
Zinc’s Association with the CmPn/CmP Signaling Network in Breast Cancer Tumorigenesis



Abstract
:1. Introduction
2. Cellular Zinc Level Is Influenced by Dietary Supplementation
2.1. Cellular Zinc Concentrations Depend on Dietary Consumption
2.2. Zinc Cellular Specific Actions
3. Zinc Plays a Significant Role in Tumorigenesis
3.1. Function of Zinc Contributes to the Progression of Cell Tumorigenesis
3.2. Zinc Transport Protein in Breast Cancer Cells
4. Zinc and the CmPn/CmP Signaling Network
4.1. Zinc Is a Critical Nutrient in Mammalian Female Reproductive System
4.2. Zinc Supplementation for Cancer Prevention
4.3. Zinc Is an Essential Nutrient in PRG Biogenesis and PRG-Mediated Signaling
4.4. Zinc Associated with the CSC
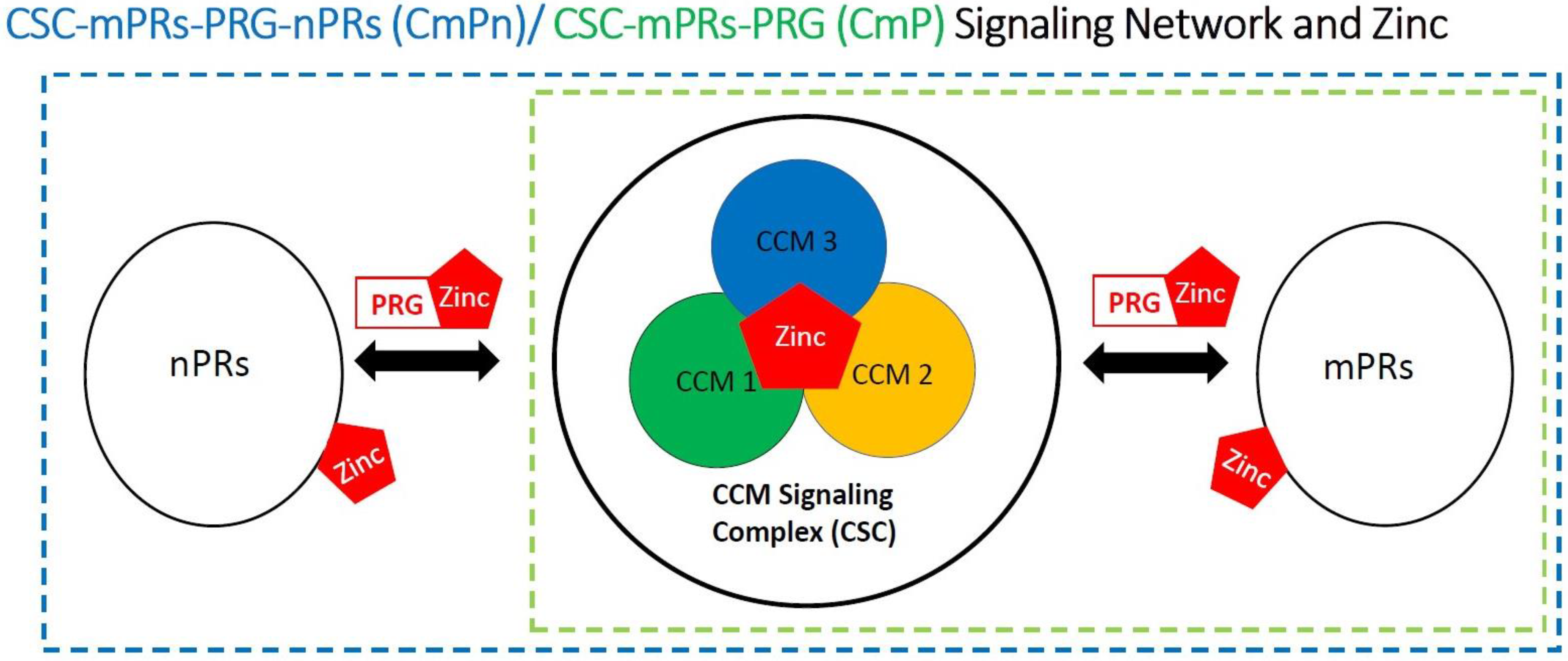
4.5. Involvement of Zinc within the CmPn/CmP Signaling Network
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bauer, K.R.; Brown, M.; Cress, R.D.; Parise, C.A.; Caggiano, V. Descriptive analysis of estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype: A population-based study from the California cancer Registry. Cancer 2007, 109, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Baquet, C.R.; Mishra, S.I.; Commiskey, P.; Ellison, G.L.; DeShields, M. Breast cancer epidemiology in blacks and whites: Disparities in incidence, mortality, survival rates and histology. J. Natl. Med. Assoc. 2008, 100, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Venturelli, S.; Leischner, C.; Helling, T.; Renner, O.; Burkard, M.; Marongiu, L. Minerals and Cancer: Overview of the Possible Diagnostic Value. Cancers 2022, 14, 1256. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Angulo, A.M.; Morales-Vasquez, F.; Hortobagyi, G.N. Overview of resistance to systemic therapy in patients with breast cancer. Adv. Exp. Med. Biol. 2007, 608, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kohler, B.A.; Sherman, R.L.; Howlader, N.; Jemal, A.; Ryerson, A.B.; Henry, K.A.; Boscoe, F.P.; Cronin, K.A.; Lake, A.; Noone, A.M.; et al. Annual Report to the Nation on the Status of Cancer, 1975–2011, Featuring Incidence of Breast Cancer Subtypes by Race/Ethnicity, Poverty, and State. J. Natl. Cancer Inst. 2015, 107, djv048. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Vail, G.; Roepke, T.A. Membrane-initiated estrogen signaling via Gq-coupled GPCR in the central nervous system. Steroids 2019, 142, 77–83. [Google Scholar] [CrossRef]
- Azeez, J.M.; Susmi, T.R.; Remadevi, V.; Ravindran, V.; Sasikumar Sujatha, A.; Ayswarya, R.N.S.; Sreeja, S. New insights into the functions of progesterone receptor (PR) isoforms and progesterone signaling. Am. J. Cancer Res. 2021, 11, 5214–5232. [Google Scholar]
- Thomas, P. Membrane Progesterone Receptors (mPRs, PAQRs): Review of Structural and Signaling Characteristics. Cells 2022, 11, 1785. [Google Scholar] [CrossRef]
- Carey, L.A.; Perou, C.M.; Livasy, C.A.; Dressler, L.G.; Cowan, D.; Conway, K.; Karaca, G.; Troester, M.A.; Tse, C.K.; Edmiston, S.; et al. Race, breast cancer subtypes, and survival in the Carolina Breast Cancer Study. JAMA 2006, 295, 2492–2502. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thurlimann, B.; Senn, H.J.; Panel, M. Strategies for subtypes—Dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef] [PubMed]
- Vogel-Gonzalez, M.; Musa-Afaneh, D.; Rivera Gil, P.; Vicente, R. Zinc Favors Triple-Negative Breast Cancer’s Microenvironment Modulation and Cell Plasticity. Int. J. Mol. Sci. 2021, 22, 9188. [Google Scholar] [CrossRef]
- Franklin, R.B.; Costello, L.C. The Important Role of the Apoptotic Effects of Zinc in the Development of Cancers. J. Cell Biochem. 2009, 106, 750–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattan, B.J.; Freake, H.C. Zinc and Cancer: Implications for LIV-1 in Breast Cancer. Nutrients 2012, 4, 648–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, K.A.; Huang, C.; Fierke, C.A. Function and mechanism of zinc metalloenzymes. J. Nutr. 2000, 130, 1437S–1446S. [Google Scholar] [CrossRef] [Green Version]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef]
- Taccioli, C.; Chen, H.; Jiang, Y.; Liu, X.P.; Huang, K.; Smalley, K.J.; Farber, J.L.; Croce, C.M.; Fong, L.Y. Dietary zinc deficiency fuels esophageal cancer development by inducing a distinct inflammatory signature. Oncogene 2012, 31, 4550–4558. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Chen, H. Aberrance of Zinc Metalloenzymes-Induced Human Diseases and Its Potential Mechanisms. Nutrients 2021, 13, 4456. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, H.; Xu, Z.; Cheng, X. Zinc dysregulation in cancers and its potential as a therapeutic target. Cancer Biol. Med. 2020, 17, 612–625. [Google Scholar] [CrossRef]
- Woo, W.; Xu, Z.M. Body zinc distribution profile during N-methyl-N-nitrosourea-induced mammary tumorigenesis in rats at various levels of dietary zinc intake. Biol. Trace Elem Res. 2002, 87, 157–169. [Google Scholar] [PubMed]
- Chakraborty, M.; Hershfinkel, M. Zinc Signaling in the Mammary Gland: For Better and for Worse. Biomedicines 2021, 9, 1204. [Google Scholar] [CrossRef] [PubMed]
- Skrajnowska, D.; Bobrowska-Korczak, B. Role of Zinc in Immune System and Anti-Cancer Defense Mechanisms. Nutrients 2019, 11, 2273. [Google Scholar] [CrossRef] [Green Version]
- Fong, L.Y.Y.; Nguyen, V.T.; Farber, J.L. Esophageal cancer prevention in zinc-deficient rats: Rapid induction of apoptosis by replenishing zinc. J. Natl. Cancer Inst. 2001, 93, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.Y.Y.; Jiang, Y.; Riley, M.; Liu, X.; Smalley, K.J.; Guttridge, D.C.; Farber, J.L. Prevention of upper aerodigestive tract cancer in zinc-deficient rodents: Inefficacy of genetic or pharmacological disruption of COX-2. Int. J. Cancer 2008, 122, 978–989. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.; Kelleher, S.L. Cellular mechanisms of zinc dysregulation: A perspective on zinc homeostasis as an etiological factor in the development and progression of breast cancer. Nutrients 2012, 4, 875–903. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, A.; Kaya, Y.; Tanriverdi, O. Effect of the Interaction Between Selenium and Zinc on DNA Repair in Association With Cancer Prevention. J. Cancer Prev. 2019, 24, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. Regulation of Cellular Zinc Ions and Their Signaling Functions. In Zinc Signaling; Fukada, T., Kambe, T., Eds.; Springer: Singapore, 2019; pp. 5–22. [Google Scholar]
- Maret, W. Zinc in Cellular Regulation: The Nature and Significance of “Zinc Signals”. Int. J. Mol. Sci. 2017, 18, 2285. [Google Scholar] [CrossRef] [Green Version]
- Colvin, R.A.; Holmes, W.R.; Fontaine, C.P.; Maret, W. Cytosolic zinc buffering and muffling: Their role in intracellular zinc homeostasis. Metallomics 2010, 2, 306–317. [Google Scholar] [CrossRef]
- Lonergan, Z.R.; Skaar, E.P. Nutrient Zinc at the Host-Pathogen Interface. Trends Biochem. Sci. 2019, 44, 1041–1056. [Google Scholar] [CrossRef]
- Chandler, P.; Kochupurakkal, B.S.; Alam, S.; Richardson, A.L.; Soybel, D.I.; Kelleher, S.L. Subtype-specific accumulation of intracellular zinc pools is associated with the malignant phenotype in breast cancer. Mol. Cancer 2016, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Margalioth, E.J.; Schenker, J.G.; Chevion, M. Copper and zinc levels in normal and malignant tissues. Cancer 1983, 52, 868–872. [Google Scholar] [CrossRef]
- Feng, Y.; Zeng, J.W.; Ma, Q.; Zhang, S.; Tang, J.; Feng, J.F. Serum copper and zinc levels in breast cancer: A meta-analysis. J. Trace Elem. Med. Biol. 2020, 62, 126629. [Google Scholar] [CrossRef] [PubMed]
- Riesop, D.; Hirner, A.V.; Rusch, P.; Bankfalvi, A. Zinc distribution within breast cancer tissue: A possible marker for histological grading? J. Cancer Res. Clin. Oncol. 2015, 141, 1321–1331. [Google Scholar] [CrossRef]
- Yamasaki, S.; Sakata-Sogawa, K.; Hasegawa, A.; Suzuki, T.; Kabu, K.; Sato, E.; Kurosaki, T.; Yamashita, S.; Tokunaga, M.; Nishida, K.; et al. Zinc is a novel intracellular second messenger. J. Cell Biol. 2007, 177, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Nagamatsu, S.; Nishito, Y.; Yuasa, H.; Yamamoto, N.; Komori, T.; Suzuki, T.; Yasui, H.; Kambe, T. Sophisticated expression responses of ZNT1 and MT in response to changes in the expression of ZIPs. Sci. Rep. 2022, 12, 7334. [Google Scholar] [CrossRef]
- Eide, D. Molecular biology of iron and zinc uptake in eukaryotes. Curr. Opin. Cell Biol. 1997, 9, 573–577. [Google Scholar] [CrossRef]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta 2000, 1465, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Cuajungco, M.P.; Ramirez, M.S.; Tolmasky, M.E. Zinc: Multidimensional Effects on Living Organisms. Biomedicines 2021, 9, 208. [Google Scholar] [CrossRef]
- Taylor, K.M.; Morgan, H.E.; Johnson, A.; Nicholson, R.I. Structure-function analysis of HKE4, a member of the new LIV-1 subfamily of zinc transporters. Biochem. J. 2004, 377, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.M.; Morgan, H.E.; Smart, K.; Zahari, N.M.; Pumford, S.; Ellis, I.O.; Robertson, J.F.; Nicholson, R.I. The emerging role of the LIV-1 subfamily of zinc transporters in breast cancer. Mol. Med. 2007, 13, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M. A distinct role in breast cancer for two LIV-1 family zinc transporters. Biochem. Soc. Trans 2008, 36, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Matsui, C.; Takatani-Nakase, T.; Hatano, Y.; Kawahara, S.; Nakase, I.; Takahashi, K. Zinc and its transporter ZIP6 are key mediators of breast cancer cell survival under high glucose conditions. FEBS Lett. 2017, 591, 3348–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogstrand, C.; Kille, P.; Nicholson, R.I.; Taylor, K.M. Zinc transporters and cancer: A potential role for ZIP7 as a hub for tyrosine kinase activation. Trends Mol. Med. 2009, 15, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Kasper, G.; Weiser, A.A.; Rump, A.; Sparbier, K.; Dahl, E.; Hartmann, A.; Wild, P.; Schwidetzky, U.; Castanos-Velez, E.; Lehmann, K. Expression levels of the putative zinc transporter LIV-1 are associated with a better outcome of breast cancer patients. Int. J. Cancer 2005, 117, 961–973. [Google Scholar] [CrossRef]
- Jones, S.; Farr, G.; Nimmanon, T.; Ziliotto, S.; Gee, J.M.W.; Taylor, K.M. The importance of targeting signalling mechanisms of the SLC39A family of zinc transporters to inhibit endocrine resistant breast cancer. Explor. Target Antitumor Ther. 2022, 3, 224–239. [Google Scholar] [CrossRef]
- Lisle, R.S.; Anthony, K.; Randall, M.A.; Diaz, F.J. Oocyte-cumulus cell interactions regulate free intracellular zinc in mouse oocytes. Reproduction 2013, 145, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Sharir, H.; Zinger, A.; Nevo, A.; Sekler, I.; Hershfinkel, M. Zinc released from injured cells is acting via the Zn2+-sensing receptor, ZnR, to trigger signaling leading to epithelial repair. J. Biol. Chem. 2010, 285, 26097–26106. [Google Scholar] [CrossRef] [Green Version]
- Hershfinkel, M. The Zinc Sensing Receptor, ZnR/GPR39, in Health and Disease. Int. J. Mol. Sci. 2018, 19, 439. [Google Scholar] [CrossRef] [Green Version]
- Ventura-Bixenshpaner, H.; Asraf, H.; Chakraborty, M.; Elkabets, M.; Sekler, I.; Taylor, K.M.; Hershfinkel, M. Enhanced ZnR/GPR39 Activity in Breast Cancer, an Alternative Trigger of Signaling Leading to Cell Growth. Sci. Rep. 2018, 8, 8119. [Google Scholar] [CrossRef]
- Mero, M.; Asraf, H.; Sekler, I.; Taylor, K.M.; Hershfinkel, M. ZnR/GPR39 upregulation of K(+)/Cl(-)-cotransporter 3 in tamoxifen resistant breast cancer cells. Cell Calcium 2019, 81, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Vogt, S.; O’Halloran, T.V.; Woodruff, T.K. Zinc availability regulates exit from meiosis in maturing mammalian oocytes. Nat. Chem. Biol. 2010, 6, 674–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, M.L.; Kong, B.Y.; Kim, A.M.; O’Halloran, T.V.; Woodruff, T.K. A zinc-dependent mechanism regulates meiotic progression in mammalian oocytes. Biol. Reprod. 2012, 86, 114. [Google Scholar] [CrossRef] [PubMed]
- Que, E.L.; Duncan, F.E.; Bayer, A.R.; Philips, S.J.; Roth, E.W.; Bleher, R.; Gleber, S.C.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc sparks induce physiochemical changes in the egg zona pellucida that prevent polyspermy. Integr. Biol. 2017, 9, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Diaz, F.J. Zinc depletion causes multiple defects in ovarian function during the periovulatory period in mice. Endocrinology 2012, 153, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Bedwal, R.S.; Bahuguna, A. Zinc, copper and selenium in reproduction. Experientia 1994, 50, 626–640. [Google Scholar] [CrossRef]
- Sandstead, H.H.; Fosmire, G.J.; McKenzie, J.M.; Halas, E.S. Zinc deficiency and brain development in the rat. Fed. Proc. 1975, 34, 86–88. [Google Scholar]
- Keen, C.L.; Hurley, L.S. Effects of zinc deficiency on prenatal and postnatal development. Neurotoxicology 1987, 8, 379–387. [Google Scholar]
- Uriu-Adams, J.Y.; Keen, C.L. Zinc and reproduction: Effects of zinc deficiency on prenatal and early postnatal development. Birth Defects Res. B Dev. Reprod. Toxicol. 2010, 89, 313–325. [Google Scholar] [CrossRef]
- Aliarabi, H.; Fadayifar, A.; Alimohamady, R.; Dezfoulian, A.H. The Effect of Maternal Supplementation of Zinc, Selenium, and Cobalt as Slow-Release Ruminal Bolus in Late Pregnancy on Some Blood Metabolites and Performance of Ewes and Their Lambs. Biol. Trace Elem. Res. 2019, 187, 403–410. [Google Scholar] [CrossRef]
- Tamura, T.; Goldenberg, R.L.; Johnston, K.E.; DuBard, M. Maternal plasma zinc concentrations and pregnancy outcome. Am. J. Clin. Nutr. 2000, 71, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grober, U.; Holzhauer, P.; Kisters, K.; Holick, M.F.; Adamietz, I.A. Micronutrients in Oncological Intervention. Nutrients 2016, 8, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalczyk, K.; Cymbaluk-Ploska, A. The Role of Zinc and Copper in Gynecological Malignancies. Nutrients 2020, 12, 3732. [Google Scholar] [CrossRef]
- Zhang, R.; Zhao, G.; Shi, H.; Zhao, X.; Wang, B.; Dong, P.; Watari, H.; Pfeffer, L.M.; Yue, J. Zinc regulates primary ovarian tumor growth and metastasis through the epithelial to mesenchymal transition. Free Radic. Biol. Med. 2020, 160, 775–783. [Google Scholar] [CrossRef]
- Huang, H.Y.; Caballero, B.; Chang, S.; Alberg, A.J.; Semba, R.D.; Schneyer, C.R.; Wilson, R.F.; Cheng, T.Y.; Vassy, J.; Prokopowicz, G.; et al. The efficacy and safety of multivitamin and mineral supplement use to prevent cancer and chronic disease in adults: A systematic review for a National Institutes of Health state-of-the-science conference. Ann. Intern. Med. 2006, 145, 372–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvie, M. Nutritional supplements and cancer: Potential benefits and proven harms. Am. Soc. Clin. Oncol. Educ. Book 2014, 34, e478–e486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, W.; Ullah, H.; Santarcangelo, C.; Di Minno, A.; Khan, H.; Daglia, M.; Arciola, C.R. Micronutrient Food Supplements in Patients with Gastro-Intestinal and Hepatic Cancers. Int. J. Mol. Sci. 2021, 22, 8014. [Google Scholar] [CrossRef]
- Grungreiff, K.; Gottstein, T.; Reinhold, D.; Blindauer, C.A. Albumin Substitution in Decompensated Liver Cirrhosis: Don’t Forget Zinc. Nutrients 2021, 13, 4011. [Google Scholar] [CrossRef]
- Lewandowska, A.; Religioni, U.; Czerw, A.; Deptala, A.; Karakiewicz, B.; Partyka, O.; Pajewska, M.; Sygit, K.; Cipora, E.; Kmiec, K.; et al. Nutritional Treatment of Patients with Colorectal Cancer. Int. J. Environ. Res. Public Health 2022, 19, 6881. [Google Scholar] [CrossRef]
- Kasprzyk, A.; Bilmin, K.; Chmielewska-Ignatowicz, T.; Pawlikowski, J.; Religioni, U.; Merks, P. The Role of Nutritional Support in Malnourished Patients With Lung Cancer. Vivo 2021, 35, 53–60. [Google Scholar] [CrossRef]
- Nett, H.; Steegmann, J.; Tollkuhn-Prott, B.; Holzle, F.; Modabber, A. A prospective randomized comparative trial evaluating postoperative nutritional intervention in patients with oral cancer. Sci. Rep. 2022, 12, 14213. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, S.P.; Burda, B.U.; Senger, C.A.; Lin, J.S.; Whitlock, E.P. Vitamin and mineral supplements in the primary prevention of cardiovascular disease and cancer: An updated systematic evidence review for the U.S. Preventive Services Task Force. Ann. Intern. Med. 2013, 159, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Boeing, H.; Stelmach-Mardas, M.; Gottschald, M.; Dietrich, S.; Hoffmann, G.; Chaimani, A. Dietary Supplements and Risk of Cause-Specific Death, Cardiovascular Disease, and Cancer: A Systematic Review and Meta-Analysis of Primary Prevention Trials. Adv. Nutr. 2017, 8, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasiadek, M.; Stragierowicz, J.; Klimczak, M.; Kilanowicz, A. The Role of Zinc in Selected Female Reproductive System Disorders. Nutrients 2020, 12, 2464. [Google Scholar] [CrossRef] [PubMed]
- Hurley, L.S.; Shrader, R.E. Abnormal development of preimplantation rat eggs after three days of maternal dietary zinc deficiency. Nature 1975, 254, 427–429. [Google Scholar] [CrossRef]
- Peters, J.M.; Wiley, L.M.; Zidenberg-Cherr, S.; Keen, C.L. Influence of short-term maternal zinc deficiency on the in vitro development of preimplantation mouse embryos. Proc. Soc. Exp. Biol. Med. 1991, 198, 561–568. [Google Scholar] [CrossRef]
- Garner, T.B.; Hester, J.M.; Carothers, A.; Diaz, F.J. Role of zinc in female reproduction. Biol. Reprod. 2021, 104, 976–994. [Google Scholar] [CrossRef]
- Derar, D.; Ali, A.; Almundarij, T.; Abd-Elmoniem, E.; Alhassun, T.; Zeitoun, M. Association between Serum Trace Elements Levels, Steroid Concentrations, and Reproductive Disorders in Ewes and Does. Vet. Med. Int. 2022, 2022, 8525089. [Google Scholar] [CrossRef]
- Sunar, F.; Gormus, Z.I.; Baltaci, A.K.; Mogulkoc, R. The effect of low dose zinc supplementation to serum estrogen and progesterone levels in post-menopausal women. Biol. Trace Elem. Res. 2008, 126 (Suppl. S1), S11–S14. [Google Scholar] [CrossRef]
- Miro, F.; Smyth, C.D.; Hillier, S.G. Development-related effects of recombinant activin on steroid synthesis in rat granulosa cells. Endocrinology 1991, 129, 3388–3394. [Google Scholar] [CrossRef]
- Chang, H.M.; Cheng, J.C.; Klausen, C.; Leung, P.C. BMP15 suppresses progesterone production by down-regulating StAR via ALK3 in human granulosa cells. Mol. Endocrinol. 2013, 27, 2093–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.; Chang, H.M.; Cheng, J.C.; Leung, P.C.; Sun, Y.P. TGF-beta1 downregulates StAR expression and decreases progesterone production through Smad3 and ERK1/2 signaling pathways in human granulosa cells. J. Clin. Endocrinol. Metab. 2014, 99, E2234–E2243. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kelleher, S.L. Molecular regulation of lactation: The complex and requisite roles for zinc. Arch. Biochem. Biophys. 2016, 611, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Brisken, C.; Park, S.; Vass, T.; Lydon, J.P.; O’Malley, B.W.; Weinberg, R.A. A paracrine role for the epithelial progesterone receptor in mammary gland development. Proc. Natl. Acad. Sci. USA 1998, 95, 5076–5081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obr, A.E.; Edwards, D.P. The biology of progesterone receptor in the normal mammary gland and in breast cancer. Mol. Cell Endocrinol. 2012, 357, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Habib, F.K.; Maddy, S.Q.; Stitch, S.R. Zinc induced changes in the progesterone binding properties of the human endometrium. Eur. J. Endocrinol. 1980, 94, 99–106. [Google Scholar] [CrossRef]
- Vasiliauskaite-Brooks, I.; Sounier, R.; Rochaix, P.; Bellot, G.; Fortier, M.; Hoh, F.; De Colibus, L.; Bechara, C.; Saied, E.M.; Arenz, C.; et al. Structural insights into adiponectin receptors suggest ceramidase activity. Nature 2017, 544, 120–123. [Google Scholar] [CrossRef] [Green Version]
- Kelder, J.; Pang, Y.; Dong, J.; Schaftenaar, G.; Thomas, P. Molecular modeling, mutational analysis and steroid specificity of the ligand binding pocket of mPRalpha (PAQR7): Shared ligand binding with AdipoR1 and its structural basis. J. Steroid. Biochem. Mol. Biol. 2022, 219, 106082. [Google Scholar] [CrossRef]
- Padarti, A.; Zhang, J. Recent advances in cerebral cavernous malformation research. Vessel Plus 2018, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Abou-Fadel, J.; Vasquez, M.; Grajeda, B.; Ellis, C.; Zhang, J. Systems-wide analysis unravels the new roles of CCM signal complex (CSC). Heliyon 2019, 5, e02899. [Google Scholar] [CrossRef] [Green Version]
- Abou-Fadel, J.; Smith, M.; Falahati, K.; Zhang, J. Comparative omics of CCM signaling complex (CSC). Chin. Neurosurg. J. 2020, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Fadel, J.; Zhang, J. Systems Wide Analysis of CCM Signaling Complex Alterations in CCM-Deficient Models Using Omics Approaches. Methods Mol. Biol. 2020, 2152, 325–344. [Google Scholar] [CrossRef] [PubMed]
- Abou-Fadel, J.; Bhalli, M.; Grajeda, B.; Zhang, J. CmP Signaling Network Leads to Identification of Prognostic Biomarkers for Triple-Negative Breast Cancer in Caucasian Women. Genet Test Mol. Biomark. 2022, 26, 198–219. [Google Scholar] [CrossRef]
- Abou-Fadel, J.; Grajeda, B.; Jiang, X.; Cailing-De La, O.A.; Flores, E.; Padarti, A.; Bhalli, M.; Le, A.; Zhang, J. CmP signaling network unveils novel biomarkers for triple negative breast cancer in African American women. Cancer Biomark 2022, Preprint. [Google Scholar] [CrossRef]
- Abou-Fadel, J.; Jiang, X.; Grajeda, B.; Padarti, A.; Ellis, C.C.; Flores, E.; Cailing-De La, O.A.; Zhang, J. CCM signaling complex (CSC) couples both classic and non-classic Progesterone receptor signaling. Cell Commun. Signal 2022, 20, 120. [Google Scholar] [CrossRef] [PubMed]
- Sesterhenn, A.M.; Iwinska-Zelder, J.; Dalchow, C.V.; Bien, S.; Werner, J.A. Acute haemorrhage in patients with advanced head and neck cancer: Value of endovascular therapy as palliative treatment option. J. Laryngol. Otol. 2006, 120, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.B.; Shi, H.B.; Park, S.; Lee, D.G.; Shim, J.H.; Lee, D.H.; Suh, D.C. Acute bleeding in the head and neck: Angiographic findings and endovascular management. Am. J. Neuroradiol. 2014, 35, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Franklin, R.B.; Feng, P.; Milon, B.; Desouki, M.M.; Singh, K.K.; Kajdacsy-Balla, A.; Bagasra, O.; Costello, L.C. hZIP1 zinc uptake transporter down regulation and zinc depletion in prostate cancer. Mol. Cancer 2005, 4, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Meerarani, P.; Ramadass, P.; Toborek, M.; Bauer, H.-C.; Bauer, H.; Hennig, B. Zinc protects against apoptosis of endothelial cells induced by linoleic acid and tumor necrosis factor α. Am. J. Clin. Nutr. 2000, 71, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Karadas, S.; Sayin, R.; Aslan, M.; Gonullu, H.; Kati, C.; Dursun, R.; Duran, L.; Gonullu, E.; Demir, H. Serum Levels of Trace Elements and Heavy Metals in Patients with Acute Hemorrhagic Stroke. J. Membrane Biol. 2014, 247, 175–180. [Google Scholar] [CrossRef]
- Mammadova-Bach, E.; Braun, A. Zinc Homeostasis in Platelet-Related Diseases. Int. J. Mol. Sci. 2019, 20, 5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grungreiff, K.; Gottstein, T.; Reinhold, D. Zinc Deficiency-An Independent Risk Factor in the Pathogenesis of Haemorrhagic Stroke? Nutrients 2020, 12, 3548. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Rigamonti, D.; Badr, A.; Zhang, J. Ccm1 regulates microvascular morphogenesis during angiogenesis. J. Vasc. Res. 2011, 48, 130–140. [Google Scholar] [CrossRef]
- Chapman, E.M.; Lant, B.; Ohashi, Y.; Yu, B.; Schertzberg, M.; Go, C.; Dogra, D.; Koskimaki, J.; Girard, R.; Li, Y.; et al. A conserved CCM complex promotes apoptosis non-autonomously by regulating zinc homeostasis. Nat. Commun. 2019, 10, 1791. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Greiss, S.; Gartner, A.; Derry, W.B. Cell-nonautonomous regulation of C. elegans germ cell death by kri-1. Curr. Biol. 2010, 20, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Maddaluno, L.; Rudini, N.; Cuttano, R.; Bravi, L.; Giampietro, C.; Corada, M.; Ferrarini, L.; Orsenigo, F.; Papa, E.; Boulday, G.; et al. EndMT contributes to the onset and progression of cerebral cavernous malformations. Nature 2013, 498, 492–496. [Google Scholar] [CrossRef]
- Zhou, Z.; Rawnsley, D.R.; Goddard, L.M.; Pan, W.; Cao, X.-J.; Jakus, Z.; Zheng, H.; Yang, J.; Arthur, J.; Simon, C.; et al. The Cerebral Cavernous Malformation Pathway Controls Cardiac Development via Regulation of Endocardial MEKK3 Signaling and KLF Expression. Developmental. Cell 2015, 32, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Tang, A.T.; Wong, W.-Y.; Bamezai, S.; Goddard, L.M.; Shenkar, R.; Zhou, S.; Yang, J.; Wright, A.C.; Foley, M.; et al. Cerebral cavernous malformations arise from endothelial gain of MEKK3–KLF2/4 signalling. Nature 2016, 532, 122–126. [Google Scholar] [CrossRef] [Green Version]
- .Cunha, S.I.; Magnusson, P.U.; Dejana, E.; Lampugnani, M.G. Deregulated TGF-β/BMP Signaling in Vascular Malformations. Circ. Res. 2017, 121, 981–999. [Google Scholar] [CrossRef]
- Lopez-Ramirez, M.A.; Fonseca, G.; Zeineddine, H.A.; Girard, R.; Moore, T.; Pham, A.; Cao, Y.; Shenkar, R.; de Kreuk, B.-J.; Lagarrigue, F.; et al. Thrombospondin1 (TSP1) replacement prevents cerebral cavernous malformations. J. Exp. Med. 2017, 214, 3331–3346. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.P.; Wang, R.; Yang, X.; Wang, X.; Wang, L.; Ting, K.K.; Foley, M.; Cogger, V.; Yang, Z.; Liu, F.; et al. Ponatinib (AP24534) inhibits MEKK3-KLF signaling and prevents formation and progression of cerebral cavernous malformations. Sci. Adv. 2018, 4, eaau0731. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Ramirez, M.A.; Pham, A.; Girard, R.; Wyseure, T.; Hale, P.; Yamashita, A.; Koskimaki, J.; Polster, S.; Saadat, L.; Romero, I.A.; et al. Cerebral cavernous malformations form an anticoagulant vascular domain in humans and mice. Blood 2019, 133, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Su, V.L.; Calderwood, D.A. Signalling through cerebral cavernous malformation protein networks. Open Biol. 2020, 10, 200263. [Google Scholar] [CrossRef]
- Wei, S.; Li, Y.; Polster, S.P.; Weber, C.R.; Awad, I.A.; Shen, L. Cerebral Cavernous Malformation Proteins in Barrier Maintenance and Regulation. Int. J. Mol. Sci. 2020, 21, 675. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Ramirez, M.A.; Lai, C.C.; Soliman, S.I.; Hale, P.; Pham, A.; Estrada, E.J.; McCurdy, S.; Girard, R.; Verma, R.; Moore, T.; et al. Astrocytes propel neurovascular dysfunction during cerebral cavernous malformation lesion formation. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Jiang, X.; Padarti, A.; Qu, Y.; Sheng, S.; Abou-Fadel, J.; Badr, A.; Zhang, J. Alternatively spliced isoforms reveal a novel type of PTB domain in CCM2 protein. Sci. Rep. 2019, 9, 15808–15819. [Google Scholar] [CrossRef] [Green Version]
- Antwi-Adjei, E.; Burguete, A.S.; Ghabrial, A.S. Furry is a component of the CCM3-GCKIII signaling pathway. Vessel Plus 2021, 5, 35. [Google Scholar] [CrossRef]
- Zhang, J. Learn from the past, review the present, and look towards the future. Vessel Plus 2022, 6, 20. [Google Scholar] [CrossRef]
- Berman, J.R.; Kenyon, C. Germ-cell loss extends C. elegans life span through regulation of DAF-16 by kri-1 and lipophilic-hormone signaling. Cell 2006, 124, 1055–1068. [Google Scholar] [CrossRef] [Green Version]
- Yamawaki, T.M.; Arantes-Oliveira, N.; Berman, J.R.; Zhang, P.; Kenyon, C. Distinct activities of the germline and somatic reproductive tissues in the regulation of Caenorhabditis elegans’ longevity. Genetics 2008, 178, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Baumbach, G.L.; Didion, S.P.; Faraci, F.M. Hypertrophy of cerebral arterioles in mice deficient in expression of the gene for CuZn superoxide dismutase. Stroke 2006, 37, 1850–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didion, S.P.; Kinzenbaw, D.A.; Schrader, L.I.; Faraci, F.M. Heterozygous CuZn superoxide dismutase deficiency produces a vascular phenotype with aging. Hypertension 2006, 48, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didion, S.P.; Kinzenbaw, D.A.; Faraci, F.M. Critical role for CuZn-superoxide dismutase in preventing angiotensin II-induced endothelial dysfunction. Hypertension 2005, 46, 1147–1153. [Google Scholar] [CrossRef]
- Didion, S.P.; Ryan, M.J.; Didion, L.A.; Fegan, P.E.; Sigmund, C.D.; Faraci, F.M. Increased superoxide and vascular dysfunction in CuZnSOD-deficient mice. Circ. Res. 2002, 91, 938–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita-Fujimura, Y.; Fujimura, M.; Gasche, Y.; Copin, J.C.; Chan, P.H. Overexpression of copper and zinc superoxide dismutase in transgenic mice prevents the induction and activation of matrix metalloproteinases after cold injury-induced brain trauma. J Cerebr. Blood Flow Metab. 2000, 20, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Goitre, L.; Balzac, F.; Degani, S.; Degan, P.; Marchi, S.; Pinton, P.; Retta, S.F. KRIT1 regulates the homeostasis of intracellular reactive oxygen species. PLoS ONE 2010, 5, e11786. [Google Scholar] [CrossRef] [Green Version]
- Goitre, L.; De Luca, E.; Braggion, S.; Trapani, E.; Guglielmotto, M.; Biasi, F.; Forni, M.; Moglia, A.; Trabalzini, L.; Retta, S.F. KRIT1 loss of function causes a ROS-dependent upregulation of c-Jun. Free Radic. Biol. Med. 2014, 68, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.H.; Han, D.M.; Jeong, S.G.; Choi, M.R.; Chai, Y.G.; Cho, G.W. Proteomic analysis reveals KRIT1 as a modulator for the antioxidant effects of valproic acid in human bone-marrow mesenchymal stromal cells. Drug Chem. Toxicol. 2015, 38, 286–292. [Google Scholar] [CrossRef]
- Trapani, E.; Retta, S.F. Cerebral cavernous malformation (CCM) disease: From monogenic forms to genetic susceptibility factors. J. Neurosurg Sci. 2015, 59, 201–209. [Google Scholar]
- Retta, S.F.; Glading, A.J. Oxidative stress and inflammation in cerebral cavernous malformation disease pathogenesis: Two sides of the same coin. Int. J. Biochem. Cell Biol. 2016, 81, 254–270. [Google Scholar] [CrossRef] [Green Version]
- Fontes-Dantas, F.L.; da Fontoura Galvao, G.; Veloso da Silva, E.; Alves-Leon, S.; Cecilia da Silva Rego, C.; Garcia, D.G.; Marques, S.A.; Blanco Martinez, A.M.; Reis da Silva, M.; Marcondes de Souza, J. Novel CCM1 (KRIT1) Mutation Detection in Brazilian Familial Cerebral Cavernous Malformation: Different Genetic Variants in Inflammation, Oxidative Stress, and Drug Metabolism Genes Affect Disease Aggressiveness. World Neurosurg. 2020, 138, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Perrelli, A.; Retta, S.F. Fluorescence Analysis of Reactive Oxygen Species (ROS) in Cellular Models of Cerebral Cavernous Malformation Disease. Methods Mol. Biol. 2020, 2152, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Swamy, H.; Glading, A.J. Contribution of protein-protein interactions to the endothelial-barrier-stabilizing function of KRIT1. J. Cell Sci. 2022, 135, jcs258816. [Google Scholar] [CrossRef] [PubMed]
- Abou-Fadel, J.; Jiang, X.; Padarti, A.; Goswami, D.; Smith, M.; Grajeda, B.; Walker, W.; Zhang, J. CCM signaling complex (CSC) is a master regulator governing homeostasis of progestins and their mediated signaling cascades. bioRxiv 2020. [Google Scholar] [CrossRef]
- Abou-Fadel, J.; Qu, Y.; Gonzalez, E.; Smith, M.; Zhang, J. Emerging roles of CCM genes during tumorigenesis with potential application as novel biomarkers across major types of cancers. Oncol. Rep. 2020, 43, 1945–1963. [Google Scholar] [CrossRef] [Green Version]
- Abou-Fadel, J.; Jiang, X.; Padarti, A.; Goswami, D.G.; Smith, M.; Grajeda, B.; Bhalli, M.; Le, A.; Walker, W.E.; Zhang, J. mPR-Specific Actions Influence Maintenance of the Blood–Brain Barrier (BBB). Int. J. Mol. Sci. 2022, 23, 9684. [Google Scholar]
- Renteria, M.; Belkin, O.; Jang, D.; Aickareth, J.; Bhalli, M.; Zhang, J. CmPn signaling networks in the tumorigenesis of breast cancer. Front. Endocrinol. 2022, 13, 1013892. [Google Scholar] [CrossRef]


{kind=link}
| Main Point | Key Findings | Pubmed | References |
|---|---|---|---|
| The CSC is linked to tumorigenesis with mPRs in various cancers. | Differential expression of CCM and mPR correlated with various types and grades of major human cancers, especially breast and liver cancers. | PMID: 32186778 | [136,138] |
| Establishing the CmPn signaling network in nPR(+) breast cancers | The CSC role in coupling classic, non-classic, or combined PRG signaling pathways via the effects of an intricate homeostatic concentration of progesterone to form the CmPn signaling network | PMID: 35971177 | [96,138] |
| Establishing the CmP signaling network in nPR(-) breast cancers | Through establishingCmP signaling network in nPR(-) breast cancers, we discovered novel biomarker signature panels for Triple-Negative Breast Cancers (TNBCs) between African and Caucasian Women | PMID: 35431232; 35481969 | [94,95,138] |
| Establishing the CmP signaling network in nPR(-) vasular ECs | Deficiency of any CCM genes, in combination with mPR-specific PRG actions, leads to perturbed CmP signaling network in nPR(-) ECs both in vitro and in vivo, result in compromising blood brain barrier integrity and increases the risk of hemorrhage | PMID: 36077089; 35098046 | [135,137,138] |
| Exploring molecuular signaling within the CmPn/CmP signaling networks with multiomics | molecuular signaling within the CmPn/CmP signaling networks were investigated with multiomics, such as RNAseq and proteomics. Major molecular pathways were presented with pathway analysis and visualization | PMID: 36077089; 35098046 | [91,94,95,96,135,136] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renteria, M.; Belkin, O.; Aickareth, J.; Jang, D.; Hawwar, M.; Zhang, J. Zinc’s Association with the CmPn/CmP Signaling Network in Breast Cancer Tumorigenesis. Biomolecules 2022, 12, 1672. https://doi.org/10.3390/biom12111672
Renteria M, Belkin O, Aickareth J, Jang D, Hawwar M, Zhang J. Zinc’s Association with the CmPn/CmP Signaling Network in Breast Cancer Tumorigenesis. Biomolecules. 2022; 12(11):1672. https://doi.org/10.3390/biom12111672
Chicago/Turabian StyleRenteria, Mellisa, Ofek Belkin, Justin Aickareth, David Jang, Majd Hawwar, and Jun Zhang. 2022. "Zinc’s Association with the CmPn/CmP Signaling Network in Breast Cancer Tumorigenesis" Biomolecules 12, no. 11: 1672. https://doi.org/10.3390/biom12111672
APA StyleRenteria, M., Belkin, O., Aickareth, J., Jang, D., Hawwar, M., & Zhang, J. (2022). Zinc’s Association with the CmPn/CmP Signaling Network in Breast Cancer Tumorigenesis. Biomolecules, 12(11), 1672. https://doi.org/10.3390/biom12111672