
Reversal of G-Quadruplexes’ Role in Translation Control When Present in the Context of an IRES


Abstract
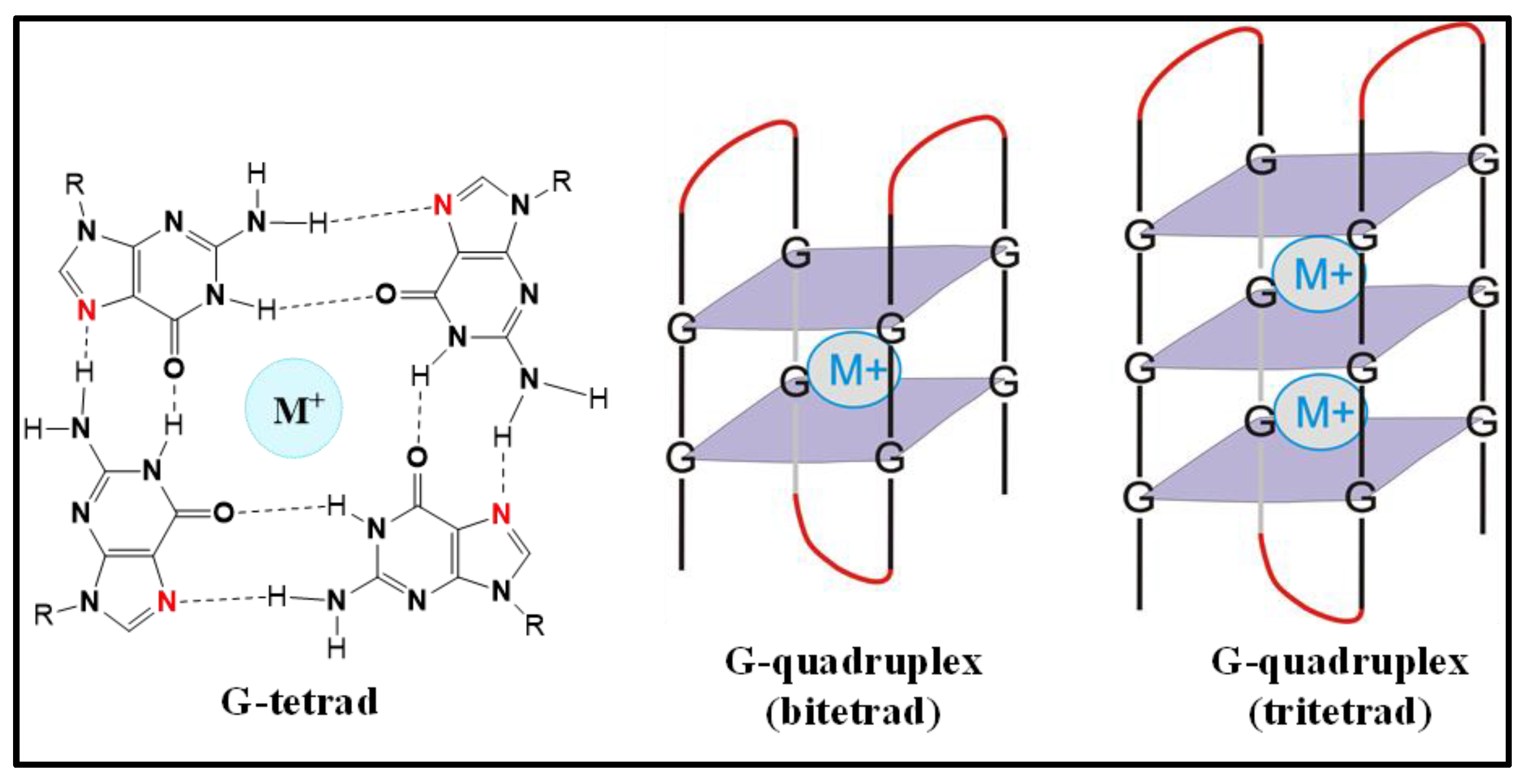
:1. Introduction
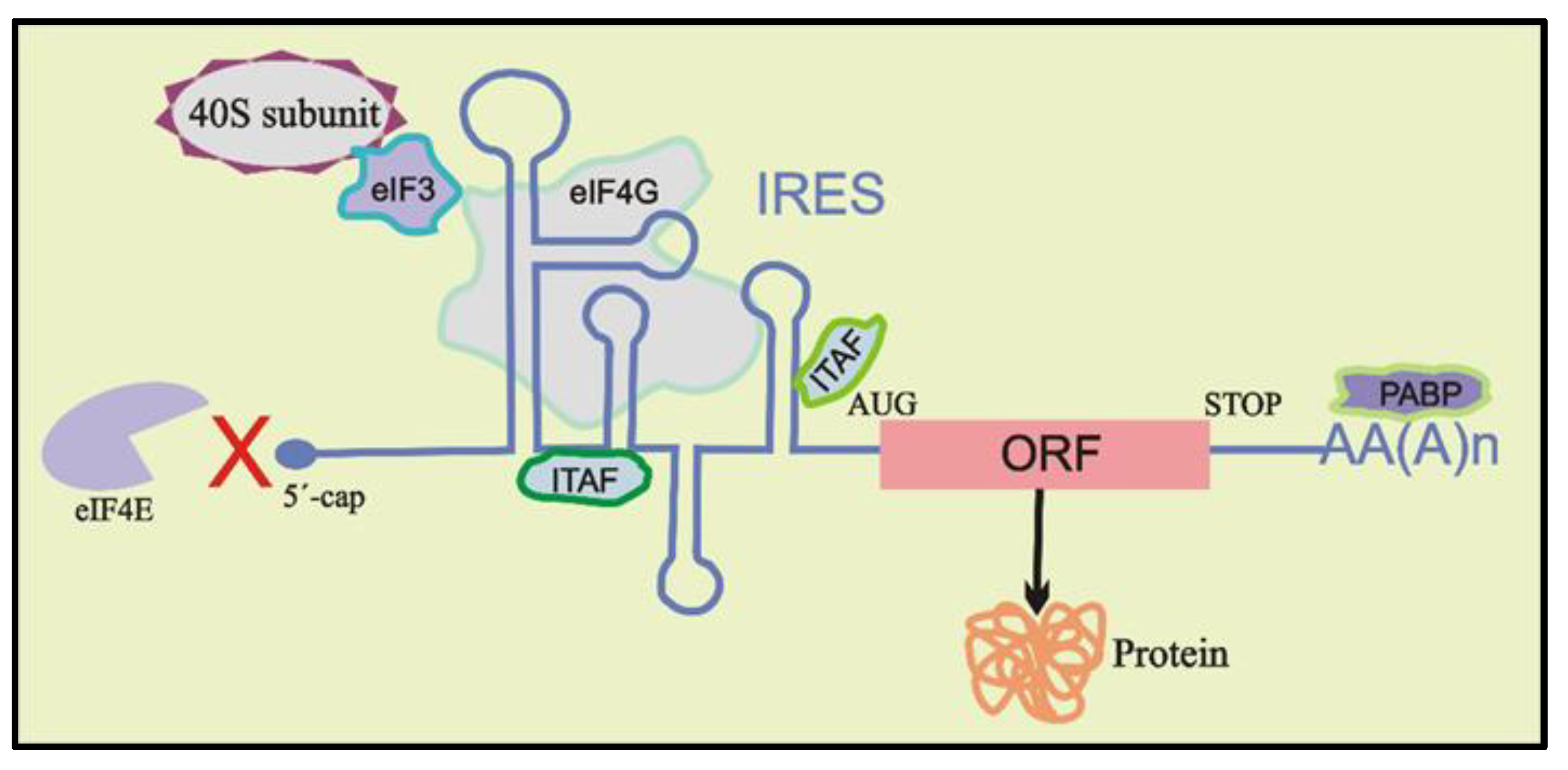
2. The Role of RNA G-Quadruplexes on IRES-Driven Cap-Independent Translation
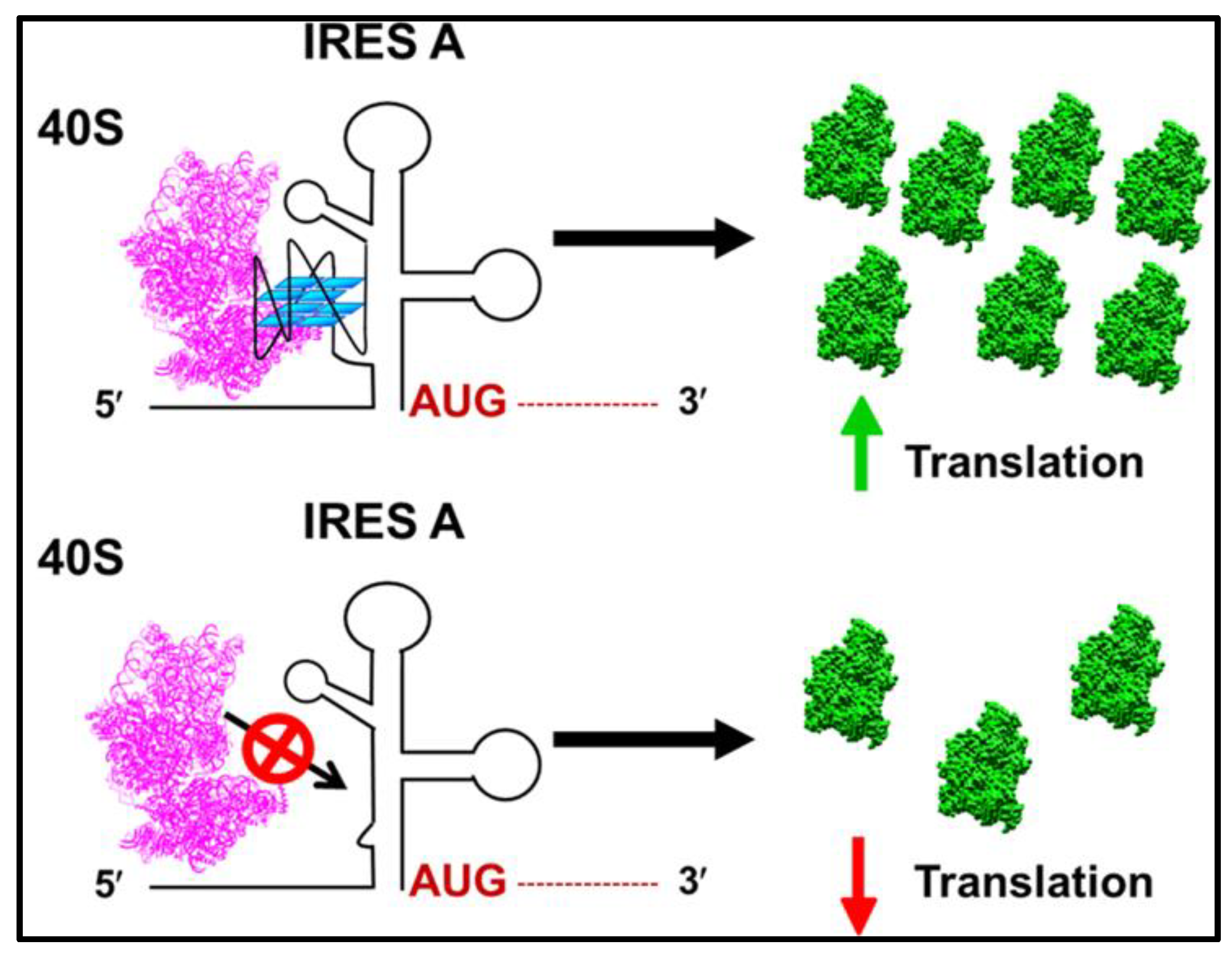
3. Mechanism of the Role of G-Quadruplexes in IRES-Mediated Translation Initiation
4. Future Perspectives of G-Quadruplexes’ Effects on IRES-Mediated Translation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bhattacharyya, D.; Mirihana Arachchilage, G.; Basu, S. Metal Cations in G-Quadruplex Folding and Stability. Front. Chem. 2016, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, D.E.; Feigon, J. Multistranded DNA Structures. Curr. Opin. Struct. Biol. 1999, 9, 305–314. [Google Scholar] [CrossRef]
- Henderson, E.; Hardin, C.C.; Walk, S.K.; Tinoco, I.; Blackburn, E.H. Telomeric DNA Oligonucleotides Form Novel Intramolecular Structures Containing Guanine-Guanine Base Pairs. Cell 1987, 51, 899–908. [Google Scholar] [CrossRef]
- Kim, J.; Cheong, C.; Moore, P.B. Tetramerization of an RNA Oligonucleotide Containing a GGGG Sequence. Nature 1991, 351, 331–332. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-Quadruplexes in Gene Promoters: A Novel Anticancer Strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, T.A.; Kendrick, S.; Hurley, L. Making Sense of G-Quadruplex and i-Motif Functions in Oncogene Promoters. FEBS J. 2010, 277, 3459–3469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Zhao, Y.; Li, N. Genome-Wide Analysis Reveals Regulatory Role of G4 DNA in Gene Transcription. Genome Res. 2008, 18, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.J.B.; Schultes, C.M.; Cuesta, J.; Cuenca, F.; Gunaratnam, M.; Tanious, F.A.; Wilson, W.D.; Neidle, S. Trisubstituted Acridines as G-Quadruplex Telomere Targeting Agents. Effects of Extensions of the 3,6- and 9-Side Chains on Quadruplex Binding, Telomerase Activity, and Cell Proliferation. J. Med. Chem. 2006, 49, 582–599. [Google Scholar] [CrossRef]
- Rizzo, A.; Salvati, E.; Porru, M.; D’Angelo, C.; Stevens, M.F.; D’Incalci, M.; Leonetti, C.; Gilson, E.; Zupi, G.; Biroccio, A. Stabilization of Quadruplex DNA Perturbs Telomere Replication Leading to the Activation of an ATR-Dependent ATM Signaling Pathway. Nucleic Acids Res. 2009, 37, 5353–5364. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct Evidence for a G-Quadruplex in a Promoter Region and Its Targeting with a Small Molecule to Repress c-MYC Transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef] [Green Version]
- Arora, A.; Dutkiewicz, M.; Scaria, V.; Hariharan, M.; Maiti, S.; Kurreck, J. Inhibition of Translation in Living Eukaryotic Cells by an RNA G-Quadruplex Motif. RNA 2008, 14, 1290–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, A.; Suess, B. An RNA G-Quadruplex in the 3’ UTR of the Proto-Oncogene PIM1 Represses Translation. RNA Biol. 2011, 8, 802–805. [Google Scholar] [CrossRef] [Green Version]
- Balkwill, G.D.; Derecka, K.; Garner, T.P.; Hodgman, C.; Flint, A.P.F.; Searle, M.S. Repression of Translation of Human Estrogen Receptor Alpha by G-Quadruplex Formation. Biochemistry 2009, 48, 11487–11495. [Google Scholar] [CrossRef] [PubMed]
- Bonnal, S.; Schaeffer, C.; Créancier, L.; Clamens, S.; Moine, H.; Prats, A.-C.; Vagner, S. A Single Internal Ribosome Entry Site Containing a G Quartet RNA Structure Drives Fibroblast Growth Factor 2 Gene Expression at Four Alternative Translation Initiation Codons. J. Biol. Chem. 2003, 278, 39330–39336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derecka, K.; Balkwill, G.D.; Garner, T.P.; Hodgman, C.; Flint, A.P.F.; Searle, M.S. Occurrence of a Quadruplex Motif in a Unique Insert within Exon C of the Bovine Estrogen Receptor α Gene (ESR1). Biochemistry 2010, 49, 7625–7633. [Google Scholar] [CrossRef]
- Gomez, D.; Guédin, A.; Mergny, J.-L.; Salles, B.; Riou, J.-F.; Teulade-Fichou, M.-P.; Calsou, P. A G-Quadruplex Structure within the 5′-UTR of TRF2 mRNA Represses Translation in Human Cells. Nucleic Acids Res. 2010, 38, 7187–7198. [Google Scholar] [CrossRef] [Green Version]
- Halder, K.; Wieland, M.; Hartig, J.S. Predictable Suppression of Gene Expression by 5′-UTR-Based RNA Quadruplexes. Nucleic Acids Res. 2009, 37, 6811–6817. [Google Scholar] [CrossRef]
- Kumari, S.; Bugaut, A.; Huppert, J.L.; Balasubramanian, S. An RNA G-Quadruplex in the 5′ UTR of the NRAS Proto-Oncogene Modulates Translation. Nat. Chem. Biol. 2007, 3, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Lammich, S.; Kamp, F.; Wagner, J.; Nuscher, B.; Zilow, S.; Ludwig, A.-K.; Willem, M.; Haass, C. Translational Repression of the Disintegrin and Metalloprotease ADAM10 by a Stable G-Quadruplex Secondary Structure in Its 5′-Untranslated Region. J. Biol. Chem. 2011, 286, 45063–45072. [Google Scholar] [CrossRef] [Green Version]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA Structures in Eukaryotic Translation Regulation and How to Find Them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef]
- Morris, M.J.; Negishi, Y.; Pazsint, C.; Schonhoft, J.D.; Basu, S. An RNA G-Quadruplex Is Essential for Cap-Independent Translation Initiation in Human VEGF IRES. J. Am. Chem. Soc. 2010, 132, 17831–17839. [Google Scholar] [CrossRef]
- Morris, M.J.; Basu, S. An Unusually Stable G-Quadruplex within the 5′-UTR of the MT3 Matrix Metalloproteinase mRNA Represses Translation in Eukaryotic Cells. Biochemistry 2009, 48, 5313–5319. [Google Scholar] [CrossRef]
- Shahid, R.; Bugaut, A.; Balasubramanian, S. The BCL-2 5′ Untranslated Region Contains an RNA G-Quadruplex-Forming Motif That Modulates Protein Expression. Biochemistry 2010, 49, 8300–8306. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Perreault, J.-P.; Topisirovic, I.; Richard, S. RNA G-Quadruplexes and Their Potential Regulatory Roles in Translation. Translation 2016, 4, e1244031. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, J.; Kofod, M.; Nielsen, F.C. A Guanosine Quadruplex and Two Stable Hairpins Flank a Major Cleavage Site in Insulin-like Growth Factor II mRNA. Nucleic Acids Res. 1994, 22, 5709–5716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decorsière, A.; Cayrel, A.; Vagner, S.; Millevoi, S. Essential Role for the Interaction between HnRNP H/F and a G Quadruplex in Maintaining P53 Pre-mRNA 3’-End Processing and Function during DNA Damage. Genes Dev. 2011, 25, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Wanrooij, P.H.; Uhler, J.P.; Simonsson, T.; Falkenberg, M.; Gustafsson, C.M. G-Quadruplex Structures in RNA Stimulate Mitochondrial Transcription Termination and Primer Formation. Proc. Natl. Acad. Sci. USA 2010, 107, 16072–16077. [Google Scholar] [CrossRef] [Green Version]
- Didiot, M.-C.; Tian, Z.; Schaeffer, C.; Subramanian, M.; Mandel, J.-L.; Moine, H. The G-Quartet Containing FMRP Binding Site in FMR1 mRNA Is a Potent Exonic Splicing Enhancer. Nucleic Acids Res. 2008, 36, 4902–4912. [Google Scholar] [CrossRef] [Green Version]
- Gomez, D.; Lemarteleur, T.; Lacroix, L.; Mailliet, P.; Mergny, J.-L.; Riou, J.-F. Telomerase Downregulation Induced by the G-Quadruplex Ligand 12459 in A549 Cells Is Mediated by HTERT RNA Alternative Splicing. Nucleic Acids Res. 2004, 32, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Zhang, J.; Harvey, S.E.; Hu, X.; Cheng, C. RNA G-Quadruplex Secondary Structure Promotes Alternative Splicing via the RNA-Binding Protein HnRNPF. Genes Dev. 2017, 31, 2296–2309. [Google Scholar] [CrossRef]
- Marcel, V.; Tran, P.L.T.; Sagne, C.; Martel-Planche, G.; Vaslin, L.; Teulade-Fichou, M.-P.; Hall, J.; Mergny, J.-L.; Hainaut, P.; Van Dyck, E. G-Quadruplex Structures in TP53 Intron 3: Role in Alternative Splicing and in Production of P53 MRNA Isoforms. Carcinogenesis 2011, 32, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.P.; Das, P. Novel Splicing in IGFN1 Intron 15 and Role of Stable G-Quadruplex in the Regulation of Splicing in Renal Cell Carcinoma. PLoS ONE 2018, 13, e0205660. [Google Scholar] [CrossRef]
- Imperatore, J.A.; McAninch, D.S.; Valdez-Sinon, A.N.; Bassell, G.J.; Mihailescu, M.R. FUS Recognizes G Quadruplex Structures Within Neuronal mRNAs. Front. Mol. Biosci. 2020, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Maltby, C.J.; Schofield, J.P.R.; Houghton, S.D.; O’Kelly, I.; Vargas-Caballero, M.; Deinhardt, K.; Coldwell, M.J. A 5′ UTR GGN Repeat Controls Localisation and Translation of a Potassium Leak Channel mRNA through G-Quadruplex Formation. Nucleic Acids Res. 2020, 48, 9822–9839. [Google Scholar] [CrossRef]
- Subramanian, M.; Rage, F.; Tabet, R.; Flatter, E.; Mandel, J.-L.; Moine, H. G-Quadruplex RNA Structure as a Signal for Neurite MRNA Targeting. EMBO Rep. 2011, 12, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Darnell, J.C.; Jensen, K.B.; Jin, P.; Brown, V.; Warren, S.T.; Darnell, R.B. Fragile X Mental Retardation Protein Targets G Quartet mRNAs Important for Neuronal Function. Cell 2001, 107, 489–499. [Google Scholar] [CrossRef]
- Schaeffer, C.; Bardoni, B.; Mandel, J.L.; Ehresmann, B.; Ehresmann, C.; Moine, H. The Fragile X Mental Retardation Protein Binds Specifically to Its mRNA via a Purine Quartet Motif. EMBO J. 2001, 20, 4803–4813. [Google Scholar] [CrossRef]
- Serikawa, T.; Spanos, C.; von Hacht, A.; Budisa, N.; Rappsilber, J.; Kurreck, J. Comprehensive Identification of Proteins Binding to RNA G-Quadruplex Motifs in the 5′ UTR of Tumor-Associated mRNAs. Biochimie 2018, 144, 169–184. [Google Scholar] [CrossRef]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA Binding to TRF2 Facilitates Heterochromatin Formation and ORC Recruitment at Telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Huppert, J.L.; Bugaut, A.; Kumari, S.; Balasubramanian, S. G-Quadruplexes: The Beginning and End of UTRs. Nucleic Acids Res. 2008, 36, 6260–6268. [Google Scholar] [CrossRef] [Green Version]
- López de Silanes, I.; Stagno d’Alcontres, M.; Blasco, M.A. TERRA Transcripts Are Bound by a Complex Array of RNA-Binding Proteins. Nat. Commun. 2010, 1, 33. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.J.; Phan, A.T.; Kuryavyi, V. Human Telomere, Oncogenic Promoter and 5′-UTR G-Quadruplexes: Diverse Higher Order DNA and RNA Targets for Cancer Therapeutics. Nucleic Acids Res. 2007, 35, 7429–7455. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Suzuki, Y.; Ito, K.; Komiyama, M. Telomeric Repeat-Containing RNA Structure in Living Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 14579–14584. [Google Scholar] [CrossRef] [Green Version]
- Cheong, C.; Moore, P.B. Solution Structure of an Unusually Stable RNA Tetraplex Containing G- and U-Quartet Structures. Biochemistry 1992, 31, 8406–8414. [Google Scholar] [CrossRef]
- Liu, H.; Matsugami, A.; Katahira, M.; Uesugi, S. A Dimeric RNA Quadruplex Architecture Comprised of Two G:G(:A):G:G(:A) Hexads, G:G:G:G Tetrads and UUUU Loops. J. Mol. Biol. 2002, 322, 955–970. [Google Scholar] [CrossRef]
- Saccà, B.; Lacroix, L.; Mergny, J.-L. The Effect of Chemical Modifications on the Thermal Stability of Different G-Quadruplex-Forming Oligonucleotides. Nucleic Acids Res. 2005, 33, 1182–1192. [Google Scholar] [CrossRef] [Green Version]
- Cobbold, L.C.; Spriggs, K.A.; Haines, S.J.; Dobbyn, H.C.; Hayes, C.; de Moor, C.H.; Lilley, K.S.; Bushell, M.; Willis, A.E. Identification of Internal Ribosome Entry Segment (IRES)-Trans-Acting Factors for the Myc Family of IRESs. Mol. Cell. Biol. 2008, 28, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Garant, J.-M.; Perreault, J.-P.; Scott, M.S. G4RNA Screener Web Server: User Focused Interface for RNA G-Quadruplex Prediction. Biochimie 2018, 151, 115–118. [Google Scholar] [CrossRef]
- Garst, A.D.; Batey, R.T. A Switch in Time: Detailing the Life of a Riboswitch. Biochim. Biophys. Acta 2009, 1789, 584–591. [Google Scholar] [CrossRef] [Green Version]
- Kikin, O.; D’Antonio, L.; Bagga, P.S. QGRS Mapper: A Web-Based Server for Predicting G-Quadruplexes in Nucleotide Sequences. Nucleic Acids Res. 2006, 34, W676–W682. [Google Scholar] [CrossRef]
- Agarwala, P.; Pandey, S.; Mapa, K.; Maiti, S. The G-Quadruplex Augments Translation in the 5′ Untranslated Region of Transforming Growth Factor Β2. Biochemistry 2013, 52, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Bornes, S.; Boulard, M.; Hieblot, C.; Zanibellato, C.; Iacovoni, J.S.; Prats, H.; Touriol, C. Control of the Vascular Endothelial Growth Factor Internal Ribosome Entry Site (IRES) Activity and Translation Initiation by Alternatively Spliced Coding Sequences. J. Biol. Chem. 2004, 279, 18717–18726. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.; Morris, M.J.; Kharel, P.; Mirihana Arachchilage, G.; Fedeli, K.M.; Basu, S. Engineered Domain Swapping Indicates Context Dependent Functional Role of RNA G-Quadruplexes. Biochimie 2017, 137, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Bugaut, A.; Balasubramanian, S. 5′-UTR RNA G-Quadruplexes: Translation Regulation and Targeting. Nucleic Acids Res. 2012, 40, 4727–4741. [Google Scholar] [CrossRef] [Green Version]
- Beaudoin, J.-D.; Perreault, J.-P. 5′-UTR G-Quadruplex Structures Acting as Translational Repressors. Nucleic Acids Res. 2010, 38, 7022–7036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serikawa, T.; Eberle, J.; Kurreck, J. Effects of Genomic Disruption of a Guanine Quadruplex in the 5′ UTR of the Bcl-2 MRNA in Melanoma Cells. FEBS Lett. 2017, 591, 3649–3659. [Google Scholar] [CrossRef] [Green Version]
- Rouleau, S.G.; Beaudoin, J.-D.; Bisaillon, M.; Perreault, J.-P. Small Antisense Oligonucleotides against G-Quadruplexes: Specific mRNA Translational Switches. Nucleic Acids Res. 2015, 43, 595–606. [Google Scholar] [CrossRef] [Green Version]
- Menon, L.; Mader, S.A.; Mihailescu, M.-R. Fragile X Mental Retardation Protein Interactions with the Microtubule Associated Protein 1B RNA. RNA 2008, 14, 1644–1655. [Google Scholar] [CrossRef] [Green Version]
- Melko, M.; Bardoni, B. The Role of G-Quadruplex in RNA Metabolism: Involvement of FMRP and FMR2P. Biochimie 2010, 92, 919–926. [Google Scholar] [CrossRef]
- Wolfe, A.L.; Singh, K.; Zhong, Y.; Drewe, P.; Rajasekhar, V.K.; Sanghvi, V.R.; Mavrakis, K.J.; Jiang, M.; Roderick, J.E.; Van der Meulen, J.; et al. RNA G-Quadruplexes Cause EIF4A-Dependent Oncogene Translation in Cancer. Nature 2014, 513, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Weng, H.-Y.; Huang, H.-L.; Zhao, P.-P.; Zhou, H.; Qu, L.-H. Translational Repression of Cyclin D3 by a Stable G-Quadruplex in Its 5′ UTR. RNA Biol. 2012, 9, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Murat, P.; Marsico, G.; Herdy, B.; Ghanbarian, A.; Portella, G.; Balasubramanian, S. RNA G-Quadruplexes at Upstream Open Reading Frames Cause DHX36- and DHX9-Dependent Translation of Human MRNAs. Genome Biol. 2018, 19, 229. [Google Scholar] [CrossRef]
- Jodoin, R.; Bauer, L.; Garant, J.-M.; Mahdi Laaref, A.; Phaneuf, F.; Perreault, J.-P. The Folding of 5′-UTR Human G-Quadruplexes Possessing a Long Central Loop. RNA 2014, 20, 1129–1141. [Google Scholar] [CrossRef] [Green Version]
- Jodoin, R.; Perreault, J.-P. G-Quadruplexes Formation in the 5′UTRs of mRNAs Associated with Colorectal Cancer Pathways. PLoS ONE 2018, 13, e0208363. [Google Scholar] [CrossRef]
- Lacerda, R.; Menezes, J.; Romão, L. More than Just Scanning: The Importance of Cap-Independent mRNA Translation Initiation for Cellular Stress Response and Cancer. Cell. Mol. Life Sci. CMLS 2017, 74, 1659–1680. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z. IRES-Mediated Cap-Independent Translation, a Path Leading to Hidden Proteome. J. Mol. Cell Biol. 2019, 11, 911–919. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Lu, H. Conjunction of Potential G-Quadruplex and Adjacent Cis-Elements in the 5′ UTR of Hepatocyte Nuclear Factor 4-Alpha Strongly Inhibit Protein Expression. Sci. Rep. 2017, 7, 17444. [Google Scholar] [CrossRef]
- Al-Zeer, M.A.; Dutkiewicz, M.; von Hacht, A.; Kreuzmann, D.; Röhrs, V.; Kurreck, J. Alternatively Spliced Variants of the 5′-UTR of the ARPC2 mRNA Regulate Translation by an Internal Ribosome Entry Site (IRES) Harboring a Guanine-Quadruplex Motif. RNA Biol. 2019, 16, 1622–1632. [Google Scholar] [CrossRef] [PubMed]
- Koukouraki, P.; Doxakis, E. Constitutive Translation of Human α-Synuclein Is Mediated by the 5′-Untranslated Region. Open Biol. 2016, 6, 160022. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Zhang, J.; Strom, J.; Yang, D.; Dinh, T.N.; Kappeler, K.; Chen, Q.M. G-Quadruplex in the NRF2 MRNA 5′ Untranslated Region Regulates De Novo NRF2 Protein Translation under Oxidative Stress. Mol. Cell. Biol. 2016, 37, e00122-16. [Google Scholar] [CrossRef] [Green Version]
- Waller, Z.A.E.; Howell, L.A.; MacDonald, C.J.; O’Connell, M.A.; Searcey, M. Identification and Characterisation of a G-Quadruplex Forming Sequence in the Promoter Region of Nuclear Factor (Erythroid-Derived 2)-like 2 (Nrf2). Biochem. Biophys. Res. Commun. 2014, 447, 128–132. [Google Scholar] [CrossRef]
- Jodoin, R.; Carrier, J.C.; Rivard, N.; Bisaillon, M.; Perreault, J.-P. G-Quadruplex Located in the 5′UTR of the BAG-1 mRNA Affects Both Its Cap-Dependent and Cap-Independent Translation through Global Secondary Structure Maintenance. Nucleic Acids Res. 2019, 47, 10247–10266. [Google Scholar] [CrossRef]
- Marques-Ramos, A.; Candeias, M.M.; Menezes, J.; Lacerda, R.; Willcocks, M.; Teixeira, A.; Locker, N.; Romão, L. Cap-Independent Translation Ensures MTOR Expression and Function upon Protein Synthesis Inhibition. RNA 2017, 23, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brödel, A.K.; Sonnabend, A.; Roberts, L.O.; Stech, M.; Wüstenhagen, D.A.; Kubick, S. IRES-Mediated Translation of Membrane Proteins and Glycoproteins in Eukaryotic Cell-Free Systems. PLoS ONE 2013, 8, e82234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, K.D.; Semler, B.L. Bridging IRES Elements in MRNAs to the Eukaryotic Translation Apparatus. Biochim. Biophys. Acta 2009, 1789, 518–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godet, A.-C.; David, F.; Hantelys, F.; Tatin, F.; Lacazette, E.; Garmy-Susini, B.; Prats, A.-C. IRES Trans-Acting Factors, Key Actors of the Stress Response. Int. J. Mol. Sci. 2019, 20, 924. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Salas, E.; Piñeiro, D.; Fernández, N. Alternative Mechanisms to Initiate Translation in Eukaryotic mRNAs. Comp. Funct. Genomics 2012, 2012, e391546. [Google Scholar] [CrossRef] [Green Version]
- Shatsky, I.N.; Dmitriev, S.E.; Terenin, I.M.; Andreev, D.E. Cap- and IRES-Independent Scanning Mechanism of Translation Initiation as an Alternative to the Concept of Cellular IRESs. Mol. Cells 2010, 30, 285–293. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Diamond, P.; Basu, S. An Independently Folding RNA G-Quadruplex Domain Directly Recruits the 40S Ribosomal Subunit. Biochemistry 2015, 54, 1879–1885. [Google Scholar] [CrossRef]
- Hellen, C.U.; Sarnow, P. Internal Ribosome Entry Sites in Eukaryotic mRNA Molecules. Genes Dev. 2001, 15, 1593–1612. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The Mechanism of Eukaryotic Translation Initiation and Principles of Its Regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komar, A.A.; Mazumder, B.; Merrick, W.C. A New Framework for Understanding IRES-Mediated Translation. Gene 2012, 502, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weingarten-Gabbay, S.; Elias-Kirma, S.; Nir, R.; Gritsenko, A.A.; Stern-Ginossar, N.; Yakhini, Z.; Weinberger, A.; Segal, E. Comparative Genetics. Systematic Discovery of Cap-Independent Translation Sequences in Human and Viral Genomes. Science 2016, 351, aad4939. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W.V. Alternative Ways to Think about Cellular Internal Ribosome Entry. J. Biol. Chem. 2010, 285, 29033–29038. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.J. The Current Status of Vertebrate Cellular MRNA IRESs. Cold Spring Harb. Perspect. Biol. 2013, 5, a011569. [Google Scholar] [CrossRef]
- Kozak, M. A Second Look at Cellular mRNA Sequences Said to Function as Internal Ribosome Entry Sites. Nucleic Acids Res. 2005, 33, 6593–6602. [Google Scholar] [CrossRef]
- Terenin, I.M.; Andreev, D.E.; Dmitriev, S.E.; Shatsky, I.N. A Novel Mechanism of Eukaryotic Translation Initiation That Is Neither M7G-Cap-, nor IRES-Dependent. Nucleic Acids Res. 2013, 41, 1807–1816. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.-B.; Jaffrey, S.R. 5′ UTR m(6)A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Rode, K.A.; Qian, S.-B. M(6)A: A Novel Hallmark of Translation. Cell Cycle Georget. Tex 2016, 15, 309–310. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.-B. Dynamic m(6)A MRNA Methylation Directs Translational Control of Heat Shock Response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef] [Green Version]
- Kharel, P.; Balaratnam, S.; Beals, N.; Basu, S. The Role of RNA G-Quadruplexes in Human Diseases and Therapeutic Strategies. Wiley Interdiscip. Rev. RNA 2020, 11, e1568. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The Regulation and Functions of DNA and RNA G-Quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 1–16. [Google Scholar] [CrossRef]
- Komar, A.A.; Hatzoglou, M. Cellular IRES-Mediated Translation: The War of ITAFs in Pathophysiological States. Cell Cycle Georget. Tex 2011, 10, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Spriggs, K.A.; Bushell, M.; Willis, A.E. Translational Regulation of Gene Expression during Conditions of Cell Stress. Mol. Cell 2010, 40, 228–237. [Google Scholar] [CrossRef]
- Pelletier, J.; Sonenberg, N. Internal Initiation of Translation of Eukaryotic mRNA Directed by a Sequence Derived from Poliovirus RNA. Nature 1988, 334, 320–325. [Google Scholar] [CrossRef]
- Lozano, G.; Martínez-Salas, E. Structural Insights into Viral IRES-Dependent Translation Mechanisms. Curr. Opin. Virol. 2015, 12, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Komar, A.A.; Hatzoglou, M. Exploring Internal Ribosome Entry Sites as Therapeutic Targets. Front. Oncol. 2015, 5, 233. [Google Scholar] [CrossRef] [Green Version]
- Hellen, C.U.T. IRES-Induced Conformational Changes in the Ribosome and the Mechanism of Translation Initiation by Internal Ribosomal Entry. Biochim. Biophys. Acta 2009, 1789, 558–570. [Google Scholar] [CrossRef] [Green Version]
- Balvay, L.; Rifo, R.S.; Ricci, E.P.; Decimo, D.; Ohlmann, T. Structural and Functional Diversity of Viral IRESes. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2009, 1789, 542–557. [Google Scholar] [CrossRef]
- Chen, T.-M.; Shih, Y.-H.; Tseng, J.T.; Lai, M.-C.; Wu, C.-H.; Li, Y.-H.; Tsai, S.-J.; Sun, H.S. Overexpression of FGF9 in Colon Cancer Cells Is Mediated by Hypoxia-Induced Translational Activation. Nucleic Acids Res. 2014, 42, 2932–2944. [Google Scholar] [CrossRef] [Green Version]
- Huez, I.; Créancier, L.; Audigier, S.; Gensac, M.C.; Prats, A.C.; Prats, H. Two Independent Internal Ribosome Entry Sites Are Involved in Translation Initiation of Vascular Endothelial Growth Factor mRNA. Mol. Cell. Biol. 1998, 18, 6178–6190. [Google Scholar] [CrossRef] [Green Version]
- Cammas, A.; Dubrac, A.; Morel, B.; Lamaa, A.; Touriol, C.; Teulade-Fichou, M.-P.; Prats, H.; Millevoi, S. Stabilization of the G-Quadruplex at the VEGF IRES Represses Cap-Independent Translation. RNA Biol. 2015, 12, 320–329. [Google Scholar] [CrossRef]
- von Hacht, A.; Seifert, O.; Menger, M.; Schütze, T.; Arora, A.; Konthur, Z.; Neubauer, P.; Wagner, A.; Weise, C.; Kurreck, J. Identification and Characterization of RNA Guanine-Quadruplex Binding Proteins. Nucleic Acids Res. 2014, 42, 6630–6644. [Google Scholar] [CrossRef]
- Li, W.; Thakor, N.; Xu, E.Y.; Huang, Y.; Chen, C.; Yu, R.; Holcik, M.; Kong, A.-N. An Internal Ribosomal Entry Site Mediates Redox-Sensitive Translation of Nrf2. Nucleic Acids Res. 2010, 38, 778–788. [Google Scholar] [CrossRef] [Green Version]
- Kieft, J.S.; Zhou, K.; Jubin, R.; Doudna, J.A. Mechanism of Ribosome Recruitment by Hepatitis C IRES RNA. RNA N. Y. N 2001, 7, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Sachs, A.B.; Sarnow, P.; Hentze, M.W. Starting at the Beginning, Middle, and End: Translation Initiation in Eukaryotes. Cell 1997, 89, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Poulin, F.; Sonenberg, N. Mechanism of Translation Initiation in Eukaryotes; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Merrick, W.C.; Pavitt, G.D. Protein Synthesis Initiation in Eukaryotic Cells. Cold Spring Harb. Perspect. Biol. 2018, 10, a033092. [Google Scholar] [CrossRef]
- Pain, V.M. Initiation of Protein Synthesis in Eukaryotic Cells. Eur. J. Biochem. 1996, 236, 747–771. [Google Scholar] [CrossRef]
- Merrick, W.C. Cap-Dependent and Cap-Independent Translation in Eukaryotic Systems. Gene 2004, 332, 1–11. [Google Scholar] [CrossRef]
- Sun, D.; Hurley, L.H. Biochemical Techniques for the Characterization of G-Quadruplex Structures: EMSA, DMS Footprinting, and DNA Polymerase Stop Assay. Methods Mol. Biol. Clifton NJ 2010, 608, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Kwok, C.K.; Marsico, G.; Sahakyan, A.B.; Chambers, V.S.; Balasubramanian, S. RG4-Seq Reveals Widespread Formation of G-Quadruplex Structures in the Human Transcriptome. Nat. Methods 2016, 13, 841–844. [Google Scholar] [CrossRef]
- Yang, T.-H.; Wang, C.-Y.; Tsai, H.-C.; Liu, C.-T. Human IRES Atlas: An Integrative Platform for Studying IRES-Driven Translational Regulation in Humans. Database J. Biol. Databases Curation 2021, 2021, baab025. [Google Scholar] [CrossRef] [PubMed]
- Mokrejs, M.; Masek, T.; Vopálensky, V.; Hlubucek, P.; Delbos, P.; Pospísek, M. IRESite—A Tool for the Examination of Viral and Cellular Internal Ribosome Entry Sites. Nucleic Acids Res. 2010, 38, D131–D136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokrejs, M.; Vopálenský, V.; Kolenaty, O.; Masek, T.; Feketová, Z.; Sekyrová, P.; Skaloudová, B.; Kríz, V.; Pospísek, M. IRESite: The Database of Experimentally Verified IRES Structures (Www.Iresite.Org). Nucleic Acids Res. 2006, 34, D125–D130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, Y.; Wang, C.; Zhang, H.; Zhang, H.; Jiang, B.; Guo, X.; Song, X. IRESbase: A Comprehensive Database of Experimentally Validated Internal Ribosome Entry Sites. Genom. Proteom. Bioinform. 2020, 18, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kolekar, P.; Pataskar, A.; Kulkarni-Kale, U.; Pal, J.; Kulkarni, A. IRESPred: Web Server for Prediction of Cellular and Viral Internal Ribosome Entry Site (IRES). Sci. Rep. 2016, 6, 27436. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, J.; Xu, T.; Yang, Q.; He, J.; Song, X. IRESfinder: Identifying RNA Internal Ribosome Entry Site in Eukaryotic Cell Using Framed k-Mer Features. J. Genet. Genom. 2018, 45, 403–406. [Google Scholar] [CrossRef]
- Wang, J.; Gribskov, M. IRESpy: An XGBoost Model for Prediction of Internal Ribosome Entry Sites. BMC Bioinformatics 2019, 20, 409. [Google Scholar] [CrossRef]
- Bohálová, N.; Cantara, A.; Bartas, M.; Kaura, P.; Šťastný, J.; Pečinka, P.; Fojta, M.; Mergny, J.-L.; Brázda, V. Analyses of Viral Genomes for G-Quadruplex Forming Sequences Reveal Their Correlation with the Type of Infection. Biochimie 2021, 186, 13–27. [Google Scholar] [CrossRef]
- Bedrat, A.; Lacroix, L.; Mergny, J.-L. Re-Evaluation of G-Quadruplex Propensity with G4Hunter. Nucleic Acids Res. 2016, 44, 1746–1759. [Google Scholar] [CrossRef]
- Yadav, V.K.; Abraham, J.K.; Mani, P.; Kulshrestha, R.; Chowdhury, S. QuadBase: Genome-Wide Database of G4 DNA—Occurrence and Conservation in Human, Chimpanzee, Mouse and Rat Promoters and 146 Microbes. Nucleic Acids Res. 2008, 36, D381–D385. [Google Scholar] [CrossRef] [PubMed]
- Brázda, V.; Kolomazník, J.; Lýsek, J.; Bartas, M.; Fojta, M.; Šťastný, J.; Mergny, J.-L. G4Hunter Web Application: A Web Server for G-Quadruplex Prediction. Bioinformatics 2019, 35, 3493–3495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garant, J.-M.; Perreault, J.-P.; Scott, M.S. Motif Independent Identification of Potential RNA G-Quadruplexes by G4RNA Screener. Bioinformatics 2017, 33, 3532–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaudoin, J.-D.; Jodoin, R.; Perreault, J.-P. New Scoring System to Identify RNA G-Quadruplex Folding. Nucleic Acids Res. 2014, 42, 1209–1223. [Google Scholar] [CrossRef] [Green Version]
- Kwok, C.K.; Merrick, C.J. G-Quadruplexes: Prediction, Characterization, and Biological Application. Trends Biotechnol. 2017, 35, 997–1013. [Google Scholar] [CrossRef]
- Puig Lombardi, E.; Londoño-Vallejo, A. A Guide to Computational Methods for G-Quadruplex Prediction. Nucleic Acids Res. 2020, 48, 1–15. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name of the mRNAs | Predicted Highest G Score (QGRS Mapper) | cGcC | G4H | G4NN |
|---|---|---|---|---|
| Leucine zipper protein 6 (LUZP6) | 40 | 6.7778 | 2.1176 | 0.9985 |
| Patched 1 (PTCH1) | 60 | 6.0323 | 2.8000 | 0.9991 |
| Baculoviral IAP repeat containing 2 (BIRC2) | 42 | 2.6731 | 1.6333 | 0.9395 |
| Nuclear factor erythroid 2 like 2 (NFE2L2) | 35 | 1.5571 | 2.2963 | 0.9918 |
| MYCN proto-oncogene, bHLH transcription factor (MYCN) | 35 | 4.1154 | 1.7083 | 0.9746 |
| Lymphoid enhancer binding factor 1 (LEF1) | 36 | 2.6429 | 1.4286 | 0.9449 |
| F-box and WD repeat domain containing 7 (FBXW7) | 42 | 1.1585 | 1.8889 | 0.9857 |
| Fibroblast growth factor 2 (FGF2) | 41 | 3.2069 | 2.3070 | 0.9962 |
| APC regulator of WNT signaling pathway (APC) | 34 | 3.1556 | 1.2500 | 0.9211 |
| Serine hydroxymethyltransferase 1 (SHMT1) | 40 | 3.3871 | 1.6400 | 0.9595 |
| MAX network transcriptional repressor (MNT) | 38 | 3.7647 | 1.5000 | 0.8408 |
| Insulin receptor (INSR) | 34 | 2.0533 | 1.0000 | 0.6700 |
| RUNX family transcription factor 1 (RUNX1) | 40 | 1.1207 | 1.8333 | 0.9901 |
| SNF2 histone linker PHD RING helicase (SHPRH) | 42 | 7.3125 | 2.4000 | 0.9983 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoque, M.E.; Mahendran, T.; Basu, S. Reversal of G-Quadruplexes’ Role in Translation Control When Present in the Context of an IRES. Biomolecules 2022, 12, 314. https://doi.org/10.3390/biom12020314
Hoque ME, Mahendran T, Basu S. Reversal of G-Quadruplexes’ Role in Translation Control When Present in the Context of an IRES. Biomolecules. 2022; 12(2):314. https://doi.org/10.3390/biom12020314
Chicago/Turabian StyleHoque, Mohammed Enamul, Thulasi Mahendran, and Soumitra Basu. 2022. "Reversal of G-Quadruplexes’ Role in Translation Control When Present in the Context of an IRES" Biomolecules 12, no. 2: 314. https://doi.org/10.3390/biom12020314
APA StyleHoque, M. E., Mahendran, T., & Basu, S. (2022). Reversal of G-Quadruplexes’ Role in Translation Control When Present in the Context of an IRES. Biomolecules, 12(2), 314. https://doi.org/10.3390/biom12020314