
Neuronal GHS-R Differentially Modulates Feeding Patterns under Normal and Obesogenic Conditions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Meal Pattern
2.3. Real-Time PCR
2.4. Statistical Analysis
3. Results
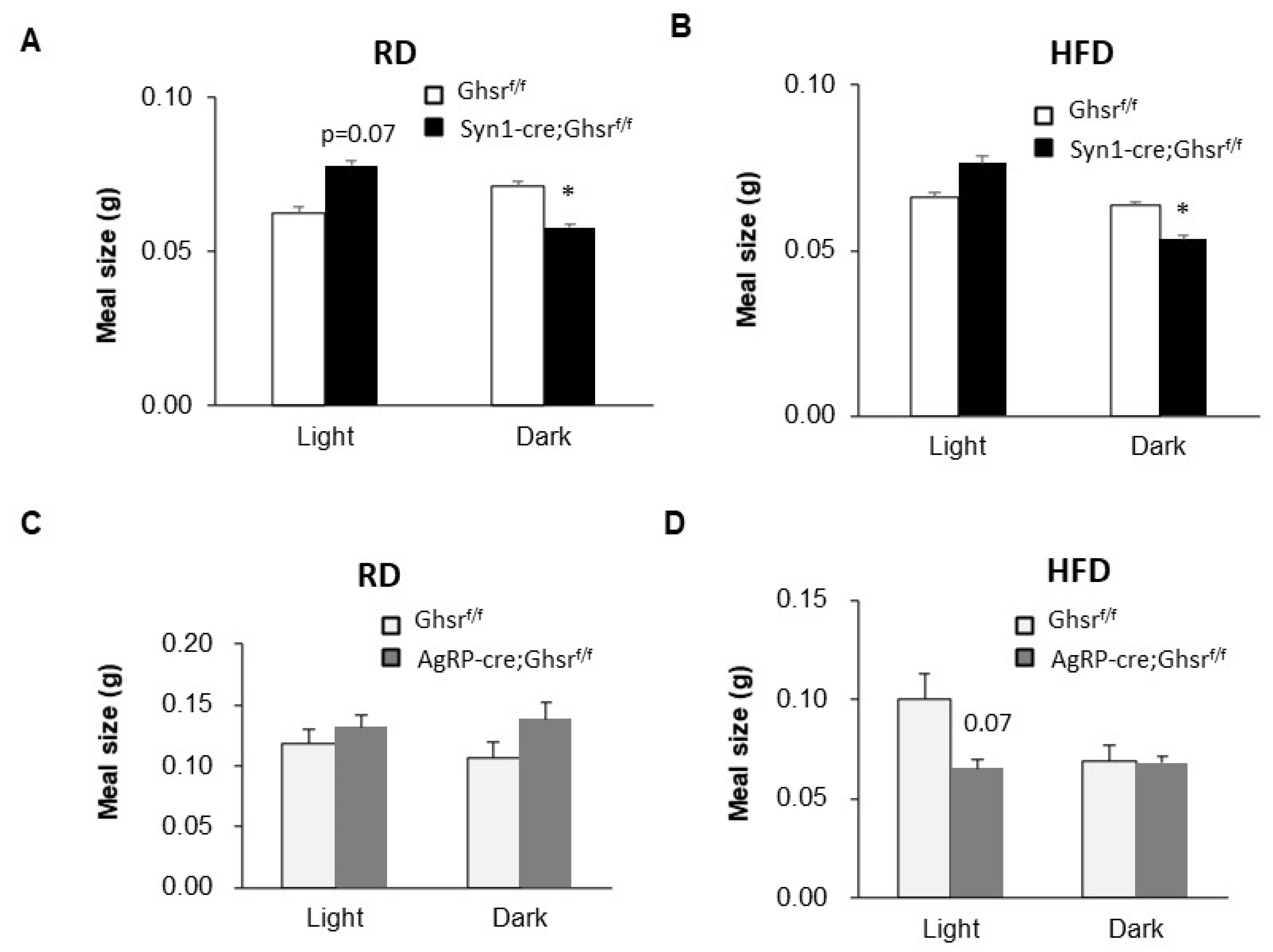
3.1. Meal Number
3.2. Meal Size
3.3. Meal Duration
3.4. Neuropeptide Changes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, G.; Kim, K.; Wilding, J.; World Obesity Federation. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Bertone, E.R.; Stanek, E.J.; Reed, G.W.; Hebert, J.R.; Cohen, N.L.; Merriam, P.A.; Ockene, I.S. Association between Eating Patterns and Obesity in a Free-living US Adult Population. Am. J. Epidemiol. 2003, 158, 85–92. [Google Scholar] [CrossRef] [PubMed]
- McCrory, M.A.; Howarth, N.C.; Roberts, S.B.; Huang, T.T.-K. Eating Frequency and Energy Regulation in Free-Living Adults Consuming Self-Selected Diets. J. Nutr. 2010, 141, 148–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodrick, C.; Ingram, D.; Reynolds, M.; Freeman, J.; Cider, N. Effects of intermittent feeding upon body weight and lifespan in inbred mice: Interaction of genotype and age. Mech. Ageing Dev. 1990, 55, 69–87. [Google Scholar] [CrossRef]
- Mattson, M.P.; Duan, W.; Guo, Z. Meal size and frequency affect neuronal plasticity and vulnerability to disease: Cellular and molecular mechanisms. J. Neurochem. 2003, 84, 417–431. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 4679–4684. [Google Scholar] [CrossRef] [Green Version]
- Tschop, M.H.; Smiley, D.L.; Heiman, M.L. Ghrelin induces adiposity in rodents. Nature 2000, 407, 908–913. [Google Scholar] [CrossRef]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E.; Weigle, D.S. A Preprandial Rise in Plasma Ghrelin Levels Suggests a Role in Meal Initiation in Humans. Diabetes 2001, 50, 1714–1719. [Google Scholar] [CrossRef] [Green Version]
- Labarthe, A.; Zizzari, P.; Fiquet, O.; Lebrun, N.; Veldhuis, J.D.; Roelfsema, F.; Chauveau, C.; Bohlooly-Y, M.; Epelbaum, J.; Tolle, V. Effect of GHS-R deletion on growth, pulsatile GH secretion and meal pattern in male and female mice. Neuroendocrinology 2021. [Google Scholar] [CrossRef]
- Sun, Y.; Ahmed, S.; Smith, R.G. Deletion of Ghrelin Impairs neither Growth nor Appetite. Mol. Cell. Biol. 2003, 23, 7973–7981. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Lin, L.; Qin, G.; Lu, X.; Fiorotto, M.; Dixit, V.D.; Sun, Y. Ablations of Ghrelin and Ghrelin Receptor Exhibit Differential Metabolic Phenotypes and Thermogenic Capacity during Aging. PLoS ONE 2011, 6, e16391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfluger, P.T.; Kirchner, H.; Günnel, S.; Schrott, B.; Perez-Tilve, D.; Fu, S.; Benoit, S.C.; Horváth, T.; Joost, H.-G.; Wortley, K.E.; et al. Simultaneous deletion of ghrelin and its receptor increases motor activity and energy expenditure. Am. J. Physiol. Liver Physiol. 2008, 294, G610–G618. [Google Scholar] [CrossRef]
- Faulconbridge, L.F.; Cummings, D.E.; Kaplan, J.M.; Grill, H.J. Hyperphagic Effects of Brainstem Ghrelin Administration. Diabetes 2003, 52, 2260–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Nuotio-Antar, A.M.; Ma, X.; Liu, F.; Fiorotto, M.L.; Sun, Y. Ghrelin Receptor Regulates Appetite and Satiety during Aging in Mice by Regulating Meal Frequency and Portion Size but Not Total Food Intake. J. Nutr. 2014, 144, 1349–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Lin, L.; Xu, P.; Saito, K.; Wei, Q.; Meadows, A.G.; Bongmba, O.Y.N.; Pradhan, G.; Zheng, H.; Xu, Y.; et al. Neuronal Deletion of Ghrelin Receptor Almost Completely Prevents Diet-Induced Obesity. Diabetes 2016, 65, 2169–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-S.; Bongmba, O.Y.N.; Yue, J.; Lee, J.H.; Lin, L.; Saito, K.; Pradhan, G.; Li, D.-P.; Pan, H.-L.; Xu, A.; et al. Suppression of GHS-R in AgRP Neurons Mitigates Diet-Induced Obesity by Activating Thermogenesis. Int. J. Mol. Sci. 2017, 18, 832. [Google Scholar] [CrossRef] [Green Version]
- Timper, K.; Brüning, J.C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model. Mech. 2017, 10, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.T.; Kola, B.; Feltrin, D.; Perez-Tilve, D.; Tschöp, M.H.; Grossman, A.B.; Korbonits, M. Ghrelin and cannabinoids require the ghrelin receptor to affect cellular energy metabolism. Mol. Cell. Endocrinol. 2013, 365, 303–308. [Google Scholar] [CrossRef]
- Sohn, J.-W. Network of hypothalamic neurons that control appetite. BMB Rep. 2015, 48, 229–233. [Google Scholar] [CrossRef]
- Sindelar, D.K.; Mystkowski, P.; Marsh, D.J.; Palmiter, R.D.; Schwartz, M.W. Attenuation of Diabetic Hyperphagia in Neuropeptide Y–Deficient Mice. Diabetes 2002, 51, 778–783. [Google Scholar] [CrossRef] [Green Version]
- Serrenho, D.; Santos, S.; Carvalho, A.L. The Role of Ghrelin in Regulating Synaptic Function and Plasticity of Feeding-Associated Circuits. Front. Cell. Neurosci. 2019, 13, 205. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, M.; Murakami, N.; Date, Y.; Kojima, M.; Matsuo, H.; Kangawa, K.; Matsukura, S. A role for ghrelin in the central regulation of feeding. Nature 2001, 409, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Kohno, D.; Gao, H.-Z.; Muroya, S.; Kikuyama, S.; Yada, T. Ghrelin Directly Interacts With Neuropeptide-Y-Containing Neurons in the Rat Arcuate Nucleus. Diabetes 2003, 52, 948–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takase, K.; Tsuneoka, Y.; Oda, S.; Kuroda, M.; Funato, H. High-fat diet feeding alters olfactory-, social-, and reward-related behaviors of mice independent of obesity. Obesity 2016, 24, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Beutler, L.R.; Corpuz, T.V.; Ahn, J.S.; Kosar, S.; Song, W.; Chen, Y.; A Knight, Z. Obesity causes selective and long-lasting desensitization of AgRP neurons to dietary fat. eLife 2020, 9, 55909. [Google Scholar] [CrossRef]
- Cardinal, P.; André, C.; Quarta, C.; Bellocchio, L.; Clark, S.; Elie, M.; Leste-Lasserre, T.; Maitre, M.; Gonzales, D.; Cannich, A.; et al. CB1 cannabinoid receptor in SF1-expressing neurons of the ventromedial hypothalamus determines metabolic responses to diet and leptin. Mol. Metab. 2014, 3, 705–716. [Google Scholar] [CrossRef]
- Fabelo, C.; Hernandez, J.; Chang, R.; Seng, S.; Alicea, N.; Tian, S.; Conde, K.; Wagner, E.J. Endocannabinoid Signaling at Hypothalamic Steroidogenic Factor-1/Proopiomelanocortin Synapses Is Sex- and Diet-Sensitive. Front. Mol. Neurosci. 2018, 11, 214. [Google Scholar] [CrossRef]
- Soria-Gomez, E.; Bellocchio, L.; Reguero, L.; Lepousez, G.; Martin, C.; Bendahmane, M.; Ruehle, S.; Remmers, F.; Desprez, T.; Matias, I.; et al. The endocannabinoid system controls food intake via olfactory processes. Nat. Neurosci. 2014, 17, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Dias-Rocha, C.P.; Almeida, M.M.; Santana, E.M.; Costa, J.C.; Franco, J.G.; Pazos-Moura, C.C.; Trevenzoli, I.H. Maternal high-fat diet induces sex-specific endocannabinoid system changes in newborn rats and programs adiposity, energy expenditure and food preference in adulthood. J. Nutr. Biochem. 2018, 51, 56–68. [Google Scholar] [CrossRef]
- Drummond, S.; Crombie, N.; Cursiter, M.; Kirk, T. Evidence that eating frequency is inversely related to body weight status in male, but not female, non-obese adults reporting valid dietary intakes. Int. J. Obes. 1998, 22, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Leidy, H.J.; Campbell, W.W. The Effect of Eating Frequency on Appetite Control and Food Intake: Brief Synopsis of Controlled Feeding Studies. J. Nutr. 2010, 141, 154–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anson, R.; Guo, Z.; de Cabo, R.; Iyun, T.; Rios, M.; Hagepanos, A.; Ingram, D.K.; Lane, M.A.; Mattson, M.P. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc. Natl. Acad. Sci. USA 2003, 100, 6216–6220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Slattery, J.A.; Milte, C.; Laker, R.; O’Donnell, T.; Dorian, C.; Brierley, S.M.; Blackshaw, L.A. Ghrelin selectively reduces mechanosensitivity of upper gastrointestinal vagal afferents. Am. J. Physiol. Liver Physiol. 2007, 292, G1376–G1384. [Google Scholar] [CrossRef] [Green Version]
- Ziotopoulou, M.; Mantzoros, C.S.; Hileman, S.M.; Flier, J.S. Differential expression of hypothalamic neuropeptides in the early phase of diet-induced obesity in mice. Am. J. Physiol. Metab. 2000, 279, E838–E845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E La Fleur, S.; Van Rozen, A.J.; Luijendijk, M.C.M.; Groeneweg, F.; Adan, R.A.H. A free-choice high-fat high-sugar diet induces changes in arcuate neuropeptide expression that support hyperphagia. Int. J. Obes. 2009, 34, 537–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, L.H.; Hurd, Y.L. Endocannabinoid signalling in reward and addiction. Nat. Rev. Neurosci. 2015, 16, 579–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, M. Cannabinoid Receptor Signaling in Central Regulation of Feeding Behavior: A Mini-Review. Front. Neurosci. 2017, 11, 293. [Google Scholar] [CrossRef]
- Castillo, P.E.; Younts, T.J.; Chávez, A.E.; Hashimotodani, Y. Endocannabinoid Signaling and Synaptic Function. Neuron 2012, 76, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Robbe, D.; Alonso, G.; Duchamp, F.; Bockaert, J.; Manzoni, O. Localization and Mechanisms of Action of Cannabinoid Receptors at the Glutamatergic Synapses of the Mouse Nucleus Accumbens. J. Neurosci. 2001, 21, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; He, Y.; Bi, G.-H.; Zhang, H.-Y.; Song, R.; Liu, Q.-R.; Egan, J.M.; Gardner, E.L.; Li, J.; Xi, Z.-X. CB1 Receptor Activation on VgluT2-Expressing Glutamatergic Neurons Underlies Δ9-Tetrahydrocannabinol (Δ9-THC)-Induced Aversive Effects in Mice. Sci. Rep. 2017, 7, 12315. [Google Scholar] [CrossRef] [Green Version]
- Kola, B. Role of AMP-Activated Protein Kinase in the Control of Appetite. J. Neuroendocr. 2008, 20, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-S.; Bongmba, O.Y.N.; Lee, J.H.; Tuchaai, E.; Zhou, Y.; Li, D.-P.; Xue, B.; Chen, Z.; Sun, Y. Ghrelin receptor in agouti-related peptide neurones regulates metabolic adaptation to calorie restriction. J. Neuroendocr. 2019, 31, e12763. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Xue, B.; Chen, Z.; Sun, Y. Neuronal GHS-R Differentially Modulates Feeding Patterns under Normal and Obesogenic Conditions. Biomolecules 2022, 12, 293. https://doi.org/10.3390/biom12020293
Lee JH, Xue B, Chen Z, Sun Y. Neuronal GHS-R Differentially Modulates Feeding Patterns under Normal and Obesogenic Conditions. Biomolecules. 2022; 12(2):293. https://doi.org/10.3390/biom12020293
Chicago/Turabian StyleLee, Jong Han, Bingzhong Xue, Zheng Chen, and Yuxiang Sun. 2022. "Neuronal GHS-R Differentially Modulates Feeding Patterns under Normal and Obesogenic Conditions" Biomolecules 12, no. 2: 293. https://doi.org/10.3390/biom12020293
APA StyleLee, J. H., Xue, B., Chen, Z., & Sun, Y. (2022). Neuronal GHS-R Differentially Modulates Feeding Patterns under Normal and Obesogenic Conditions. Biomolecules, 12(2), 293. https://doi.org/10.3390/biom12020293