
Biosynthesis and Biological Significances of LacdiNAc Group on N- and O-Glycans in Human Cancer Cells

Abstract
:1. Introduction
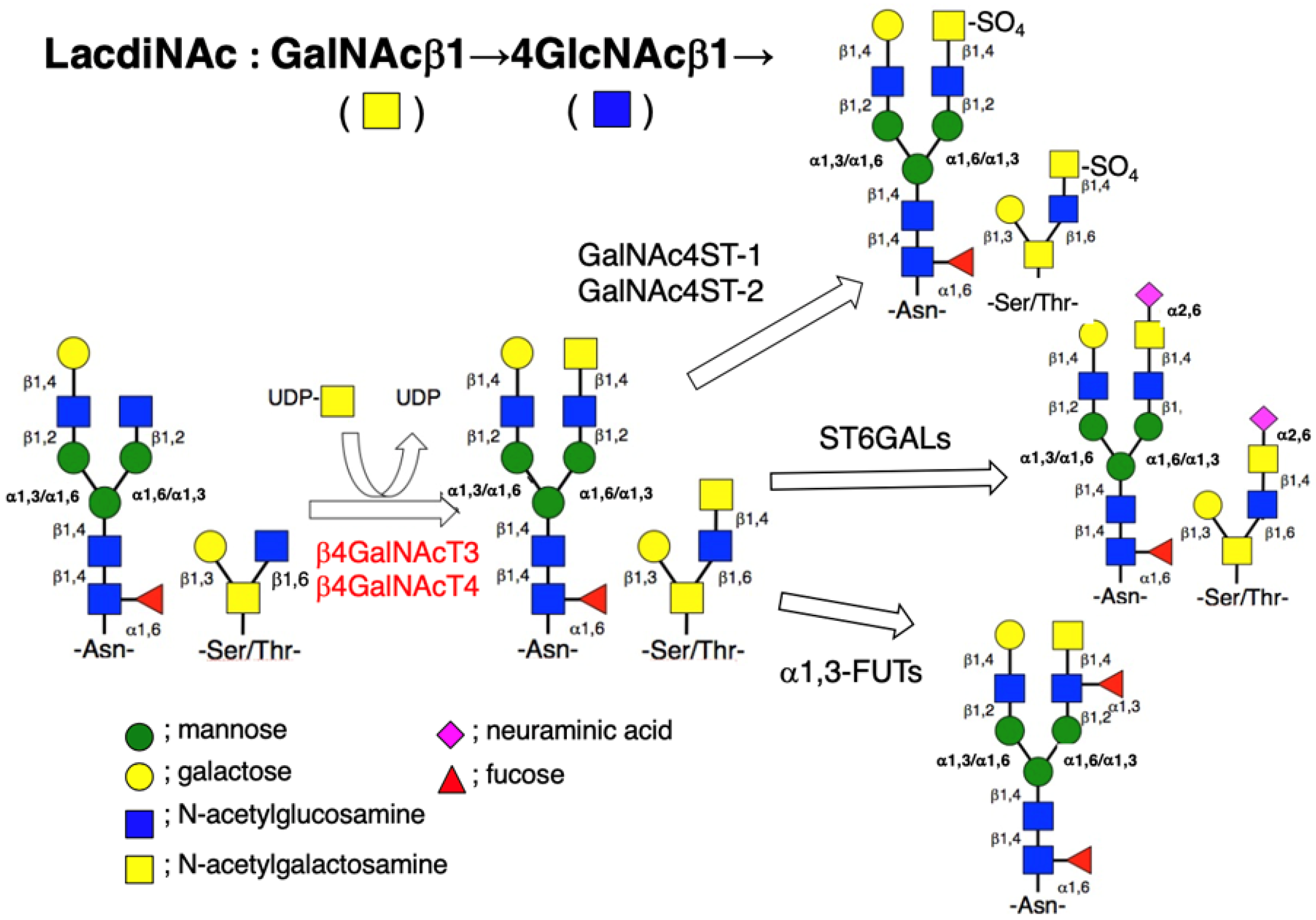
2. Biosynthesis of LacdiNAc Group and Its Modification in Mammalian Cells
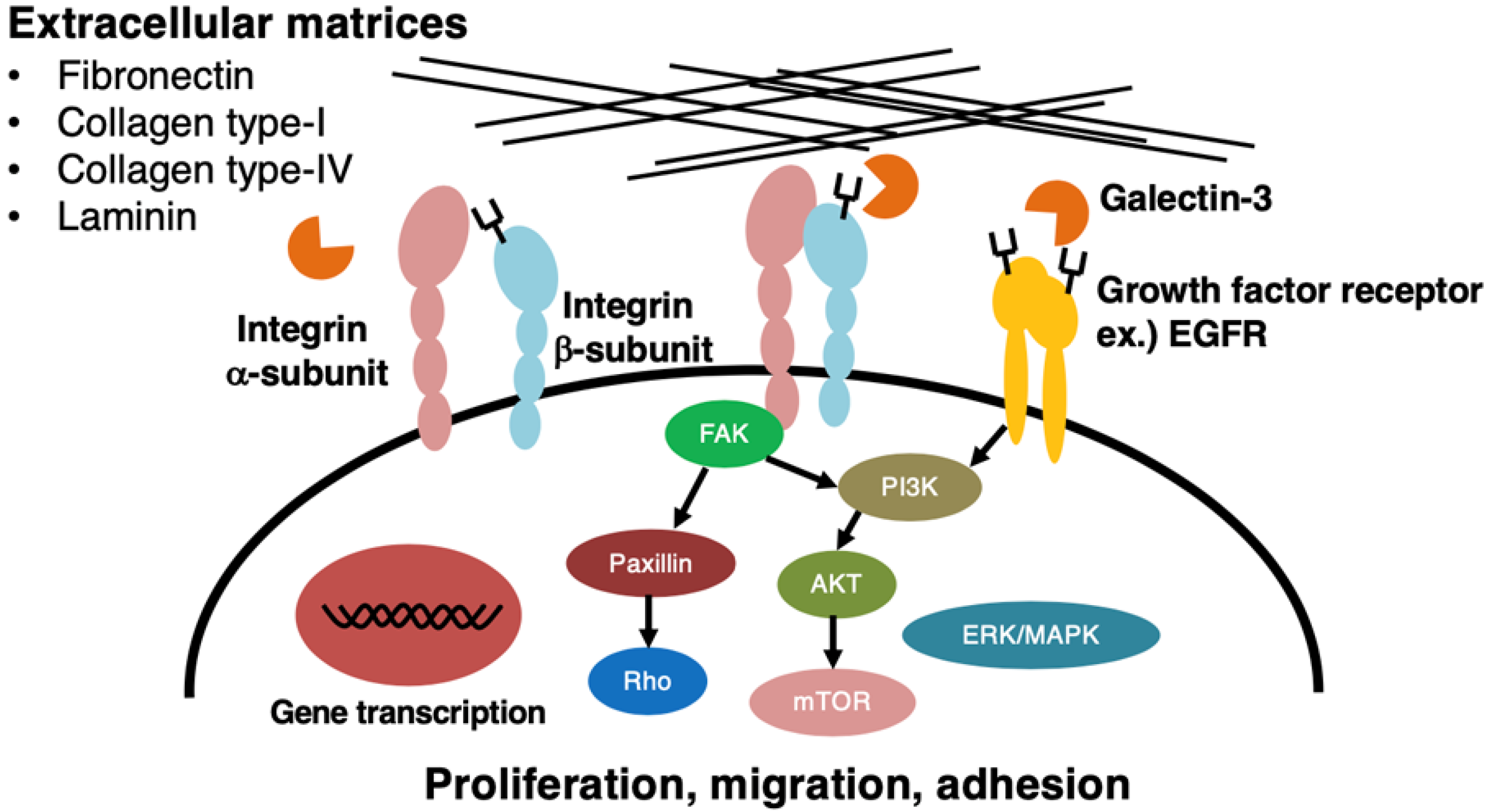
3. Biological Roles of LacdiNAc Group on N- and O-glycans
4. Differential Expression of LacdiNAc Group among Cancer Cells
5. Occurrence of LacdiNAc Group on N-glycans of Cell Surface Molecules
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varki, A. Biological roles of glycans. Glycobology 2017, 27, 3–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Die, I.; Cummings, R.D. Glycan gimmickry by parasitic helminths: A strategy for modulating the host immune response? Glycobiology 2010, 20, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Kitamura, N.; Sato, T.; Hiraizumi, S. Differentiation-associated expression of β-N-acetylgalactosaminylated N-linked oligosaccharides in mammary epithelial cells. Adv. Exp. Med. Biol. 2001, 491, 313–323. [Google Scholar] [PubMed]
- Hirano, K.; Matsuda, A.; Shirai, T.; Furukawa, K. Expression of LacdiNAc groups on N-glycans among human tumor is complex. BioMed Res. Int. 2014, 2014, 981627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Gotoh, M.; Kiyohara, K.; Kameyama, A.; Kubota, T.; Kikuchi, N.; Ishizuka, Y.; Iwasaki, H.; Togayachi, A.; Kubo, T.; et al. Molecular cloning and characterization of a novel human β1,4-N-acetylgalactosaminyltransferase, β4GalNAc-T3, responsible for the synthesis of N,N′-diacetyllactosediamine, GalNAcβ1-4GlcNAc. J. Biol. Chem. 2003, 278, 47534–47544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, M.; Sato, T.; Kiyohara, K.; Kameyama, A.; Kikuchi, N.; Kwon, Y.D.; Ishizuka, Y.; Nakanishi, H.; Narimatsu, H. Molecular cloning and characterization of β1,4-N-acetylgalactosaminyltransferases IV synthesizing N,N′-diacetyllactosediamine. FEBS Lett. 2004, 562, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Fiete, D.; Beranek, M.; Baenziger, J.U. Molecular basis for protein-specific transfer of N-acetylgalactosamine to N-linked glycans by the glycosyltransferases β1,4-N-acetylgalactosaminyl transferase 3 (β4GalNAc-T3) and β4GalNAc-T4. J. Biol. Chem. 2012, 287, 29194–29203. [Google Scholar] [CrossRef] [Green Version]
- Fiete, D.; Beranek, M.; Baenziger, J.U. Peptide-specific transfer of N-acetylgalactosamine to O-linked glycans by the glycosyltransferases β1,4-N-acetylgalactosaminyl transferase 3 (β4GalNAc-T3) and β4GalNAc-T4. J. Biol. Chem. 2012, 287, 29204–29212. [Google Scholar] [CrossRef] [Green Version]
- Ikehara, Y.; Sato, T.; Niwa, T.; Nakamura, S.; Gotoh, M.; Ikehara, S.K.; Kiyohara, K.; Aoki, C.; Iwai, T.; Nakanishi, H.; et al. Apical Golgi localization of N,N′-diacetyllactosediamine synthase, β4GalNAc-T3, is responsible for LacdiNAc expression on gastric mucosa. Glycobiology 2006, 16, 777–785. [Google Scholar] [CrossRef]
- Hirano, K.; Matsuda, A.; Kuji, R.; Nakandakari, S.; Shirai, T.; Furukawa, K. Enhanced expression of the β4-N-acetylgalactosaminyltransferase 4 gene impairs tumor growth of human breast cancer cells. Biochem. Biophys. Res. Commun. 2015, 461, 80–85. [Google Scholar] [CrossRef]
- Kawar, Z.S.; Haslam, S.M.; Morris, H.R.; Dell, A.; Cummings, R.D. Nobel poly-GalNAcβ1-4GlcNAc (LacdiNAc) and fucosylated poly-LacdiNAc N-glycans from mammalian cells expressing β1,4-N-acetylgalactosaminyltransferase and α1,3-fucosyltransferase. J. Biol. Chem. 2005, 280, 12810–12819. [Google Scholar] [CrossRef] [Green Version]
- Nakata, N.; Furukawa, K.; Greenwalt, D.E.; Sato, T.; Kibata, A. Structural study of the sugar chains of CD36 purified from bovine mammary epithelial cells: Occurrence of novel hybrid-type sugar chains containing the Neu5Acα2→6GalNAcβ1→4GlcNAc and the Manα1→2Manα1→3Manα1→6Man groups. Biochemistry 1993, 32, 4369–4383. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.B.; Chao, Y.B.; van Halbeek, H. Novel Asn-linked oligosaccharides terminating in GalNAcβ(1→4)[Fucα(1→3)]GlcNAcβ(1→) are present in recombinant human protein C expressed in human kidney 293 cells. Glycobiology 1993, 3, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Dell, A.; Morris, H.R.; Easton, R.L.; Panico, M.; Patankar, M.; Oehniger, S.; Koistinen, R.; Koistinen, H.; Seppala, M.; Clark, G.F. Structural analysis of the oligosaccharides derived from glycodelin, a human glycoproteins with potent immunosuppressive and contraceptive activities. J. Biol. Chem. 1995, 270, 24116–24126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, E.D.; van Halbeek, H.; Boime, I.; Baenziger, J.U. Structural elucidation of the disulfated oligosaccharide from bovine lutropin. J. Biol. Chem. 1985, 260, 15623–15630. [Google Scholar] [CrossRef]
- Fiete, D.; Srivastava, V.; Hindsgual, O.; Baenziger, J.U. A hepatic reticuloendothelial cell receptor specific for SO4-4GalNAcβ1,4GlcNAcβ1,2Manα that mediates rapid clearance of lutropin. Cell 1991, 67, 1103–1110. [Google Scholar] [CrossRef]
- Yu, S.Y.; Chang, L.Y.; Cheng, C.W.; Chou, C.C.; Fukuda, M.N.; Khoo, K.H. Priming mass spectrometry-based sulfoglycomic mapping for identification of terminal sulfated LacdiNAc glycotope. Glycoconj. J. 2013, 30, 183–194. [Google Scholar] [CrossRef]
- Benicky, J.; Sanda, M.; Brnakova Kennedy, Z.; Goldman, R. N-Glycosylation is required for secretion of the precursor to brain-derived neurotrophic factor (proBDNF) carrying sulfated LacdiNAc structures. J. Biol. Chem. 2019, 294, 16816–16830. [Google Scholar] [CrossRef]
- Xia, G.; Evers, M.R.; Kang, H.G.; Schachner, M.; Baenziger, J.U. Molecular cloning and expression of the pituitary glycoprotein N-acetylglucosamine-4-O-sulfotransferase. J. Biol. Chem. 2000, 275, 38402–38409. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, N.; Misra, A.; Belot, F.; Hindsgaul, O.; Fukuda, M. Molecular cloning and expression of the two distinct human N-acetylglucosamine-4-O-sulfotransferases that transfer sulfate to GalNAcβ1→4GlcNAcβ1-R in both N- and O-glycans. Glycobiology 2001, 11, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Gatermann, M.; Nimtz, M.; Kandzia, S.; Glatzel, M.; Conradt, H.S. N-glycosylation of Extracellular Vesicles from HEK-293 and Glioma Cell Lines. Anal. Chem. 2018, 90, 7871–7879. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, A.; Schnaar, R.L. Siglec regands. Cells 2021, 10, 1260. [Google Scholar] [CrossRef] [PubMed]
- Fiete, D.J.; Beranek, M.C.; Baenziger, J.U. A cysteine-rich domain of the “mannose” receptor mediates GalNAc-4-SO4 binding. Proc. Natl. Acad. Sci. USA 1998, 95, 2089–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.I.; Manzella, S.M.; Baenziger, J.U. Rapid clearance of sialylated glycoproteins by the asialoglycoprotein receptor. J. Biol. Chem. 2003, 278, 4597–4602. [Google Scholar] [CrossRef] [Green Version]
- Böhm, E.; Seyfried, B.K.; Dockal, M.; Graninger, M.; Hasslacher, M.; Neurath, M.; Konetschny, C.; Matthiessen, P.; Mitterer, A.; Scheiflinger, F. Differences in N-glycosylation of recombinant human coagulation factor VII derived from BHK, CHO, and HEK293 cells. BMC Biotechnol. 2015, 15, 87. [Google Scholar] [CrossRef] [Green Version]
- Uhler, R.; Popa-Wagner, R.; Kröning, M.; Brehm, A.; Rennert, P.; Seifried, A.; Peschke, M.; Krieger, M.; Kohla, G.; Kannicht, C.; et al. Glyco-engineered HEK 293-F cell lines for the production of therapeutic glycoproteins with human N-glycosyaltion and improved pharmacokinetics. Glycobiology 2021, 37, 859–872. [Google Scholar] [CrossRef]
- Tsukimura, W.; Kurogochi, M.; Mori, M.; Osumi, K.; Matsuda, A.; Takegawa, K.; Furukawa, K.; Shirai, T. Preparation and biological activities of anti-HER2 monoclonal antibodies with fully core-fucosylated homogeneous bi-antennary complex-type glycans. Biosci. Biotechnol. Biochem. 2017, 81, 2353–2359. [Google Scholar] [CrossRef] [Green Version]
- Giddens, J.P.; Lomino, J.V.; DiLillo, D.J.; Ravetch, J.V.; Wang, L.X. Site-selective chemoenzymatic glycoengineering of Fab and Fc glycans of a therapeutic antibody. Proc. Natl. Acad. Sci. USA 2018, 115, 12023–12027. [Google Scholar] [CrossRef] [Green Version]
- Kenny, D.T.; Skoog, E.C.; Lindén, S.K.; Struwe, W.B.; Rudd, P.M.; Karlsson, N.G. The presence of terminal N-acetylgalactosamine β1-4N-acetylglucosamine residues on O-linked oligosaccharides from gastric MUC5AC: Involvement in Helicobacter pylori colocalization? Glycobiology 2012, 22, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Matos, R.; Amorim, I.; Magalhães, A.; Haesebrouck, F.; Gärtner, F.; Reis, C.A. Adhesion of Helicobacter species to the human gastric mucosa: A deep look into glycans role. Front. Mol. Biosci. 2021, 8, 656439. [Google Scholar] [CrossRef]
- van den Berg, T.K.; Honing, H.; Franke, N.; van Remoortere, A.; Schiphorst, W.E.; Liu, F.T.; Deelder, A.M.; Cummings, R.D.; Hokke, C.H.; van Die, I. LacdiNAc-glycans constitute a parasitic pattern for galectin-3-mediated immune recognition. J. Immunol. 2004, 173, 1902–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böcker, S.; Elling, L. Biotinylated N-Acetyllactosamine- and N,N-Diacetyllactosamine-Based Oligosaccharides as Novel Ligands for Human Galectin-3. Bioengineering 2017, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, C.D.L.; Coelho, H.; Gimeno, A.; Trovão, F.; Diniz, A.; Dias, J.S.; Jiménez-Barbero, J.; Corzana, F.; Carvalho, A.L.; Cabrita, E.J.; et al. Structural insights of molecular recognition mechanism of cancer and pathogenic epitope LacdiNAc, by immune-related lectins. Chemistry 2021, 27, 7951–7958. [Google Scholar] [CrossRef] [PubMed]
- Sedlář, A.; Trávníčková, M.; Bojarová, P.; Vlachová, M.; Slámová, K.; Křen, V.; Bačáková, L. Interaction between galectin-3 and integrins mediates cell-matrix adhesion in endothelial cells and mesenchymal stem cells. Int. J. Mol. Sci. 2021, 22, 5144. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Dong, X.W.; Guo, X.L. Role of the interaction between galectin-3 and cell adhesion molecules in cancer metastasis. Biomed. Pharmacother. 2015, 69, 179–185. [Google Scholar] [CrossRef]
- Wang, L.; Guo, X.L. Molecular regulation of galectin-3 expression and therapeutic implication in cancer progression. Biomed. Pharmacother. 2016, 78, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laaf, D.; Bojarová, P.; Elling, L.; Křen, V. Galectin-carbohydrate interactions in biomedicine and biotechnology. Trends. Biotechnol. 2019, 37, 402–415. [Google Scholar] [CrossRef]
- Sato, T.; Taka, J.; Aoki, N.; Matsuda, T.; Furukawa, K. Expression of β-N-acetylgalactosaminylated N-linked sugar chains is associated with functional differentiation of bovine mammary gland. J. Biochem. 1997, 122, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, N.; Shinomi, M.; Hirano, K.; Ui-Tei, K.; Nishihara, S. LacdiNAc (GalNAcβ1-4GlcNAc) contributes to self-renewal of mouse embryonic stem cells by regulating leukemia inhibitory factor/STAT3 signaling. Stem Cell 2011, 29, 641–650. [Google Scholar] [CrossRef]
- Fukushima, K.; Satoh, T.; Baba, S.; Yamashita, K. α1,2-Fucosylated and β-N-acetylgalactosaminylated prostate-specific antigen as an efficient marker of prostatic cancer. Glycobiology 2010, 20, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Gratacós-Mulleras, A.; Duran, A.; Shehni, A.A.; Ferrer-Batallé, M.; Ramírez, M.; Comet, J.; de Llorens, R.; Saldova, R.; Llop, E.; Peracaula, R. Characterisation of the main PSA glycoforms in aggressive prostate cancer. Sci. Rep. 2020, 10, 18974. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, K.; Tobisawa, Y.; Kaya, T.; Kaneko, T.; Hatakeyama, S.; Mori, K.; Hashimoto, Y.; Koie, T.; Suda, Y.; Ohyama, C.; et al. Wisteria floribunda Agglutinin and its reactive-glycan carrying prostate specific antigen as a novel diagnostic and prognostic marker of prostate cancer. Int. J. Mol. Sci. 2017, 18, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, T.; Kaneko, T.; Kojima, S.; Nakamura, Y.; Ide, Y.; Ishida, K.; Suda, Y.; Yamashita, K. High-sensitivity immunoassay with surface plasmon field-enhanced fluorescence spectroscopy using a plastic sensor chip: Application to quantitative analysis of total prostate-specific antigen and GalNAcβ1-4GlcNAc-linked prostate-specific antigen for prostate cancer diagnosis. Anal. Chem. 2015, 87, 1797–1803. [Google Scholar]
- Haga, Y.; Uemura, M.; Baba, S.; Inamura, K.; Takeuchi, K.; Nonomura, N.; Ueda, K. Identification of multisiaylated LacdiNAc structures as highly prostate cancer specific glycan signatures on PSA. Anal. Chem. 2019, 91, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liang, J.T.; Huang, H.C.; Shen, T.L.; Chen, H.Y.; Lin, N.Y.; Che, M.I.; Lin, W.C.; Huang, M.C. β1,4-N-acetylgalactosaminyltransferase III enhances malignant phenotypes of colon cancer cells. Mol. Cancer Res. 2007, 5, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Peracaula, R.; Royle, L.; Tabarés, G.; Mallorqui-Fernandez, G.; Barrabés, S.; Harvey, D.J.; Dwek, R.A.; Rudd, P.M.; Llorens, R. Glycosylation of human pancreatic ribonuclease: Differences between normal and tumor states. Glycobiology 2003, 13, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Sogabe, M.; Nozaki, H.; Tanaka, N.; Kubota, T.; Kaji, H.; Kuno, A.; Togayachi, A.; Gotoh, M.; Nakanishi, H.; Nakanishi, T.; et al. Nobel glycobiomarker for ovarian cancer that detects clear cell carcinoma. J. Proteome Res. 2014, 13, 1624–1635. [Google Scholar] [CrossRef]
- Anugraham, M.; Jacob, F.; Everest-Dass, A.V.; Schoetzau, A.; Nixdorf, S.; Hacker, N.F.; Fink, D.; Heinzelmann-Schwarz, V.; Packer, N.H. Tissue glycomics distinguish tumour sites in women with advanced serous adenocarcinoma. Mol. Oncol. 2017, 11, 1595–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, W.M.; Che, M.I.; Liao, Y.F.; Chang, H.H.; Chen, C.H.; Huang, Y.M.; Jeng, Y.M.; Huang, J.; Quon, M.J.; Lee, H.; et al. B4GALNT3 Expression Predicts a Favorable Prognosis and Suppresses Cell Migration and Invasion via β1-Integrin Signaling in Neuroblastoma. Am. J. Pathol. 2011, 179, 1394–1404. [Google Scholar] [CrossRef]
- Kitamura, N.; Guo, S.; Sato, T.; Hiraizumi, S.; Taka, J.; Ikekita, M.; Sawada, S.; Fujisawa, H.; Furukawa, K. Prognostic significance of reduced expression of β-N-acetylgalactosaminylated N-linked oligosaccharides in human breast cancer. Int. J. Cancer 2003, 105, 533–541. [Google Scholar] [CrossRef]
- Jin, C.; Kenny, D.T.; Skoog, E.C.; Padra, M.; Adamczyk, B.; Vitizeva, V.; Thorell, A.; Venkatakrishnan, V.; Lindén, S.K.; Karlsson, N.G. Structural diversity of human gastric mucin glycans. Mol. Cell. Proteom. 2017, 16, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.; Jin, C.; Polom, K.; Muñoz, P.; Rojas-Macias, M.A.; Zeeberg, D.; Borén, M.; Roviello, F.; Karlsson, N.G. Sample handling of gastric tissue and O-glycan alterations in paired gastric cancer and non-tumorigenic tissues. Sci. Rep. 2018, 8, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, K.; Takada, Y.; Furukawa, K. LacdiNAcylation of N-glycans in MDA-MB-231 human breast cancer cells results in changes in morphological appearance and adhesive properties of the cells. Histochem. Cell Biol. 2020, 153, 17–26. [Google Scholar] [CrossRef]
- Che, M.I.; Huang, J.; Hung, J.S.; Lin, Y.C.; Huang, M.J.; Lai, H.S.; Hsu, W.M.; Liang, J.T.; Huang, M.C. β1,4-N-acetylgalactosaminyltransferase III modulates cancer stemness through EGFR signaling pathway in colon cancer cells. Oncotarget 2014, 15, 3673–3684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Isaji, T.; Im, S.; Fukuda, T.; Hashii, N.; Takakura, D.; Kawasaki, N.; Gu, J. β-Galactoside α2,6-sialyltranferase 1 promotes transforming growth factor-β-mediated epithelial-mesenchymal transition. J. Biol. Chem. 2014, 289, 34627–34641. [Google Scholar] [CrossRef] [Green Version]
- Kadry, Y.A.; Calderwood, D.A. Structural and signaling functions of integrins. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183206. [Google Scholar] [CrossRef]
- Isaji, T.; Sato, Y.; Zhao, Y.; Miyoshi, E.; Wada, Y.; Taniguchi, N.; Gu, J. N-glycosylation of the β-propeller domain of the integrin α5 subunit is essential for α5β1 heterodimerization, expression on the cell surface, and its biological function. J. Biol. Chem. 2006, 281, 33258–33267. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Isaji, T.; Xu, Q.; Kariya, Y.; Gu, W.; Fukuda, T.; Du, Y. Potential roles of N-glycosylation in cell adhesion. Glycoconj. J. 2012, 29, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Kizuka, Y.; Ohtsubo, K.; Gu, J.; Taniguchi, N. Desease-associated glycans on cell surface proteins. Mol. Asp. Med. 2016, 51, 56–70. [Google Scholar] [CrossRef]
- Isaji, T.; Gu, J.; Nishiuchi, R.; Zhao, Y.; Takahashi, M.; Miyoshi, E.; Honke, K.; Sekiguchi, K.; Taniguchi, N. Introduction of bisecting GlcNAc into integrin α5β1 reduces ligand binding and down-regulates cell adhesion and cell migration. J. Biol. Chem. 2004, 279, 19747–19754. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Isaji, T.; Tajiri, M.; Yoshida-Yamamoto, S.; Yoshinaka, T.; Somehara, T.; Fukuda, T.; Wada, Y.; Gu, J. An N-glycosylation site on the β-propeller domain of the integrin a5 subunit plays key roles in both its function and site-specific modification by β1,4-N-acetylglucosaminyltransferase III. J. Biol. Chem. 2009, 284, 11873–11881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matarrese, P.; Fusco, O.; Tinari, N.; Natoli, C.; Liu, F.T.; Semeraro, M.L.; Malorni, W.; Iacobelli, S. Galectin-3 overexpression protects from apoptosis by improving cell adhesion properties. Int. J. Cancer. 2000, 85, 545–554. [Google Scholar] [CrossRef]
- Takenaka, Y.; Fukumori, T.; Raz, A. Galectin-3 and metastasis. Glycoconj. J. 2002, 19, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K. Changes in gene expression of β-1,4-galactosyltransferases by malignant transformation. Int. Congr. Ser. 2001, 443, 221–229. [Google Scholar] [CrossRef]
- Tagawa, M.; Shirane, K.; Yu, L.; Sato, T.; Furukawa, S.; Mizuguchi, H.; Kuji, R.; Kawamura, K.; Takahashi, N.; Kato, K.; et al. Enhanced expression of β4-galactosyltransferase 2 gene impairs mammalian tumor growth. Cancer Gene Ther. 2014, 21, 219–227. [Google Scholar] [CrossRef]
- Shirane, K.; Kuji, R.; Tareyanagi, C.; Sato, T.; Kobayashi, Y.; Furukawa, S.; Murata, T.; Kubota, S.; Ishikawa, Y.; Segawa, K.; et al. Gene expression levels of β4-galactosyltransferase 5 correlate with the tumorigenic potentials of B16-F10 mouse melanoma cells. Glycobiology 2014, 24, 532–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Types of Tumor/Cancer Cells | References |
|---|---|
| Positive correlation | |
| Prostate cancer cells | [40,41,42,43,44] |
| Colon cancer cells | [45] |
| Pancreatic cancer cells | [46] |
| Ovarian cancer tissues and cells | [47,48] |
| Negative correlation | |
| Neuroblastoma tissues and cells | [49] |
| Breast cancer tissues and cells | [50] |
| Gastric cancer tissues and cells | [51,52] |
| Malignant Properties | References |
|---|---|
| Proliferation | |
| promoted in HCT116 colon cancer cells | [45] |
| suppressed in SK-N-SH/SH-SY5Y neuroblastoma cells | [49] |
| suppressed in MDA-MB-231 breast cancer cells | [10] |
| Migration and invasion | |
| promoted in HCT116 colon cancer cells | [45] |
| suppressed in SK-N-SH/SH-SY5Y neuroblastoma cells | [49] |
| suppressed in MDA-MB-231 breast cancer cells | [10] |
| Adhesion to extracellular matrices | |
| promoted in HCT116 colon cancer cells | [45] |
| suppressed in SK-N-SH/SH-SY5Y neuroblastoma cells | [49] |
| promoted in MDA-MB-231 breast cancer cells | [53] |
| Mesenchymal-epithelial transition | |
| induced in MDA-MB-231 breast cancer cells | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirano, K.; Furukawa, K. Biosynthesis and Biological Significances of LacdiNAc Group on N- and O-Glycans in Human Cancer Cells. Biomolecules 2022, 12, 195. https://doi.org/10.3390/biom12020195
Hirano K, Furukawa K. Biosynthesis and Biological Significances of LacdiNAc Group on N- and O-Glycans in Human Cancer Cells. Biomolecules. 2022; 12(2):195. https://doi.org/10.3390/biom12020195
Chicago/Turabian StyleHirano, Kiyoko, and Kiyoshi Furukawa. 2022. "Biosynthesis and Biological Significances of LacdiNAc Group on N- and O-Glycans in Human Cancer Cells" Biomolecules 12, no. 2: 195. https://doi.org/10.3390/biom12020195
APA StyleHirano, K., & Furukawa, K. (2022). Biosynthesis and Biological Significances of LacdiNAc Group on N- and O-Glycans in Human Cancer Cells. Biomolecules, 12(2), 195. https://doi.org/10.3390/biom12020195