


The State of the Dopaminergic and Glutamatergic Systems in the Valproic Acid Mouse Model of Autism Spectrum Disorder

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Tyrosine Hydroxylase (TH) Immunohistochemistry at the Ventral Mesencephalon Level
2.3. Chromatography and Spectrometry Analysis
2.4. Quantitative Immunoblot Analysis
2.5. QPCR Analysis
2.6. Data Analysis
3. Results
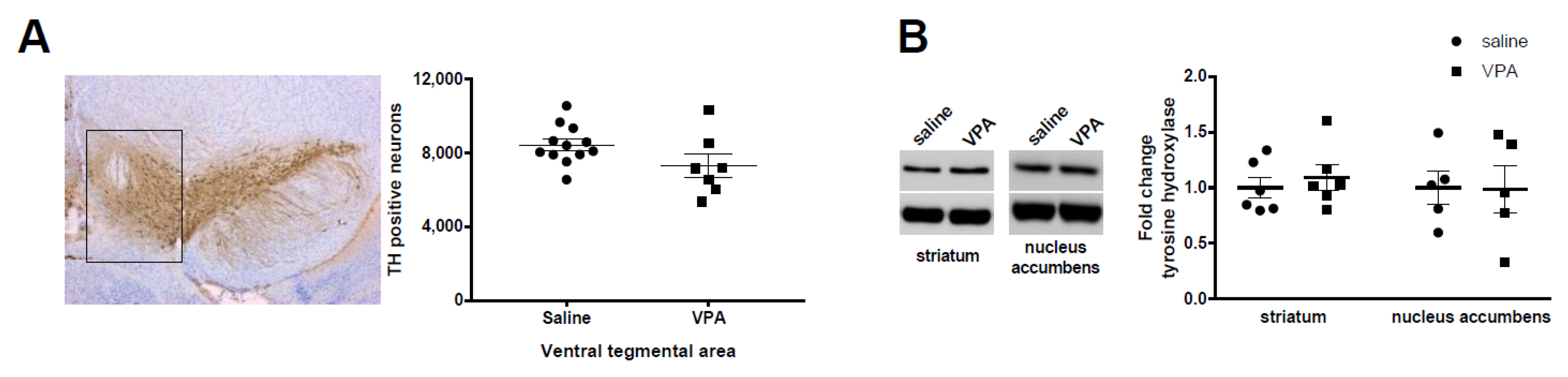
3.1. Dopamine Neurons and Tyrosine Hydroxylase Levels Are Not Altered by VPA Treatment
3.2. Dopamine and Metabolites Levels Are Altered by VPA Exposure
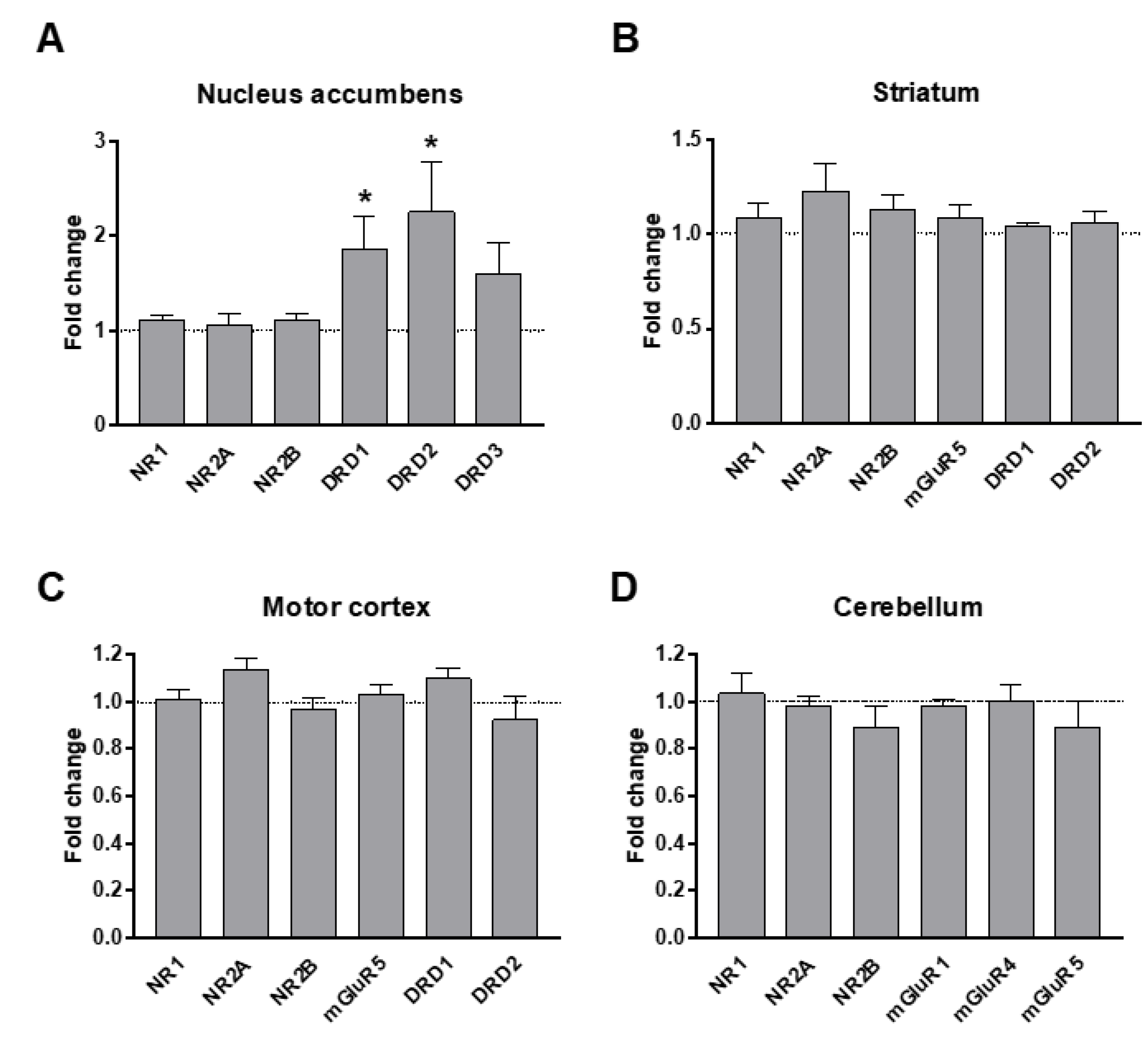
3.3. Dopamine and Glutamate Receptors’ Levels in Response to VPA Exposure
3.4. Signal Transduction Pathways within the Dopaminergic and Glutamatergic Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autism Spectrum Disorder. Lancet 2018, 392, 508–520. [Google Scholar] [CrossRef]
- Vorstman, J.A.S.; Parr, J.R.; Moreno-De-Luca, D.; Anney, R.J.L.; Nurnberger, J.I., Jr.; Hallmayer, J.F. Autism Genetics: Opportunities and Challenges for Clinical Translation. Nat. Rev. Genet. 2017, 18, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Chaste, P.; Leboyer, M. Autism Risk Factors: Genes, Environment, and Gene-Environment Interactions. Dialogues Clin. Neurosci. 2012, 14, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.A.; Peñagarikano, O.; Belgard, T.G.; Swarup, V.; Geschwind, D.H. The Emerging Picture of Autism Spectrum Disorder: Genetics and Pathology. Annu. Rev. Pathol. 2015, 10, 111–144. [Google Scholar] [CrossRef] [PubMed]
- Bölte, S.; Girdler, S.; Marschik, P.B. The Contribution of Environmental Exposure to the Etiology of Autism Spectrum Disorder. Cell Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.-Z.; Zhan, X.-L.; Jin, C.-Y.; Liang, J.-H.; Jing, J.; Dong, G.-H. The Epidemiological Evidence Linking Exposure to Ambient Particulate Matter with Neurodevelopmental Disorders: A Systematic Review and Meta-Analysis. Environ. Res. 2022, 209, 112876. [Google Scholar] [CrossRef]
- Miani, A.; Imbriani, G.; De Filippis, G.; De Giorgi, D.; Peccarisi, L.; Colangelo, M.; Pulimeno, M.; Castellone, M.D.; Nicolardi, G.; Logroscino, G.; et al. Autism Spectrum Disorder and Prenatal or Early Life Exposure to Pesticides: A Short Review. Int. J. Environ. Res. Public Health 2021, 18, 10991. [Google Scholar] [CrossRef]
- Perera, F.; Herbstman, J. Prenatal Environmental Exposures, Epigenetics, and Disease. Reprod. Toxicol. 2011, 31, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Al Sagheer, T.; Haida, O.; Balbous, A.; Francheteau, M.; Matas, E.; Fernagut, P.-O.; Jaber, M. Motor Impairments Correlate with Social Deficits and Restricted Neuronal Loss in an Environmental Model of Autism. Int. J. Neuropsychopharmacol. 2018, 21, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Haida, O.; Al Sagheer, T.; Balbous, A.; Francheteau, M.; Matas, E.; Soria, F.; Fernagut, P.O.; Jaber, M. Sex-Dependent Behavioral Deficits and Neuropathology in a Maternal Immune Activation Model of Autism. Transl. Psychiatry 2019, 9, 124. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Aldinger, K.A.; Ashwood, P.; Bauman, M.L.; Blaha, C.D.; Blatt, G.J.; Chauhan, A.; Chauhan, V.; Dager, S.R.; Dickson, P.E.; et al. Consensus Paper: Pathological Role of the Cerebellum in Autism. Cerebellum 2012, 11, 777–807. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.R.; Hewedi, D.H.; Eissa, A.M.; Moustafa, A.A. The Cerebellum and Psychiatric Disorders. Front. Public Health 2015, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Nisar, S.; Bhat, A.A.; Masoodi, T.; Hashem, S.; Akhtar, S.; Ali, T.A.; Amjad, S.; Chawla, S.; Bagga, P.; Frenneaux, M.P.; et al. Genetics of Glutamate and Its Receptors in Autism Spectrum Disorder. Mol. Psychiatry 2022, 27, 2380–2392. [Google Scholar] [CrossRef]
- Pavăl, D.; Micluția, I.V. The Dopamine Hypothesis of Autism Spectrum Disorder Revisited: Current Status and Future Prospects. Dev. Neurosci. 2021, 43, 73–83. [Google Scholar] [CrossRef]
- Carlson, G.C. Glutamate Receptor Dysfunction and Drug Targets across Models of Autism Spectrum Disorders. Pharmacol. Biochem. Behav. 2012, 100, 850–854. [Google Scholar] [CrossRef] [Green Version]
- Montanari, M.; Martella, G.; Bonsi, P.; Meringolo, M. Autism Spectrum Disorder: Focus on Glutamatergic Neurotransmission. Int. J. Mol. Sci. 2022, 23, 3861. [Google Scholar] [CrossRef]
- Carlsson, M.L. Hypothesis: Is Infantile Autism a Hypoglutamatergic Disorder? Relevance of Glutamate—Serotonin Interactions for Pharmacotherapy. J. Neural. Transm. 1998, 105, 525. [Google Scholar] [CrossRef]
- Fatemi, S.H. The Hyperglutamatergic Hypothesis of Autism. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 911. [Google Scholar] [CrossRef]
- Moreno-Fuenmayor, H.; Borjas, L.; Arrieta, A.; Valera, V.; Socorro-Candanoza, L. Plasma Excitatory Amino Acids in Autism. Investig. Clin. 1996, 37, 113–128. [Google Scholar]
- Cai, J.; Ding, L.; Zhang, J.-S.; Xue, J.; Wang, L.-Z. Elevated Plasma Levels of Glutamate in Children with Autism Spectrum Disorders. NeuroReport 2016, 27, 272–276. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhu, T.; Qu, Y.; Mu, D. Blood Glutamate Levels in Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158688. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-W.; Park, K.; Kang, R.J.; Gonzales, E.L.T.; Kim, D.G.; Oh, H.A.; Seung, H.; Ko, M.J.; Kwon, K.J.; Kim, K.C.; et al. Pharmacological Modulation of AMPA Receptor Rescues Social Impairments in Animal Models of Autism. Neuropsychopharmacology 2019, 44, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhondzadeh, S.; Tajdar, H.; Mohammadi, M.-R.; Mohammadi, M.; Nouroozinejad, G.-H.; Shabstari, O.L.; Ghelichnia, H.-A. A Double-Blind Placebo Controlled Trial of Piracetam Added to Risperidone in Patients with Autistic Disorder. Child Psychiatry Hum. Dev. 2008, 39, 237–245. [Google Scholar] [CrossRef]
- Won, H.; Lee, H.-R.; Gee, H.Y.; Mah, W.; Kim, J.-I.; Lee, J.; Ha, S.; Chung, C.; Jung, E.S.; Cho, Y.S.; et al. Autistic-like Social Behaviour in Shank2-Mutant Mice Improved by Restoring NMDA Receptor Function. Nature 2012, 486, 261–265. [Google Scholar] [CrossRef]
- Eltokhi, A.; Santuy, A.; Merchan-Perez, A.; Sprengel, R. Glutamatergic Dysfunction and Synaptic Ultrastructural Alterations in Schizophrenia and Autism Spectrum Disorder: Evidence from Human and Rodent Studies. Int. J. Mol. Sci. 2020, 22, 59. [Google Scholar] [CrossRef]
- Lee, E.; Lee, J.; Kim, E. Excitation/Inhibition Imbalance in Animal Models of Autism Spectrum Disorders. Biol. Psychiatry 2017, 81, 838–847. [Google Scholar] [CrossRef] [Green Version]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in Motivational Control: Rewarding, Aversive, and Alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef] [Green Version]
- Báez-Mendoza, R.; Schultz, W. The Role of the Striatum in Social Behavior. Front. Neurosci. 2013, 7, 233. [Google Scholar] [CrossRef] [Green Version]
- Ernst, M.; Zametkin, A.; Matochik, J.; Pascualvaca, D.; Cohen, R. Low Medial Prefrontal Dopaminergic Activity in Autistic Children. Lancet 1997, 350, 638. [Google Scholar] [CrossRef]
- Abbott, A.E.; Linke, A.C.; Nair, A.; Jahedi, A.; Alba, L.A.; Keown, C.L.; Fishman, I.; Müller, R.-A. Repetitive Behaviors in Autism Are Linked to Imbalance of Corticostriatal Connectivity: A Functional Connectivity MRI Study. Soc. Cogn. Affect. Neurosci. 2018, 13, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, A.; Kelly, C.; Grzadzinski, R.; Zuo, X.-N.; Mennes, M.; Mairena, M.A.; Lord, C.; Castellanos, F.X.; Milham, M.P. Aberrant Striatal Functional Connectivity in Children with Autism. Biol. Psychiatry 2011, 69, 847–856. [Google Scholar] [CrossRef]
- Schuetze, M.; Park, M.T.M.; Cho, I.Y.; MacMaster, F.P.; Chakravarty, M.M.; Bray, S.L. Morphological Alterations in the Thalamus, Striatum, and Pallidum in Autism Spectrum Disorder. Neuropsychopharmacology 2016, 41, 2627–2637. [Google Scholar] [CrossRef] [Green Version]
- Akkermans, S.E.A.; Rheinheimer, N.; Bruchhage, M.M.K.; Durston, S.; Brandeis, D.; Banaschewski, T.; Boecker-Schlier, R.; Wolf, I.; Williams, S.C.R.; Buitelaar, J.K.; et al. Frontostriatal Functional Connectivity Correlates with Repetitive Behaviour across Autism Spectrum Disorder and Obsessive–Compulsive Disorder. Psychol. Med. 2019, 49, 2247–2255. [Google Scholar] [CrossRef]
- DiCarlo, G.E.; Wallace, M.T. Modeling Dopamine Dysfunction in Autism Spectrum Disorder: From Invertebrates to Vertebrates. Neurosci. Biobehav. Rev. 2022, 133, 104494. [Google Scholar] [CrossRef]
- Hulbert, S.W.; Jiang, Y.-H. Monogenic Mouse Models of Autism Spectrum Disorders: Common Mechanisms and Missing Links. Neuroscience 2016, 321, 3–23. [Google Scholar] [CrossRef]
- Nicolini, C.; Fahnestock, M. The Valproic Acid-Induced Rodent Model of Autism. Exp. Neurol. 2018, 299, 217–227. [Google Scholar] [CrossRef]
- Chaliha, D.; Albrecht, M.; Vaccarezza, M.; Takechi, R.; Lam, V.; Al-Salami, H.; Mamo, J. A Systematic Review of the Valproic-Acid-Induced Rodent Model of Autism. Dev. Neurosci. 2020, 42, 12–48. [Google Scholar] [CrossRef]
- Bey, A.L.; Jiang, Y. Overview of Mouse Models of Autism Spectrum Disorders. Curr. Protoc. Pharmacol. 2014, 66, 5–66. [Google Scholar] [CrossRef] [Green Version]
- Matas, E.; Maisterrena, A.; Thabault, M.; Balado, E.; Francheteau, M.; Balbous, A.; Galvan, L.; Jaber, M. Major Motor and Gait Deficits with Sexual Dimorphism in a Shank3 Mutant Mouse Model. Mol. Autism 2021, 12, 2. [Google Scholar] [CrossRef]
- Thabault, M.; Turpin, V.; Maisterrena, A.; Jaber, M.; Egloff, M.; Galvan, L. Cerebellar and Striatal Implications in Autism Spectrum Disorders: From Clinical Observations to Animal Models. Int. J. Mol. Sci. 2022, 23, 2294. [Google Scholar] [CrossRef]
- Morriss-Kay, G.; Ruberte, E.; Fukiishi, Y. Mammalian Neural Crest and Neural Crest Derivatives. Ann. Anat.-Anat. Anz. 1993, 175, 501–507. [Google Scholar] [CrossRef]
- Solinas, M.; Thiriet, N.; El Rawas, R.; Lardeux, V.; Jaber, M. Environmental Enrichment during Early Stages of Life Reduces the Behavioral, Neurochemical, and Molecular Effects of Cocaine. Neuropsychopharmacology 2009, 34, 1102–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thye, M.D.; Bednarz, H.M.; Herringshaw, A.J.; Sartin, E.B.; Kana, R.K. The Impact of Atypical Sensory Processing on Social Impairments in Autism Spectrum Disorder. Dev. Cogn. Neurosci. 2018, 29, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Nishi, A.; Fisone, G.; Girault, J.-A.; Nairn, A.C.; Greengard, P. DARPP-32: An Integrator of Neurotransmission. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 269–296. [Google Scholar] [CrossRef] [PubMed]
- Andreska, T.; Lüningschrör, P.; Sendtner, M. Regulation of TrkB Cell Surface Expression—A Mechanism for Modulation of Neuronal Responsiveness to Brain-Derived Neurotrophic Factor. Cell Tissue Res. 2020, 382, 5–14. [Google Scholar] [CrossRef]
- Zikopoulos, B.; Barbas, H. Altered Neural Connectivity in Excitatory and Inhibitory Cortical Circuits in Autism. Front. Hum. Neurosci. 2013, 7, 609. [Google Scholar] [CrossRef] [Green Version]
- Filice, F.; Vörckel, K.J.; Sungur, A.Ö.; Wöhr, M.; Schwaller, B. Reduction in Parvalbumin Expression Not Loss of the Parvalbumin-Expressing GABA Interneuron Subpopulation in Genetic Parvalbumin and Shank Mouse Models of Autism. Mol. Brain 2016, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Jaber, M.; Robinson, S.W.; Missale, C.; Caron, M.G. Dopamine Receptors and Brain Function. Neuropharmacology 1996, 35, 1503–1519. [Google Scholar] [CrossRef]
- Jaber, M.; Jones, S.; Giros, B.; Caron, M.G. The Dopamine Transporter: A Crucial Component Regulating Dopamine Transmission. Mov. Disord. 1997, 12, 629–633. [Google Scholar] [CrossRef]
- Damasio, A.R.; Maurer, R.G. A Neurological Model for Childhood Autism. Arch. Neurol. 1978, 35, 777–786. [Google Scholar] [CrossRef]
- Langen, M.; Bos, D.; Noordermeer, S.D.S.; Nederveen, H.; van Engeland, H.; Durston, S. Changes in the Development of Striatum Are Involved in Repetitive Behavior in Autism. Biol. Psychiatry 2014, 76, 405–411. [Google Scholar] [CrossRef]
- Sears, L.L.; Vest, C.; Mohamed, S.; Bailey, J.; Ranson, B.J.; Piven, J. An MRI Study of the Basal Ganglia in Autism. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1999, 23, 613–624. [Google Scholar] [CrossRef]
- Langen, M.; Durston, S.; Staal, W.G.; Palmen, S.J.M.C.; van Engeland, H. Caudate Nucleus Is Enlarged in High-Functioning Medication-Naive Subjects with Autism. Biol. Psychiatry 2007, 62, 262–266. [Google Scholar] [CrossRef]
- McAlonan, G.M.; Cheung, V.; Cheung, C.; Suckling, J.; Lam, G.Y.; Tai, K.S.; Yip, L.; Murphy, D.G.M.; Chua, S.E. Mapping the Brain in Autism. A Voxel-Based MRI Study of Volumetric Differences and Intercorrelations in Autism. Brain 2005, 128, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Friedman, D.B.; Wang, Z.; Woodruff, E.; Pan, L.; O’donnell, J.; Broadie, K. Protein Expression Profiling of the Drosophila Fragile X Mutant Brain Reveals Up-Regulation of Monoamine Synthesis. Mol. Cell Proteom. 2005, 4, 278–290. [Google Scholar] [CrossRef] [Green Version]
- Campbell, N.G.; Shekar, A.; Aguilar, J.I.; Peng, D.; Navratna, V.; Yang, D.; Morley, A.N.; Duran, A.M.; Galli, G.; O’Grady, B.; et al. Structural, Functional, and Behavioral Insights of Dopamine Dysfunction Revealed by a Deletion in SLC6A3. Proc. Natl. Acad. Sci. USA 2019, 116, 3853–3862. [Google Scholar] [CrossRef] [Green Version]
- Mahabir, S.; Chatterjee, D.; Buske, C.; Gerlai, R. Maturation of Shoaling in Two Zebrafish Strains: A Behavioral and Neurochemical Analysis. Behav. Brain Res. 2013, 247, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Scerbina, T.; Chatterjee, D.; Gerlai, R. Dopamine Receptor Antagonism Disrupts Social Preference in Zebrafish: A Strain Comparison Study. Amino Acids 2012, 43, 2059–2072. [Google Scholar] [CrossRef]
- Panayotis, N.; Pratte, M.; Borges-Correia, A.; Ghata, A.; Villard, L.; Roux, J.-C. Morphological and Functional Alterations in the Substantia Nigra Pars Compacta of the Mecp2-Null Mouse. Neurobiol. Dis. 2011, 41, 385–397. [Google Scholar] [CrossRef]
- Shahbazian, M.; Young, J.; Yuva-Paylor, L.; Spencer, C.; Antalffy, B.; Noebels, J.; Armstrong, D.; Paylor, R.; Zoghbi, H. Mice with Truncated MeCP2 Recapitulate Many Rett Syndrome Features and Display Hyperacetylation of Histone H3. Neuron 2002, 35, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Rial, D.; Puighermanal, E.; Chazalon, M.; Valjent, E.; Schiffmann, S.N.; de Kerchove d’Exaerde, A. Mammalian Target of Rapamycin-RhoA Signaling Impairments in Direct Striatal Projection Neurons Induce Altered Behaviors and Striatal Physiology in Mice. Biol. Psychiatry 2020, 88, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Filice, F.; Janickova, L.; Henzi, T.; Bilella, A.; Schwaller, B. The Parvalbumin Hypothesis of Autism Spectrum Disorder. Front. Cell. Neurosci. 2020, 14, 577525. [Google Scholar] [CrossRef] [PubMed]
- He, J.L.; Oeltzschner, G.; Mikkelsen, M.; Deronda, A.; Harris, A.D.; Crocetti, D.; Wodka, E.L.; Mostofsky, S.H.; Edden, R.A.E.; Puts, N.A.J. Region-Specific Elevations of Glutamate + Glutamine Correlate with the Sensory Symptoms of Autism Spectrum Disorders. Transl. Psychiatry 2021, 11, 411. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Singel, D.; Hepburn, S.; Rojas, D.C. Increased Glutamate Concentration in the Auditory Cortex of Persons with Autism and First-Degree Relatives: A (1)H-MRS Study. Autism Res. 2013, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebartz van Elst, L.; Maier, S.; Fangmeier, T.; Endres, D.; Mueller, G.T.; Nickel, K.; Ebert, D.; Lange, T.; Hennig, J.; Biscaldi, M.; et al. Disturbed Cingulate Glutamate Metabolism in Adults with High-Functioning Autism Spectrum Disorder: Evidence in Support of the Excitatory/Inhibitory Imbalance Hypothesis. Mol. Psychiatry 2014, 19, 1314–1325. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, M.D. Synapse Structure: Glutamate Receptors Connected by the Shanks. Curr. Biol. 1999, 9, R848–R850. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maisterrena, A.; Matas, E.; Mirfendereski, H.; Balbous, A.; Marchand, S.; Jaber, M. The State of the Dopaminergic and Glutamatergic Systems in the Valproic Acid Mouse Model of Autism Spectrum Disorder. Biomolecules 2022, 12, 1691. https://doi.org/10.3390/biom12111691
Maisterrena A, Matas E, Mirfendereski H, Balbous A, Marchand S, Jaber M. The State of the Dopaminergic and Glutamatergic Systems in the Valproic Acid Mouse Model of Autism Spectrum Disorder. Biomolecules. 2022; 12(11):1691. https://doi.org/10.3390/biom12111691
Chicago/Turabian StyleMaisterrena, Alexandre, Emmanuel Matas, Helene Mirfendereski, Anais Balbous, Sandrine Marchand, and Mohamed Jaber. 2022. "The State of the Dopaminergic and Glutamatergic Systems in the Valproic Acid Mouse Model of Autism Spectrum Disorder" Biomolecules 12, no. 11: 1691. https://doi.org/10.3390/biom12111691
APA StyleMaisterrena, A., Matas, E., Mirfendereski, H., Balbous, A., Marchand, S., & Jaber, M. (2022). The State of the Dopaminergic and Glutamatergic Systems in the Valproic Acid Mouse Model of Autism Spectrum Disorder. Biomolecules, 12(11), 1691. https://doi.org/10.3390/biom12111691