HAMEC | [24] | |
| MSCs, MSC-derived hepatocytes, and HSC- and LSEC-like cells | Coculture of MSCs, MSC-derived hepatocytes, and HSC- and LSEC-like cells MSC-driven condensation on Matrigel | [25] | |
| LPCs and LSECs | Coculture of LPCs and LSECs | [26] | |
| Lentivirus vector, hiPSCs, and primary human hepatocytes | Engineering of GRN by lentiviral transduction | [28] | |
| Three-dimensional bioprinting | HepG2/C3A cells, EA.hy 926 cells, fabricated alginate solution for cell-laden bioinks and sacrificial materials, lyophilized Atelocollagen, and gelatin powder to form scaffold | Embedding of ECs on a lobule structure with microchannel built by sacrificial material to form an endothelium-lined lumen | [30] |
| Liver-bud-like spheroids generated by mature hepatocytes, HUVECs, and MSCs | Coculture of mature hepatocytes, HUVECs, and MSCs | [31] | |
| HepaRG cells, sodium alginate solution, and gelatin solution for bioink | In vivo perfusion after transplantation | [32] | |
| Decellularization | Liver tissue from mice, rats, ferrets, rabbits, and pigs, 1% Triton-X 100 with 0.1% ammonium hydroxide for decellularization, HUVECs, hFLCs | Seeding ECs on the decellularized liver with a vascular network | [36] |
| Porcine liver harvested from 5 to 8 kg piglets, 1% Triton X-100 and 0.1% ammonium hydroxide in distilled water for decellularization, rat anti-mouse CD31 antibody to improve re-endothelialization, and vascular endothelial cells expressing GFP protein (MS1) | Seeding ECs on the decellularized liver with a vascular network | [38] | |
| Porcine livers collected from adult pigs (40–50 kg), 1% Triton X-100 and 0.1% ammonium hydroxide in distilled water for decellularization, HG mixture to improve re-endothelialization, human EA.hy926 endothelial cells, hepatic carcinoma cells (HepG2 cells) | Seeding ECs on the decellularized liver with a vascular network | [39] | |
| In vivo transplantation | LBs generated from iPSC-HE cells, HUVECs and BMSCs, immunodeficient mice as hosts | Blood perfusion in host body after transplantation | [40] |
| LBs generated from LPCs and LSECs, using Fah-/-/Rag2-/-/Il2rg-/- mice as hosts | Blood perfusion in host body after transplantation | [26] | |
| LOU prepared from human liver or 2-week-old ActinGFP mice, NOD/SCID γ host mice as hosts | Blood perfusion in host body after transplantation | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Tang, L.; Yi, Q. Engineering the Vasculature of Stem-Cell-Derived Liver Organoids. Biomolecules 2021, 11, 966. https://doi.org/10.3390/biom11070966
Zhang X, Tang L, Yi Q. Engineering the Vasculature of Stem-Cell-Derived Liver Organoids. Biomolecules. 2021; 11(7):966. https://doi.org/10.3390/biom11070966
Chicago/Turabian StyleZhang, Xv, Liling Tang, and Qian Yi. 2021. "Engineering the Vasculature of Stem-Cell-Derived Liver Organoids" Biomolecules 11, no. 7: 966. https://doi.org/10.3390/biom11070966
APA StyleZhang, X., Tang, L., & Yi, Q. (2021). Engineering the Vasculature of Stem-Cell-Derived Liver Organoids. Biomolecules, 11(7), 966. https://doi.org/10.3390/biom11070966