
Redox Impact on Bacterial Macromolecule: A Promising Avenue for Discovery and Development of Novel Antibacterials


Abstract
:1. Introduction
2. Materials and Methods
2.1. Background Information on ROS Sources, Types, Generation, and Impact on Bacterial Macromolecules
2.1.1. Sources and Types of ROS in Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ROS Species | Generation Steps and Characteristics of ROS | Stepwise Reduction Equation | References |
|---|---|---|---|
| hydroperoxyl radical (HO2•) | During a series of events during respiration, molecular oxygen is reduced to hydroperoxyl radical (HO2•). | O2 + e− + H+ → HO2• | [15,25,36] |
| superoxide (O2•−) | The hydroperoxyl radical (HO2•) dissociates to form superoxide (O2•−). The reduction potential of this species is based on the environmental conditions of the solution. It can act as a mild oxidizing agent in an aqueous solution and, under other environmental conditions, it can act as a reducing agent. | HO2• → H+ + O2•− Fe2+ + O2 ↔ Fe(II)O2 ↔ Fe(III)O2− ↔ Fe3+ + O2•− | [15,25,26,36] |
| Hydrogen peroxide (H2O2) | Superoxide (O2•−) undergoes additional transformations as a result of a well-known dismutation reaction. This process is hastened by the superoxide dismutase (SOD) enzyme, which has a copper-zinc core to generate hydrogen peroxide (H2O2). However, other enzymes (e.g., urate oxidase and glucose oxidase) have been shown to catalyze this reaction. It is critical that the cells “neutralize” H2O2, since the species can be reduced through Fenton chemistry to •OH, which is lethal. | O2•− + 2H+ + e− → H2O2 2H+ + O2•− + O2•− → H2O2 + O2 | [26,32,36,37] |
| Hydroxyl radical (•OH) | A hydroxyl radical (•OH) is generated through the Fenton reaction. Of note, in live organisms and at physiological pH, the ferrous ion (Fe2+) that accelerates the Fenton reaction has a short lifespan and can quickly auto-oxidize to ferric (Fe3+). The hydroxyl radical is a highly aggressive radical that can impede the correct functioning of a variety of biological molecules. | H2O2 + e− → HO− + •OH H2O → •OH + H• + e−aq → H2O2 Fe2+ + H2O2 → Fe3+ + −OH + •OH | [25,26,36] |
| Singlet oxygen (1O2) | Singlet oxygen is considered one of the most dangerous species of ROS generated through the natural process, with huge biological significance. |
| [33,36] |
2.1.2. Defense against ROS Generation in Bacteria
2.1.3. Mechanism of Bacterial Lethality via Oxidative Stress
2.1.4. Antibiotic-Mediated ROS Lethality
Two Commonly Implicated Antibiotics in ROS-Mediated Bacterial Lethality
2.2. Natural and Synthetic Agents Contributing to ROS-Mediated Bacterial Lethality
| Basis of Opinion | Views Undermining the Opinion | Studies Solidifying the Opinion |
|---|---|---|
| Involvement of ROS in antibiotics bacterial lethality | Iron/iron-sulfur clusters’ effect on bacterial killing by antibiotics is largely dependent on how antibiotics are taken up; ROS play no role [59]. Further, ROS accumulation and cell death in an antibiotic-treated cell are discordant [21]. Hydroxyl radical (•OH) accumulation did not often correspond to antimicrobial death [21]. | Belenky et al. [60] demonstrated the involvement of ROS in antibiotic lethality by showing that cells exposed to antibiotics had cytotoxic changes including malondialdehyde adducts, protein carbonylation, double-strand DNA breaks, and nucleotide oxidation. which are indicative of ROS involvement. Further, the findings of Luan et al. [61] also laid justice to the involvement of ROS by demonstrating that katG mutants produce more ROS, which subsequently resulted in their high rate of death relative to the wild type after antibiotic treatment. |
| Type (antibiotics) and conditions that influence ROS generation | The ROS-mediated mechanism of killing is shared by all antibiotics, and anaerobiosis inhibits the lethality of norfloxacin (quinolone) at a low concentration. Furthermore, Iron chelator (dipyridyl) and hydroxyl radical scavenger (thiourea) prevented cells from being killed by antimicrobials under both aerobic and anaerobic conditions [21]. | In 2014, Dwyer et al. [17] demonstrated that antibiotics belonging to fluoroquinolones, β-lactams, and aminoglycosides generate ROS while interacting with their target sites, though information on ROS generation of other classes of antibiotics is unconvincing. Further, findings from the study showed that ampicillin, gentamicin, and norfloxacin have only attenuated lethality under highly anaerobic conditions. Furthermore, Malik et al. [62,63] showed that the choice of norfloxacin by Keren et al. [21] was not a good candidate for ROS experiments due to the intermediate reactions to anaerobic cell death caused by quinolones. |
| Quantity of ROS generated | ROS produced during antibiotic treatment are too minute to truly kill bacterial cells [22]. | The findings of Luan et al. [61] and Hong et al. [64], have, to a degree, debunked the claim on the quantity of ROS generation by demonstrating that stress can increase the number of sites available for ROS assault and that a high ROS concentration may not be required. Moreover, the findings of Luan et al. [61] and Dwyer et al. [17], through demonstration of stressors that created lesions that were hypersensitive to ROS attack, could also reduce the credibility of the argument. The actual evidence on the quantity of ROS generation relative to cell death came from Hong et al. [46] and Dorsey-Oresto et al. [54], who postulated that intracellular levels of ROS were capable of killing cells once the initial triggering stressor had been removed. |
| Detection of ROS | The low specificity of dyes for ROS detection and antibiotic therapy did not increase ROS [22]. | The claim about low specificity of dyes for ROS was addressed by Dwyer et al. [17] in a ROS quantification experiment conducted in 2014, in which they used a wide range of fluorescent dyes to identify several kinds of ROS in bacteria, including H2O2, which could not be detected with the previously utilized HPF dye (3′-(p-hydroxyphenyl) fluorescein). In their findings, bactericidal agents including ampicillin, gentamicin, and norfloxacin increased H2O2 generation in bacteria after treatment. |
| Effectiveness of exogenous antioxidant | The possibility of chemical agents such as thiourea and dipyridyl causing off-target effects cannot be fully ruled out [20,22]. | This causative argument about off-target effects of ROS scavenger could be countered by the findings of Luan et al. [61], who demonstrated that katG mutants produce more ROS, which subsequently resulted in their high rate of death relative to the wild type after antibiotic treatment. |
2.2.1. Phenolic Compounds
2.2.2. Honey
2.2.3. Carotenoid
2.2.4. Antimicrobial Peptide
2.2.5. Siderophore
2.2.6. Synthetic Agents in ROS-Mediated Bacterial Lethality
| Agent Commpound Classes | Agents | Bacteria | Implicated ROS and Mechanisms of Involvement in Antibacterial Activity | References |
|---|---|---|---|---|
| Natural agents | ||||
| Phenolics |
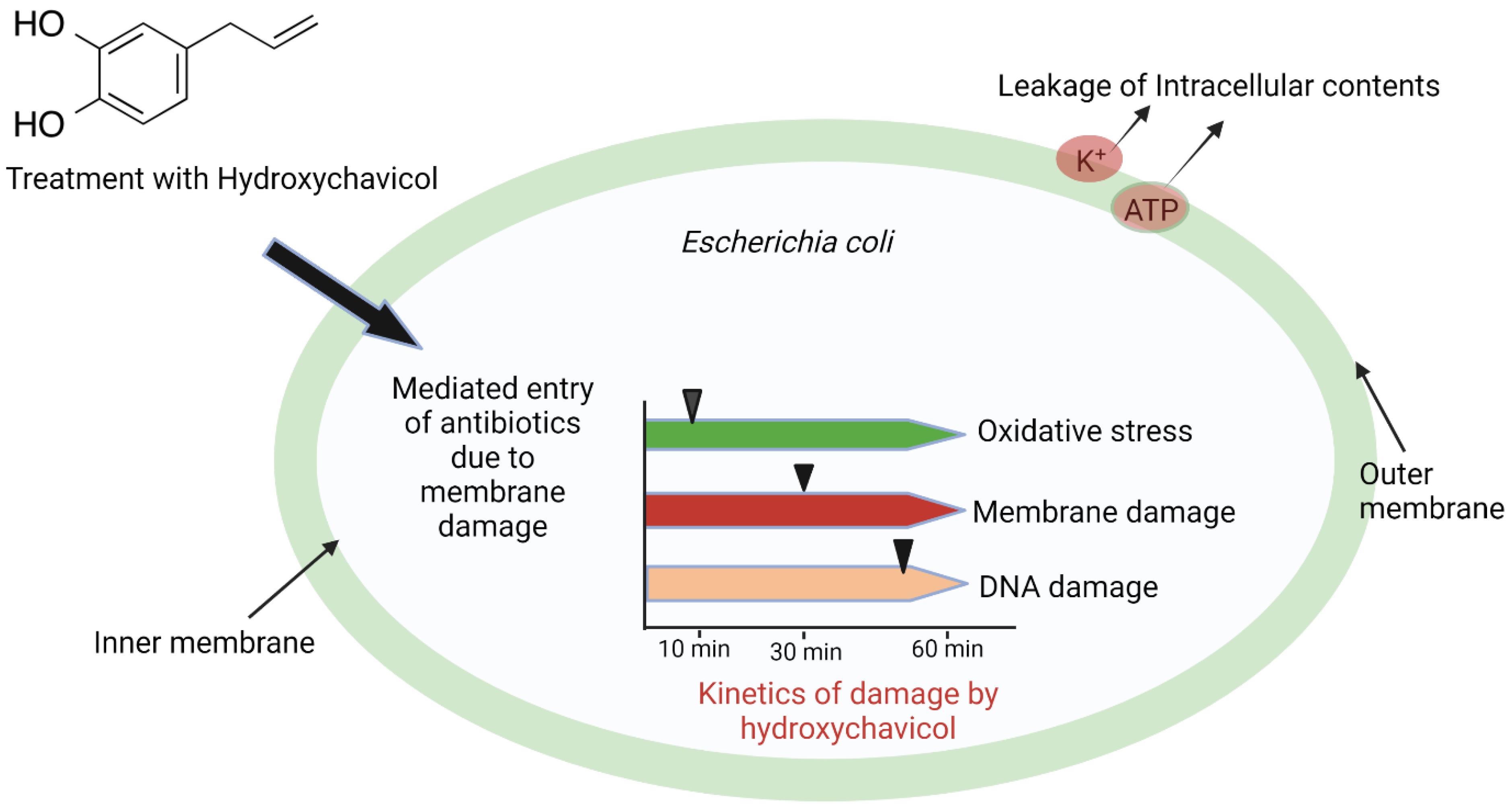
| E. coli | Time-dependent destruction of cellular macromolecules by oxidative stress caused by hydroxyl radical (•OH). Destruction of the cell membrane occurs 30 min after treatment, and DNA damage onset occurs after 30 min. | [67,68] |
| S. aureus (both methicillin-sensitive and methicillin-resistant) | Increased generation of ROS (O2•− and H2O2) resulted in the onset of oxidative stress and subsequent damage to the cell membrane, nucleic acid, and protein by hydroxyl radical (•OH). | [69] | |
| E. coli | Increased endogenous hydrogen peroxide (H2O2), leading to inhibition of E. coli growth | [71] | |
| A. baumannii (wild, sodB, and katG mutants) | Elevated superoxide anion (O2•−) production by the phenolic acids resulted in increased colistin-mediated bacterial killing via the destruction of redox homeostasis. | [70] | |
| E. coli, P. aeruginosa, and S. aureus | Protocatechuic acid instigates ROS (O2•−, H2O2, •OH) generation through Fenton chemistry, autoxidation, and hampering of electron transport chain, consequently resulting in peroxidation of lipid, breakage of DNA, and ultimate cell death of bacterial cells. | [66] | |
| S. aureus | Allyl pyrocatechol provoked internal oxidative stress in S. aureus (O2•−), thereby amplifying the transcription and activities of SODs (SodA, SodM) in S. aureus to adapt to the increased oxidative stress. | [72] | |
| Honey |
| S. aureus | Hydrogen peroxide (H2O2) | [74] |
| S. aureus | The elevated quantity of hydrogen peroxide (H2O2) resulted in higher antibacterial activity. | ||
| S. aureus | The elevated quantity of hydrogen peroxide (H2O2) resulted in higher antibacterial activity. | ||
| Carotenoid | Astaxanthin | S. aureus E. coli P. aeruginosa | ROS accumulation of superoxide anion (O2•−) and hydroxyl radical (•OH) causes a reduction in cellular glutathione level and increases the rate of bacterial death. | [75] |
| Antimicrobial peptide |
| Bacteria (K. pneumoniae, B. subtilis, S. aureus, and E. coli) and fungi (C. tropicalis and C. albicans) | Elevated accumulation of ROS (H2O2, O2•−, •OH) causes loss of cell membrane integrity and ultimate cell death. | [76] |
| Siderophore | pyochelin | E. faecalis S. aureus | Elevated ROS production (O2•−, H2O2, •OH) causes lipid peroxidation and cell death. | [77] |
| Synthetic agents | ||||
| Organic cationic salts | Bis-quaternary ammonium salt | S. aureus E. coli | Induced ROS (O2•−, H2O2, •OH) cause cytoplasm content leakage and cell membrane damage. | [80] |
| Synthetic organic cationic complexes | Derivatives of cationic heteroleptic (III) complexes with tris-diimine ligands moiety.
| S. aureus S. mutans | Photoactivation of singlet oxygen (1O2) against pathogenic bacterial cells | [78] |
| Amphiphilic copolymer | T-TCP micelles (comprising of propylene sulfide and toluidine blue O (TBO) grafted chitosan) | S. aureus L. monocytogenes | Potent antibacterial activity due to ROS (H2O2, O2•−, •OH) produced by TBO activation | [79] |
| Organotin complexes | Organotin complexes
| E. coli B. subtilis | An increase in the amount of accumulated ROS (H2O2, O2•−, •OH) caused membrane damage and leakage of cytoplasm content in the organism. | [81] |
| Modified chitosan | Catechol-modified chitosan with melanin capsule | Methicillin-resistant S. aureus (MRSA) S. aureus E. coli | Electron transfer catalyzed by Catechol-modified film from ascorbate to molecular oxygen instigated continued ROS generation (H2O2, O2•) and triggered improved antibacterial activities in vitro and in vivo. | [82] |
2.3. Safety Concerns Associated with ROS as a Contributory Antimicrobial Agent
3. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Larsson, G.D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Kohler, V.; Vaishampayan, A.; Grohmann, E. Problematic Groups of Multidrug-Resistant Bacteria and Their Resistance Mechanisms. In Antibacterial Drug Discovery to Combat; MDR; Ahmad, I., Ahmad, S., Rumbaugh, K., Eds.; Springer: Singapore, 2019. [Google Scholar]
- Malik, B.; Bhattacharyya, S. Antibiotic drug-resistance as a complex system driven by socio-economic growth and antibiotic misuse. Sci. Rep. 2019, 9, 9788. [Google Scholar]
- Nelson, R.E.; Slayton, R.B.; Stevens, V.W.; Jones, M.M.; Khader, K.; Rubin, M.A.; Jernigan, J.A.; Samore, M.H. Attributable mortality of healthcare-associated infections due to multidrug-resistant gram-negative bacteria and methicillin-resistant Staphylococcus aureus. Infect. Control Hosp. Epidemiol. 2017, 38, 848–856. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Stop Using Antibiotics in Healthy Animals to Prevent 666 the Spread of Antibiotic Resistance. Available online: https://www.who.int/news-room/detail/07-11-2017-stop-using-antibiotics-in-healthy-animals-to-prevent-the-spread-of-antibiotic-resistance (accessed on 10 September 2019).
- Rather, I.A.; Kim, B.C.; Bajpai, V.K.; Park, Y.H. Self-medication and antibiotic resistance: Crisis, current challenges, and prevention. Saudi J. Biol. Sci. 2017, 24, 808–812. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Phys. Ther. 2015, 40, 277–283. [Google Scholar]
- Lam, P.L.; Wong, R.S.M.; Lam, K.H.; Hung, L.K.; Wong, M.; Yung, L.H.; Ho, Y.W.; Wong, W.Y.; Hau, D.K.P.; Gambari, R.; et al. The Role of Reactive Oxygen Species in the Biological 1 Activity of Antimicrobial Agents: An Updated Mini Review. Chem.-Biol. Interact. 2020, 320, 109023. [Google Scholar] [CrossRef]
- Mourenza, A.; Gil, J.A.; Mateos, L.M.; Letek, M. Oxidative stress-generating antimicrobials, a novel strategy to overcome antibacterial resistance. Antioxidants 2020, 9, 361. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, C.; Jang, H.J.; Kim, B.O.; Bae, H.W.; Chung, I.Y.; Kim, E.S.; Cho, Y.H. Antibacterial strategies inspired by the oxidative stress and response networks. J. Microbiol. 2019, 57, 203–212. [Google Scholar] [CrossRef]
- Dryden, M. Reactive oxygen species: A novel antimicrobial. Int. J. Antimicrob. Agents 2018, 51, 299–303. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X. Contribution of oxidative damage to antimicrobial lethality. Antimicrob. Agents Chemother. 2009, 53, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, A.; Arce, I. Antioxidants in respiratory diseases: Basic science research and therapeutic alternatives. Clin. Res. Trials 2016, 3, 1–11. [Google Scholar] [CrossRef]
- Fanjul-Moles, M.L.; López-Riquelme, M.L. Relationship between oxidative stress, circadian rhythms, and AMD. Oxidative Med. Cell. Longev. 2016, 2016, 7420637. [Google Scholar] [CrossRef]
- Vatansever, F.; De Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species—Bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef] [Green Version]
- Vaishampayan, A.; Grohmann, E. Antimicrobials Functioning through ROS-Mediated Mechanisms: Current Insights. Microrganisms 2022, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, D.J.; Belenky, P.A.; Yang, J.H.; MacDonald, I.C.; Martell, J.D.; Takahashi, N.; Chan, C.T.Y.; Michael, A.L.; Braff, D.; Schwarz, E.G.; et al. Antibiotics induce redox-related physiological alterations as part of their lethality. Proc. Natl. Acad. Sci. USA 2014, 5, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, D.J.; Kohanski, M.A.; Hayete, B.; Collins, J.J. Gyrase inhibitors induce an oxidative damage cellular death pathway in Escherichia coli. Mol. Syst. Biol. 2007, 3, 91. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A. Diagnosing oxidative stress in bacteria: Not as easy as you might think. Curr. Opin. Microbiol. 2015, 24, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Keren, I.; Wu, Y.; Inocencio, J.; Mulchay, L.R.; Lewis, K. Killing by bactericidal antibiotics does not depend on reactive oxygen species. Science 2014, 339, 1213–1216. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Imlay, J.A. Cell death from antibiotics without the involvement of reactive oxygen species. Science 2013, 339, 1210–1213. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobritz, M.A.; Belenky, P.; Porter, C.B.; Gutierrez, A.; Yang, J.H.; Schwarz, E.G.; Dwyer, D.J.; Khalil, A.S.; Collins, J.J. Antibiotic efficacy is linked to bacterial cellular respiration. Proc. Natl. Acad. Sci. USA 2017, 112, 8173–8180. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, M.; Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.Y. Mode of action of hydrogen peroxide and other oxidizing agents: Differences between liquid and gas forms. J. Antimicrob. Chemother. 2010, 65, 2108–2115. [Google Scholar] [CrossRef] [Green Version]
- Forman, H.J.; Torres, M. Signaling by the respiratory burst in macrophages. IUBMB Life 2001, 51, 365–371. [Google Scholar] [PubMed]
- Van Acker, H.; Coenye, T. The role of reactive oxygen species in the antibiotic-mediated killing of bacteria. Trends Microbiol. 2017, 25, 456–466. [Google Scholar] [CrossRef]
- Stokes, J.M.; Lopatkin, A.J.; Lobritz, M.A.; Collins, J.J. Bacterial Metabolism and Antibiotic Efficacy. Cell Metab. 2019, 30, 251–259. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive oxygen species and neutrophil function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on neutrophil function in severe inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Langklotz, S.; Lupilova, N.; Kuhlmann, K.; Bandow, J.E.; Leichert, L.I.O. Activation of RidA chaperone function by N-chlorination. Nat. Commun. 2014, 5, 5804. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.R.; Brugarolas, P.; He, C. Redox signaling in human pathogens. Antioxid. Redox Signal. 2011, 14, 1107–1118. [Google Scholar] [CrossRef]
- Glaeser, J.; Nuss, A.M.; Berghoff, B.A.; Klug, G. Singlet oxygen stress in microorganisms. Adv. Microb. Physiol. 2011, 58, 141–173. [Google Scholar] [PubMed]
- Altman, N. The Oxygen Prescription: The Miracle of Oxidative Therapies; Healing Arts Press: Rochester, VT, USA, 2007. [Google Scholar]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, alpha-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef]
- Authen, R.L.; Davis, J.M. Oxygen toxicity and reactive oxygen species: The devil is in the Details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Kim, E.J.; Park, J.W. Control of singlet oxygen-induced oxidative damage in Escherichia coli. J. Biochem. Mol. Biol. 2002, 35, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drlica, K.; Zhao, X. Bacterial death from treatment with fluoroquinolones and other lethal stressors. Expert Rev. Anti-Infect. Ther. 2021, 19, 601–618. [Google Scholar] [CrossRef]
- Memar, M.Y.; Ghotaslou, R.; Samiei, M.; Adibkia, K. Antimicrobial use of reactive oxygen therapy: Current insights. Infect. Drug Resist. 2018, 11, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniak, P.; Laskowska, E. Antibiotics promoting oxidative stress inhibit formation of Escherichia coli biofilm via indole signaling. Res. Microbiol. 2010, 161, 847–853. [Google Scholar]
- Li, H.; Zhao, X.; Huang, Y.; Liao, B.; Cheng, L.; Ren, B. Reactive Oxygen Species in Pathogen Clearance: The Killing Mechanisms, the Adaption Response, and the Side Effects. Front. Microbiol. 2021, 11, 3610. [Google Scholar] [CrossRef]
- Akhova, A.V.; Tkachenko, A.G. Role of Secondary Oxidative Stress in the Bactericidal Action of Antibiotics. Mosc. Univ. Biol. Sci. Bull. 2020, 75, 218–223. [Google Scholar] [CrossRef]
- Yoon, S.J.; Park, J.E.; Yang, J.H.; Park, J.W. OxyR regulon controls lipid peroxidation-mediated oxidative stress in Escherichia coli. J. Biochem. Mol. Biol. 2002, 35, 297–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Mih, N.; Anand, A.; Park, J.-O.; Tan, J.; Yurkovich, I.T.; Monk, J.M.; Lloyd, C.T.; Sandberg, T.E.; Seo, S.W.; et al. Cellular responses to reactive oxygen species are predicted from molecular mechanisms. Proc. Natl. Acad. Sci. USA 2019, 116, 14368–14373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-stress bacterial cell death mediated by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2019, 116, 10064–10071. [Google Scholar] [CrossRef] [PubMed]
- Paulander, W.; Wang, Y.; Folkesson, A.; Charbon, G.; Løbner-Olesen, A.; Ingmer, H. Bactericidal antibiotics increase hydroxyphenyl fluorescein signal by altering cell morphology. PLoS ONE 2014, 9, e92231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeom, J.; Imlay, J.A.; Park, W. Iron homeostasis affects antibiotic-mediated cell death in Pseudomonas species. J. Biol. Chem. 2010, 285, 22689–22695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arriaga-Alba, M.; Rivera-Sánchez, R.; Parra-Cervantes, G.; Barro-Moreno, F.; Flores-Paz, R.; García-Jeménez, E. Antimutagenesis of beta-carotene to mutations induced by quinolone on Salmonella typhimurium. Arch. Med. Res. 2000, 31, 156–161. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Wierzbowski, J.; Cottarel, G.; Collins, J.J. Mistranslation of membrane proteins and two-component system activation trigger antibiotic-mediated cell death. Cell 2008, 135, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodkin-Gal, I.; Sat, B.; Keshet, A.; Engelberg-Kulka, H. The communication factor EDF and the toxin-antitoxin module mazEF determine the mode of action of antibiotics. PLoS Biol. 2008, 6, e319. [Google Scholar] [CrossRef]
- Couturier, M.; Bahassi, M.; Van Melderen, L. Bacterial death by DNA gyrase poisoning. Trends Microbiol. 1998, 6, 269–275. [Google Scholar] [CrossRef]
- Foti, J.J.; Devadoss, B.; Winkler, J.A.; Collins, J.J.; Walker, G.C. Oxidation of the guanine nucleotide pool underlies cell death by bactericidal antibiotics. Science 2012, 336, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Dorsey-Oresto, A.; Lu, T.; Mosel, M.; Wang, X.; Salz, T.; Drlica, K.; Zhao, X. YihE kinase is a central regulator of programmed cell death in bacteria. Cell Rep. 2013, 3, 528–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezraty, B.; Vergnes, A.; Banzhaf, M.; Duverger, Y.; Huguenot, A.; Brochado, A.R.; Su, S.-Y.; Espinosa, L.; Loiseau, L.; Py, B.; et al. Fe-S cluster biosynthesis controls uptake of aminoglycosides in a ROS-less death pathway. Science 2013, 340, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.E.; Norton, J.P.; Spivak, A.M.; Mulvey, M.A. Urinary tract infections: Current and emerging management strategies. Clin. Infect. Dis. 2013, 57, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Chua, N.G.; Zhou, Y.P.; Tan, T.T.; Lingegowda, P.B.; Lee, W.; Lim, T.P.; Teo, J.; Cai, Y.; Kwa, A.L. Polymyxin B with dual carbapenem combination therapy against carbapnemase-producing Klebsiella pneumonia. J. Infect. 2015, 70, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of Acinetobacter baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents Chemother. 2012, 56, 5642–5649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walling, C. The Nature of the Primary Oxidants in Oxidations Mediated by Metal Ions. In Oxidases and Related Redox Systems: Proceedings of the Third International Symposium; King, T.E., Mason, H.S., Morrison, M., Eds.; Pergamon Press: New York, NY, USA, 1982; pp. 85–97. [Google Scholar]
- Belenky, P.; Ye, J.D.; Porter, C.B.; Cohen, N.R.; Lobritz, M.A.; Ferrante, T.; Jain, S.; Korry, B.J.; Schwarz, E.G.; Walker, G.C.; et al. Bactericidal Antibiotics Induce Toxic Metabolic Perturbations that Lead to Cellular Damage. Cell Rep. 2015, 13, 968–980. [Google Scholar] [CrossRef] [Green Version]
- Luan, G.; Hong, Y.; Drlica, K.; Zhao, X. Suppression of Reactive Oxygen Species Accumulation Accounts for Paradoxical Bacterial Survival at High Quinolone Concentration. Antimicrob. Agents Chemother. 2018, 62, e01622-17. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Hussain, S.; Drlica, K. Effect of anaerobic growth on quinolone lethality with Escherichia coli. Antimicrob. Agents Chemother. 2007, 51, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Hoatam, G.; Chavda, K.; Kerns, R.J.; Drlica, K. Novel approach for comparing abilities of quinolones to restrict the emergence of resistant mutants during quinolone exposure. Antimicrob. Agents Chemother. 2010, 54, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Li, L.; Luan, G.; Drlica, K.; Zhao, X. Contribution of reactive oxygen species to thymineless death in Escherichia coli. Nat. Microbiol. 2017, 2, 1667–1675. [Google Scholar] [CrossRef] [Green Version]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Pharmacol. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Ajiboye, T.O.; Habibu, R.S.; Saidu, K.; Haliru, F.Z.; Ajiboye, H.O.; Aliyu, N.O.; Ibitoye, O.B.; Uwazie, J.N.; Muritala, H.F.; Bello, S.A.; et al. Involvement of oxidative stress in protocatechuic acid-mediated bacterial lethality. Microbiologyopen 2017, 6, e00472. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Narayanamoorthy, S.; Gamre, S.; Majumdar, A.G.; Goswami, M.; Gami, U.; Cherian, S.; Subramanian, M. Hydroxychavicol, a key ingredient of Piper betle induces bacterial cell death by DNA damage and inhibition of cell division. Free. Radic. Biol. Med. 2018, 120, 62–71. [Google Scholar] [CrossRef]
- Singh, D.; Majumdar, A.G.; Gamre, S. Mahesh Subramanian Membrane damage precedes DNA damage in hydroxychavicol treated E. coli cells and facilitates cooperativity with hydrophobic antibiotics. Biochimie 2021, 180, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Sinsinwar, S.; Vadivel, V. Catechin isolated from cashew nut shell exhibits antibacterial activity against clinical isolates of MRSA through ROS-mediated oxidative stress. Appl. Microbiol. Biotechnol. 2020, 104, 8279–8297. [Google Scholar] [CrossRef]
- Ajiboye, T.O.; Skiebe, E.; Wilharm, G. Phenolic acids potentiate colistin-mediated killing of Acinetobacter baumannii by inducing redox imbalance. Biomed. Pharmacother. 2018, 101, 737–744. [Google Scholar] [CrossRef]
- Xiong, L.G.; Chen, Y.J.; Tong, J.W.; Huang, J.A.; Li, J.; Gong, Y.S.; Liu, Z.H. Tea polyphenol epigallocatechin gallate inhibits Escherichia coli by increasing endogenous oxidative stress. Food Chem. 2017, 217, 196–204. [Google Scholar] [CrossRef]
- Hussain, R.M.; Abdullah, N.F.; Amom, Z. Killing of Staphylococcus aureus by allylpyrocatechol is potentiated by induction of intracellular oxidative stress and inhibition of catalase activity. J. Integr. Med. 2016, 14, 456–464. [Google Scholar] [CrossRef]
- Volk, A.P.D.; Moreland, J.G. ROS-Containing Endosomal Compartments: Implications for Signaling. Methods Enzymol. 2014, 535, 201–224. [Google Scholar]
- Cooke, J.; Dryden, M.; Patton, T.; Brennan, J.; Barrett, J. The antimicrobial activity of prototype modified honeys that generate reactive oxygen species (ROS) hydrogen peroxide. BMC Res. Notes 2015, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Aribisala, J.O.; Nkosi, S.; Idowu, K.; Nurain, I.O.; Makolomakwa, G.M.; Shode, F.O.; Sabiu, S. Astaxanthin-Mediated Bacterial Lethality: Evidence from Oxidative Stress Contribution and Molecular Dynamics Simulation. Oxidative Med. Cell. Longev. 2021, 2021, 7159652. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.T.A.; Souza, P.F.N.; Vasconcelos, M.I.; Dias, L.P.; Martins, T.F.; Van Tilburg, M.F.; Guedes, M.I.F.; Sousa, D.O.B. Mo-CBP3-PepI, Mo-CBP3-PepII, and Mo-CBP3-PepIII are synthetic antimicrobial peptides active against human pathogens by stimulating ROS generation and increasing plasma membrane permeability. Biochimie 2019, 157, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.S.; Cheow, Y.L.; Lee, S.M. The role of reactive oxygen species in the antimicrobial activity of pyochelin. J. Adv. Res. 2017, 8, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Monro, S.; Cui, P.; Yin, H.; Liu, B.; Cameron, C.G.; Xu, W.; Hetu, M.; Fuller, A.; Kilina, S.V.; et al. Heteroleptic Ir (III)N6 complexes with long-lived triplet excited states and in vitro photobiological activities. ACS Appl. Mater. Interfaces 2019, 11, 3629–3644. [Google Scholar] [CrossRef]
- Wang, Z.; Bai, H.; Lu, C.; Hou, C.; Qiu, Y.; Zhang, P.; Duan, J.; Mu, H. Light controllable chitosan micelles with ROS generation and essential oil release for the treatment of bacterial biofilm. Carbohydr. Polym. 2019, 205, 533–539. [Google Scholar] [CrossRef]
- Song, H.; Wang, Y.; Wu, J.; Gu, S.; Li, H. Fabrication of Bis-Quaternary Ammonium Salt as an Efficient Bactericidal Weapon Against Escherichia coli and Staphylococcus aureus. ACS Omega 2018, 3, 14517–14525. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wang, H.; Xia, T.; Fang, B.; Shen, Y.; Zhang, Q.; Tian, X.; Zhou, H.; Wu, J.; Tian, Y. Two-Photon-Active Organotin (IV) Complexes for Antibacterial Function and Superresolution Bacteria Imaging. Inorg. Chem. 2018, 57, 6340–6348. [Google Scholar] [CrossRef]
- Liu, H.; Qu, X.; Kim, E.; Lei, M.; Dai, K.; Tan, X.; Xu, M.; Li, J.; Liu, Y.; Shi, X.; et al. Bio inspired redox-cycling antimicrobial film for sustained generation of reactive oxygen species. Biomaterials 2018, 162, 109–122. [Google Scholar] [CrossRef]
- Lee, W.; Woo, E.R.; Lee, D.G. Effect of apigenin isolated from Aster yomena against Candida albicans: Apigenin-triggered apoptotic pathway regulated by mitochondrial calcium signaling. J. Ethnopharmacol. 2019, 231, 19–28. [Google Scholar] [CrossRef]
- Abadi, P.G.; Shirazi, F.H.; Joshaghani, M.; Moghimi, H.R. Influence of formulation of ZnO nanoblokes containing metallic ions dopants on their cytotoxicity and protective factors: An in vitro study on human skin cells exposed to UVA radiation. Toxicol. Rep. 2018, 5, 468–479. [Google Scholar] [CrossRef]
- Ali, D.; Tripathi, A.; Al Ali, H.; Shahi, Y.; Mishra, K.K.; Alarifi, S.; Alkahtane, A.A.; Manohardas, S. ROS dependent Bax/Bcl2 and caspase 3 pathway-mediated apoptosis induced by zineb in human keratinocyte cells. OncoTargets Ther. 2018, 11, 489–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.P.; Li, Z.; Choi, E.K.; Lee, S.; Kim, Y.K.; Seo, E.Y.; Chung, J.H.; Cho, S. Urban particulate 859 matter in air pollution penetrates the barrier-disrupted skin and produces ROS-dependent cutaneous inflammatory response in vivo. J. Dermatol. Sci. 2018, 91, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, B. Hydroxyl radical and its scavengers in health and disease. Oxidative Med. Cell. Longev. 2011, 2011, 809696. [Google Scholar] [CrossRef]
- Gao, F.; Shao, T.; Yu, Y.; Xiong, Y.; Yang, L. Surface-bound reactive oxygen species generating nanozymes for selective antibacterial action. Nat. Commun. 2021, 12, 745. [Google Scholar] [CrossRef]
- Kim, S.; Woo, E.R.; Lee, D.G. Synergistic Antifungal Activity of Isoquercitrin: Apoptosis and Membrane Permeabilization Related to Reactive Oxygen Species in Candida albicans. IUBMB Life 2019, 71, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.Z.; Su, Y.B.; Lin, X.M.; Lai, S.S.; Li, W.X.; Ali, F.; Zheng, J.; Peng, B. Alanine Enhances Aminoglycosides- Induced ROS Production as Revealed by Proteomic Analysis. Front. Microbiol. 2018, 9, 29. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aribisala, J.O.; Sabiu, S. Redox Impact on Bacterial Macromolecule: A Promising Avenue for Discovery and Development of Novel Antibacterials. Biomolecules 2022, 12, 1545. https://doi.org/10.3390/biom12111545
Aribisala JO, Sabiu S. Redox Impact on Bacterial Macromolecule: A Promising Avenue for Discovery and Development of Novel Antibacterials. Biomolecules. 2022; 12(11):1545. https://doi.org/10.3390/biom12111545
Chicago/Turabian StyleAribisala, Jamiu Olaseni, and Saheed Sabiu. 2022. "Redox Impact on Bacterial Macromolecule: A Promising Avenue for Discovery and Development of Novel Antibacterials" Biomolecules 12, no. 11: 1545. https://doi.org/10.3390/biom12111545
APA StyleAribisala, J. O., & Sabiu, S. (2022). Redox Impact on Bacterial Macromolecule: A Promising Avenue for Discovery and Development of Novel Antibacterials. Biomolecules, 12(11), 1545. https://doi.org/10.3390/biom12111545