
Diverse and Complementary Effects of Ghrelin and Obestatin



Abstract
:1. Introduction
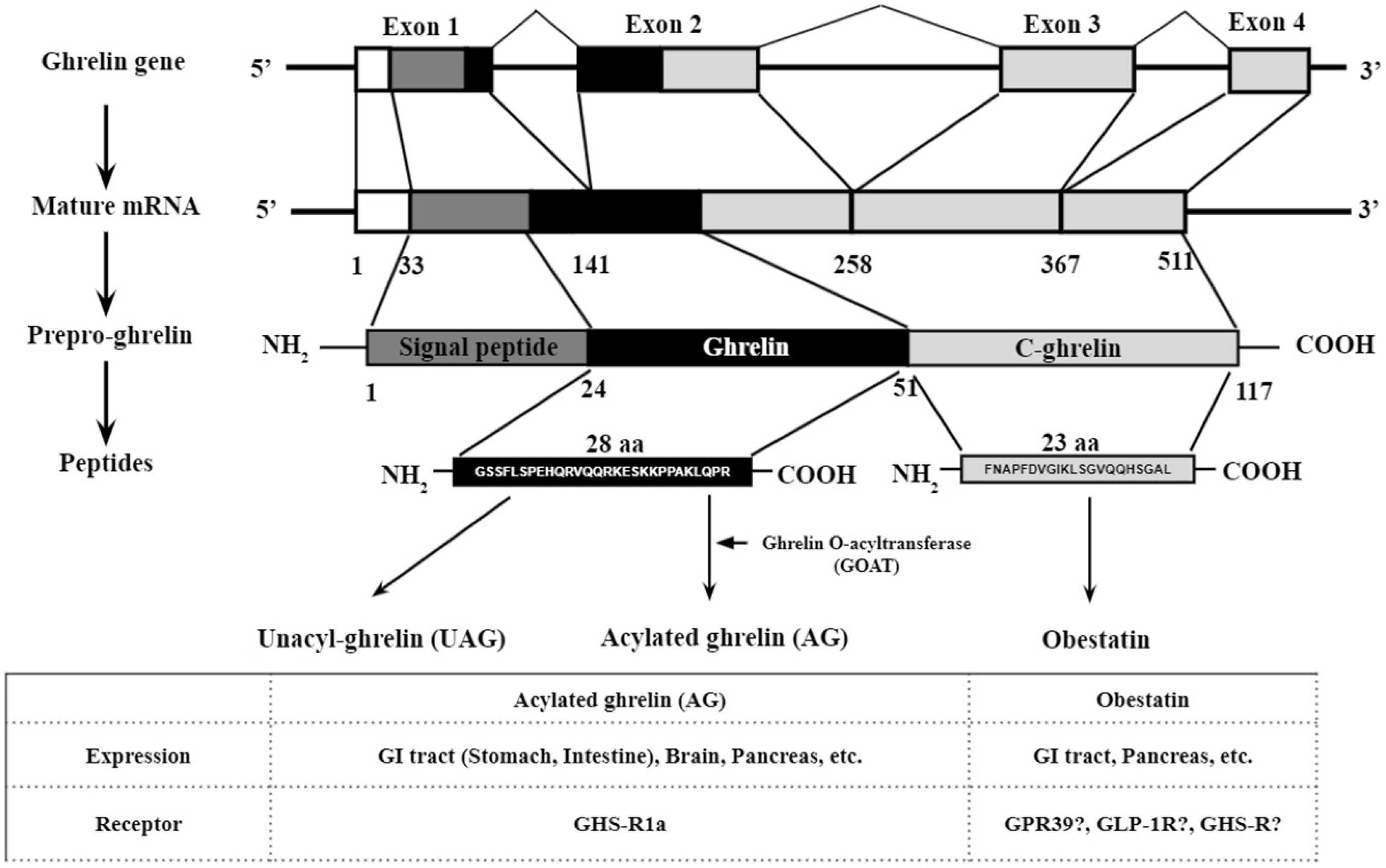
2. Gene Structure, Post Translational Processing, Expression, and Putative Receptors
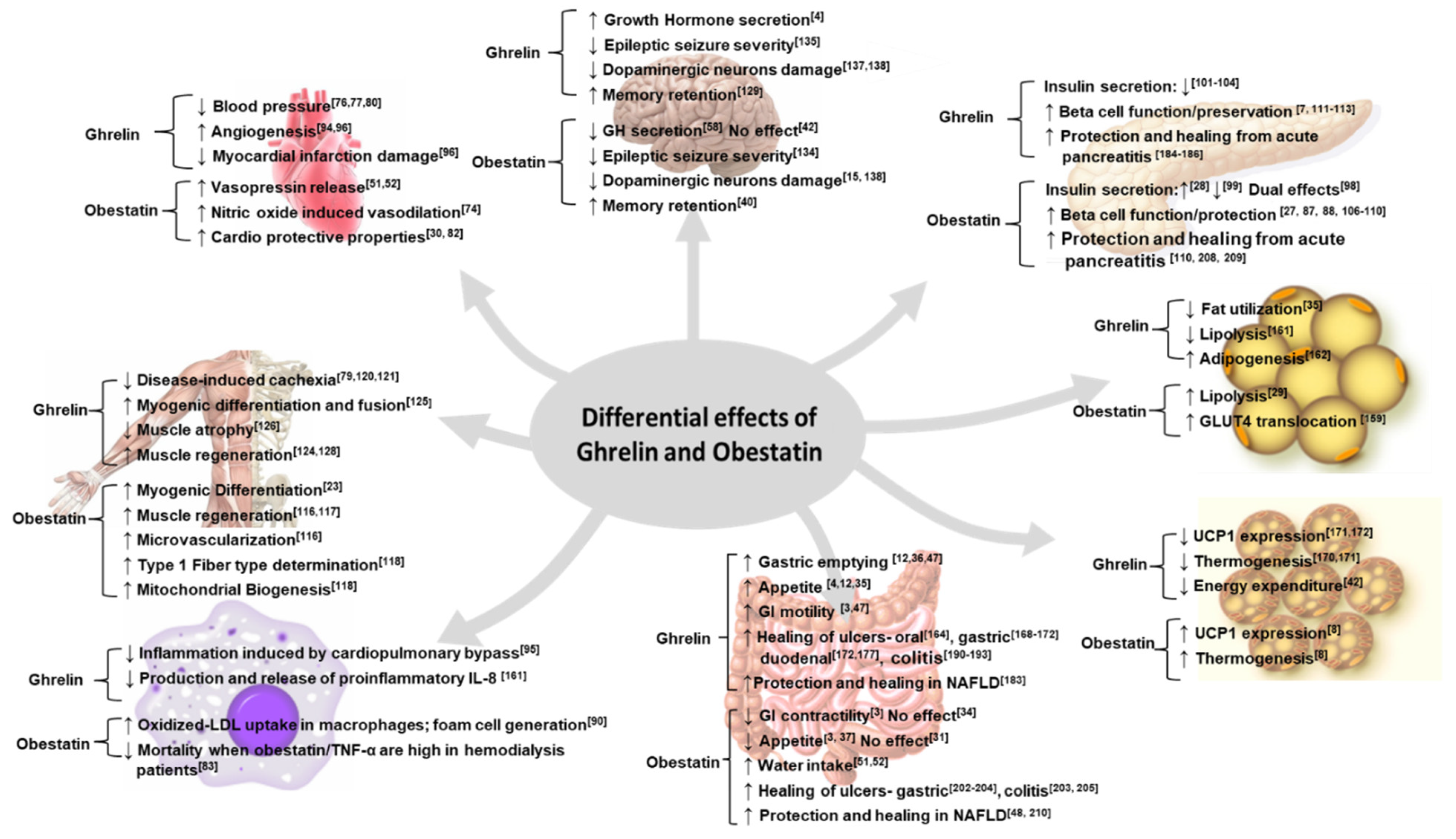
3. Obestatin and Ghrelin: Complex, Contradictory, and Complementary
3.1. Complex Functions
3.1.1. Food Intake, Weight Regulation, GI Motility, and Hydration
3.1.2. Diverse Expression Profiles under Fed and Fasted Conditions
3.1.3. Blood Pressure, Cardiovascular Disease, and Inflammation
3.1.4. Insulin Secretion
3.2. Complementary Functions
3.2.1. Pancreatic Beta Cell Protection
3.2.2. Muscle Function
3.2.3. Neuronal Function/Injury and Parkinson’s Disease
3.2.4. Cancer
3.2.5. Digestive System
3.3. Contradictory Functions
3.3.1. Lipid Metabolism
3.3.2. Thermogenesis
3.3.3. Sleep
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, X.; Coulombe-Huntington, J.; Kang, S.; Sheynkman, G.M.; Hao, T.; Richardson, A.; Sun, S.; Yang, F.; Shen, Y.A.; Murray, R.R.; et al. Widespread Expansion of Protein Interaction Capabilities by Alternative Splicing. Cell 2016, 164, 805–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blencowe, B.J. Alternative Splicing: New Insights from Global Analyses. Cell 2006, 126, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.V.; Ren, P.G.; Avsian-Kretchmer, O.; Luo, C.W.; Rauch, R.; Klein, C.; Hsueh, A.J. Obestatin, a peptide encoded by the ghrelin gene, opposes ghrelin’s effects on food intake. Science 2005, 310, 996–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepankiewicz, D.; Skrzypski, M.; Pruszynska-Oszmalek, E.; Zimmermann, D.; Andralojc, K.; Kaczmarek, P.; Wojciechowicz, T.; Sassek, M.; Nowak, K.W. Importance of ghrelin in hypothalamus-pituitary axis on growth hormone release during normal pregnancy in the rat. J. Physiol. Pharmacol. 2010, 61, 443–449. [Google Scholar] [PubMed]
- Andralojc, K.M.; Mercalli, A.; Nowak, K.W.; Albarello, L.; Calcagno, R.; Luzi, L.; Bonifacio, E.; Doglioni, C.; Piemonti, L. Ghrelin-producing epsilon cells in the developing and adult human pancreas. Diabetologia 2009, 52, 486–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakawa, A.; Inui, A.; Kaga, T.; Yuzuriha, H.; Nagata, T.; Ueno, N.; Makino, S.; Fujimiya, M.; Niijima, A.; Fujino, M.A.; et al. Ghrelin is an appetite-stimulatory signal from stomach with structural resemblance to motilin. Gastroenterology 2001, 120, 337–345. [Google Scholar] [CrossRef]
- Adeghate, E.; Ponery, A.S. Ghrelin stimulates insulin secretion from the pancreas of normal and diabetic rats. J. Neuroendocrinol. 2002, 14, 555–560. [Google Scholar] [CrossRef]
- Lin, L.; Lee, J.H.; Bongmba, O.Y.; Ma, X.; Zhu, X.; Sheikh-Hamad, D.; Sun, Y. The suppression of ghrelin signaling mitigates age-associated thermogenic impairment. Aging 2014, 6, 1019–1032. [Google Scholar] [CrossRef] [Green Version]
- Cowan, E.; Burch, K.J.; Green, B.D.; Grieve, D.J. Obestatin as a key regulator of metabolism and cardiovascular function with emerging therapeutic potential for diabetes. Br. J. Pharmacol. 2016, 173, 2165–2181. [Google Scholar] [CrossRef] [Green Version]
- Delporte, C. Structure and physiological actions of ghrelin. Scientifica (Cairo) 2013, 2013, 518909. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, H.; Gutierrez, J.A.; Solenberg, P.J.; Pfluger, P.T.; Czyzyk, T.A.; Willency, J.A.; Schurmann, A.; Joost, H.G.; Jandacek, R.J.; Hale, J.E.; et al. GOAT links dietary lipids with the endocrine control of energy balance. Nat. Med. 2009, 15, 741–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 4679–4684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, Z.B.; Liu, Z.W.; Walllingford, N.; Erion, D.M.; Borok, E.; Friedman, J.M.; Tschop, M.H.; Shanabrough, M.; Cline, G.; Shulman, G.I.; et al. UCP2 mediates ghrelin’s action on NPY/AgRP neurons by lowering free radicals. Nature 2008, 454, 846–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, J.S.; Kotokorpi, P.; Eccles, S.R.; Barnes, S.K.; Tokarczuk, P.F.; Allen, S.K.; Whitworth, H.S.; Guschina, I.A.; Evans, B.A.; Mode, A.; et al. Ghrelin induces abdominal obesity via GHS-R-dependent lipid retention. Mol. Endocrinol. 2009, 23, 914–924. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Yu, T.; Tang, Z.H.; Wu, K.M. Changes in ghrelin and obestatin levels before and after a meal in children with simple obesity and anorexia. Hormone Res. Pediatr. 2013, 79, 341–346. [Google Scholar] [CrossRef]
- Broglio, F.; Gottero, C.; Prodam, F.; Gauna, C.; Muccioli, G.; Papotti, M.; Abribat, T.; Van Der Lely, A.J.; Ghigo, E. Non-acylated ghrelin counteracts the metabolic but not the neuroendocrine response to acylated ghrelin in humans. J. Clin. Endocrinol. Metab. 2004, 89, 3062–3065. [Google Scholar] [CrossRef]
- Toshinai, K.; Yamaguchi, H.; Sun, Y.; Smith, R.G.; Yamanaka, A.; Sakurai, T.; Date, Y.; Mondal, M.S.; Shimbara, T.; Kawagoe, T.; et al. Des-acyl ghrelin induces food intake by a mechanism independent of the growth hormone secretagogue receptor. Endocrinology 2006, 147, 2306–2314. [Google Scholar] [CrossRef]
- Scrima, M.; Campiglia, P.; Esposito, C.; Gomez-Monterrey, I.; Novellino, E.; D’Ursi, A.M. Obestatin conformational features: A strategy to unveil obestatin’s biological role? Biochem. Biophys. Res. Commun. 2007, 363, 500–505. [Google Scholar] [CrossRef]
- Zhao, C.M.; Furnes, M.W.; Stenstrom, B.; Kulseng, B.; Chen, D. Characterization of obestatin- and ghrelin-producing cells in the gastrointestinal tract and pancreas of rats: An immunohistochemical and electron-microscopic study. Cell Tissue Res. 2008, 331, 575–587. [Google Scholar] [CrossRef]
- Muller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef]
- Gronberg, M.; Amini, R.M.; Stridsberg, M.; Janson, E.T.; Saras, J. Neuroendocrine markers are expressed in human mammary glands. Regul. Pept. 2010, 160, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Gronberg, M.; Tsolakis, A.V.; Magnusson, L.; Janson, E.T.; Saras, J. Distribution of obestatin and ghrelin in human tissues: Immunoreactive cells in the gastrointestinal tract, pancreas, and mammary glands. J. Histochem. Cytochem. 2008, 56, 793–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurriaran-Rodriguez, U.; Santos-Zas, I.; Al-Massadi, O.; Mosteiro, C.S.; Beiroa, D.; Nogueiras, R.; Crujeiras, A.B.; Seoane, L.M.; Senaris, J.; Garcia-Caballero, T.; et al. The obestatin/GPR39 system is up-regulated by muscle injury and functions as an autocrine regenerative system. J. Biol. Chem. 2012, 287, 38379–38389. [Google Scholar] [CrossRef] [Green Version]
- Moretti, E.; Vindigni, C.; Tripodi, S.A.; Mazzi, L.; Nuti, R.; Figura, N.; Collodel, G. Immunolocalisation of ghrelin and obestatin in human testis, seminal vesicles, prostate and spermatozoa. Andrologia 2014, 46, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Ozkan, Y.; Erman, F.; Gurates, B.; Kilic, N.; Colak, R.; Gundogan, T.; Catak, Z.; Bozkurt, M.; Akin, O.; et al. Presence of obestatin in breast milk: Relationship among obestatin, ghrelin, and leptin in lactating women. Nutrition 2008, 24, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Garcia, J.M.; Smith, R.G. Ghrelin and growth hormone secretagogue receptor expression in mice during aging. Endocrinology 2007, 148, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Granata, R.; Settanni, F.; Gallo, D.; Trovato, L.; Biancone, L.; Cantaluppi, V.; Nano, R.; Annunziata, M.; Campiglia, P.; Arnoletti, E.; et al. Obestatin promotes survival of pancreatic beta-cells and human islets and induces expression of genes involved in the regulation of beta-cell mass and function. Diabetes 2008, 57, 967–979. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, G.; Wu, C.S.; Han Lee, J.; Kanikarla, P.; Guo, S.; Yechoor, V.K.; Samson, S.L.; Sun, Y. Obestatin stimulates glucose-induced insulin secretion through ghrelin receptor GHS-R. Sci. Rep. 2017, 7, 979. [Google Scholar] [CrossRef]
- Pruszynska-Oszmalek, E.; Szczepankiewicz, D.; Hertig, I.; Skrzypski, M.; Sassek, M.; Kaczmarek, P.; Kolodziejski, P.A.; Mackowiak, P.; Nowak, K.W.; Strowski, M.Z.; et al. Obestatin inhibits lipogenesis and glucose uptake in isolated primary rat adipocytes. J. Biol. Regulat. Homeost. Agents 2013, 27, 23–33. [Google Scholar]
- Iglesias, M.J.; Salgado, A.; Pineiro, R.; Rodino, B.K.; Otero, M.F.; Grigorian, L.; Gallego, R.; Dieguez, C.; Gualillo, O.; Gonzalez-Juanatey, J.R.; et al. Lack of effect of the ghrelin gene-derived peptide obestatin on cardiomyocyte viability and metabolism. J. Endocrinol. Investig. 2007, 30, 470–476. [Google Scholar] [CrossRef]
- Gourcerol, G.; St-Pierre, D.H.; Tache, Y. Lack of obestatin effects on food intake: Should obestatin be renamed ghrelin-associated peptide (GAP)? Regul. Pept. 2007, 141, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Gordillo-Martinez, F.; Jiang, L.; He, P.; Hong, W.; Wei, X.; Staines, K.A.; Macrae, V.E.; Zhang, C.; Yu, D.; et al. Zinc ameliorates human aortic valve calcification through GPR39 mediated ERK1/2 signalling pathway. Cardiovasc. Res. 2021, 117, 820–835. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Egerod, K.L.; Jin, C.; Petersen, P.S.; Ostergaard, M.V.; Hald, J.; Sprinkel, A.M.; Storling, J.; Mandrup-Poulsen, T.; Holst, J.J.; et al. G protein-coupled receptor 39 deficiency is associated with pancreatic islet dysfunction. Endocrinology 2009, 150, 2577–2585. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, M.; Schmidt, F.N.; Guterman-Ram, G.; Khayyeri, H.; Hiram-Bab, S.; Orenbuch, A.; Katchkovsky, S.; Aflalo, A.; Isaksson, H.; Busse, B.; et al. Perturbed bone composition and integrity with disorganized osteoblast function in zinc receptor/Gpr39-deficient mice. FASEB J. 2018, 32, 2507–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, K.; Hasegawa, A.; Yamasaki, S.; Uchida, R.; Ohashi, W.; Kurashima, Y.; Kunisawa, J.; Kimura, S.; Iwanaga, T.; Watarai, H.; et al. Mast cells play role in wound healing through the ZnT2/GPR39/IL-6 axis. Sci. Rep. 2019, 9, 10842. [Google Scholar] [CrossRef]
- Zhu, D.; Su, Y.; Zheng, Y.; Fu, B.; Tang, L.; Qin, Y.X. Zinc regulates vascular endothelial cell activity through zinc-sensing receptor ZnR/GPR39. Am. J. Physiol. Cell. Physiol. 2018, 314, C404–C414. [Google Scholar] [CrossRef]
- Voelkl, J.; Tuffaha, R.; Luong, T.T.D.; Zickler, D.; Masyout, J.; Feger, M.; Verheyen, N.; Blaschke, F.; Kuro, O.M.; Tomaschitz, A.; et al. Zinc Inhibits Phosphate-Induced Vascular Calcification through TNFAIP3-Mediated Suppression of NF-kappaB. J. Am. Soc. Nephrol. 2018, 29, 1636–1648. [Google Scholar] [CrossRef] [Green Version]
- Laitakari, A.; Liu, L.; Frimurer, T.M.; Holst, B. The Zinc-Sensing Receptor GPR39 in Physiology and as a Pharmacological Target. Int. J. Mol. Sci. 2021, 22, 3872. [Google Scholar] [CrossRef]
- Xu, Y.; Barnes, A.P.; Alkayed, N.J. Role of GPR39 in Neurovascular Homeostasis and Disease. Int. J. Mol. Sci. 2021, 22, 8200. [Google Scholar] [CrossRef]
- Moran, B.M.; Miskelly, M.G.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; McKillop, A.M. Zinc-induced activation of GPR39 regulates glucose homeostasis through glucose-dependent insulinotropic polypeptide secretion from enteroendocrine K-cells. Biol. Chem. 2019. [Google Scholar] [CrossRef]
- Sharir, H.; Zinger, A.; Nevo, A.; Sekler, I.; Hershfinkel, M. Zinc released from injured cells is acting via the Zn2+-sensing receptor, ZnR, to trigger signaling leading to epithelial repair. J. Biol. Chem. 2010, 285, 26097–26106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, W.J.; Gee, K.R.; Kennedy, R.T. Imaging of Zn2+ release from pancreatic beta-cells at the level of single exocytotic events. Anal. Chem. 2003, 75, 3468–3475. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Kishi, T.; Lee, C.E.; Choi, B.J.; Fang, H.; Hollenberg, A.N.; Drucker, D.J.; Elmquist, J.K. Glucagon-like peptide-1-responsive catecholamine neurons in the area postrema link peripheral glucagon-like peptide-1 with central autonomic control sites. J. Neurosci. 2003, 23, 2939–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef]
- Unniappan, S.; Speck, M.; Kieffer, T.J. Metabolic effects of chronic obestatin infusion in rats. Peptides 2008, 29, 1354–1361. [Google Scholar] [CrossRef]
- Tschop, M.; Weyer, C.; Tataranni, P.A.; Devanarayan, V.; Ravussin, E.; Heiman, M.L. Circulating ghrelin levels are decreased in human obesity. Diabetes 2001, 50, 707–709. [Google Scholar] [CrossRef] [Green Version]
- Levin, F.; Edholm, T.; Schmidt, P.T.; Gryback, P.; Jacobsson, H.; Degerblad, M.; Hoybye, C.; Holst, J.J.; Rehfeld, J.F.; Hellstrom, P.M.; et al. Ghrelin stimulates gastric emptying and hunger in normal-weight humans. J. Clin. Endocrinol. Metab. 2006, 91, 3296–3302. [Google Scholar] [CrossRef]
- Yuan, X.; Cai, W.; Liang, X.F.; Su, H.; Yuan, Y.; Li, A.; Tao, Y.X. Obestatin partially suppresses ghrelin stimulation of appetite in "high-responders" grass carp, Ctenopharyngodon idellus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 184, 144–149. [Google Scholar] [CrossRef]
- Bresciani, E.; Rapetti, D.; Dona, F.; Bulgarelli, I.; Tamiazzo, L.; Locatelli, V.; Torsello, A. Obestatin inhibits feeding but does not modulate GH and corticosterone secretion in the rat. J. Endocrinol. Investig. 2006, 29, RC16–RC18. [Google Scholar] [CrossRef]
- Green, B.D.; Irwin, N.; Flatt, P.R. Direct and indirect effects of obestatin peptides on food intake and the regulation of glucose homeostasis and insulin secretion in mice. Peptides 2007, 28, 981–987. [Google Scholar] [CrossRef]
- Carlini, V.P.; Schioth, H.B.; Debarioglio, S.R. Obestatin improves memory performance and causes anxiolytic effects in rats. Biochem. Biophys. Res. Commun. 2007, 352, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Fujimiya, M.; Ataka, K.; Asakawa, A.; Chen, C.Y.; Kato, I.; Inui, A. Regulation of gastroduodenal motility: Acyl ghrelin, des-acyl ghrelin and obestatin and hypothalamic peptides. Digestion 2012, 85, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Nogueiras, R.; Pfluger, P.; Tovar, S.; Arnold, M.; Mitchell, S.; Morris, A.; Perez-Tilve, D.; Vazquez, M.J.; Wiedmer, P.; Castaneda, T.R.; et al. Effects of obestatin on energy balance and growth hormone secretion in rodents. Endocrinology 2007, 148, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Seoane, L.M.; Al-Massadi, O.; Pazos, Y.; Pagotto, U.; Casanueva, F.F. Central obestatin administration does not modify either spontaneous or ghrelin-induced food intake in rats. J. Endocrinol. Investig. 2006, 29, RC13–RC15. [Google Scholar] [CrossRef] [PubMed]
- De Smet, B.; Thijs, T.; Peeters, T.L.; Depoortere, I. Effect of peripheral obestatin on gastric emptying and intestinal contractility in rodents. Neurogastroenterol. Motil. 2007, 19, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Depoortere, I.; Thijs, T.; Moechars, D.; De Smet, B.; Ver Donck, L.; Peeters, T.L. Effect of peripheral obestatin on food intake and gastric emptying in ghrelin-knockout mice. Br. J. Pharmacol. 2008, 153, 1550–1557. [Google Scholar] [CrossRef] [Green Version]
- Gourcerol, G.; Million, M.; Adelson, D.W.; Wang, Y.; Wang, L.; Rivier, J.; St-Pierre, D.H.; Tache, Y. Lack of interaction between peripheral injection of CCK and obestatin in the regulation of gastric satiety signaling in rodents. Peptides 2006, 27, 2811–2819. [Google Scholar] [CrossRef]
- Bassil, A.K.; Haglund, Y.; Brown, J.; Rudholm, T.; Hellstrom, P.M.; Naslund, E.; Lee, K.; Sanger, G.J. Little or no ability of obestatin to interact with ghrelin or modify motility in the rat gastrointestinal tract. Br. J. Pharmacol. 2007, 150, 58–64. [Google Scholar] [CrossRef]
- Khaleel, E.F.; Abdel-Aleem, G.A. Obestatin protects and reverses nonalcoholic fatty liver disease and its associated insulin resistance in rats via inhibition of food intake, enhancing hepatic adiponectin signaling, and blocking ghrelin acylation. Arch. Physiol. Biochem. 2019, 125, 64–78. [Google Scholar] [CrossRef]
- Nagaraj, S.; Manjappara, U.V. Studies on the influence of CCK-8 on the ability of obestatin to reduce food intake, gain in body weight and related lipid parameters. Biochimie 2016, 125, 126–130. [Google Scholar] [CrossRef]
- Khirazova, E.E.; Maslova, M.V.; Motorykina, E.S.; Frid, D.A.; Graf, A.V.; Maklakova, A.S.; Sokolova, N.A.; Kamenskii, A.A. Effects of single intranasal administration of obestatin fragments on the body weight and feeding and drinking behaviors. Dokl. Biol. Sci. 2013, 453, 336–337. [Google Scholar] [CrossRef] [PubMed]
- Samson, W.K.; White, M.M.; Price, C.; Ferguson, A.V. Obestatin acts in brain to inhibit thirst. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R637–R643. [Google Scholar] [CrossRef] [PubMed]
- Samson, W.K.; Yosten, G.L.; Chang, J.K.; Ferguson, A.V.; White, M.M. Obestatin inhibits vasopressin secretion: Evidence for a physiological action in the control of fluid homeostasis. J. Endocrinol. 2008, 196, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, W.; Tu, H.; Kastin, A.J. Differential BBB interactions of three ingestive peptides: Obestatin, ghrelin, and adiponectin. Peptides 2006, 27, 911–916. [Google Scholar] [CrossRef]
- Vergote, V.; Van Dorpe, S.; Peremans, K.; Burvenich, C.; De Spiegeleer, B. In vitro metabolic stability of obestatin: Kinetics and identification of cleavage products. Peptides 2008, 29, 1740–1748. [Google Scholar] [CrossRef]
- Kojima, M.; Kangawa, K. Ghrelin: Structure and function. Physiol. Rev. 2005, 85, 495–522. [Google Scholar] [CrossRef]
- Mietlicki, E.G.; Nowak, E.L.; Daniels, D. The effect of ghrelin on water intake during dipsogenic conditions. Physiol. Behav. 2009, 96, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.F.; Ren, A.J.; Zheng, X.; Qin, Y.W.; Cheng, F.; Zhang, J.; Wu, H.; Yuan, W.J.; Zou, L. Different responses of circulating ghrelin, obestatin levels to fasting, re-feeding and different food compositions, and their local expressions in rats. Peptides 2008, 29, 1247–1254. [Google Scholar] [CrossRef]
- Zizzari, P.; Longchamps, R.; Epelbaum, J.; Bluet-Pajot, M.T. Obestatin partially affects ghrelin stimulation of food intake and growth hormone secretion in rodents. Endocrinology 2007, 148, 1648–1653. [Google Scholar] [CrossRef] [Green Version]
- Udum, D.; Belenli, D.; Ilhan, T.; Gunes, N.; Sonat, F.; Yalcin, M. Obestatin and Ghrelin May Have a Complementary Function During Acute and Chronic Period in Mice. Protein Pept. Lett. 2016, 23, 349–357. [Google Scholar] [CrossRef]
- Schalla, M.A.; Stengel, A. The Role of Ghrelin in Anorexia Nervosa. Int. J. Mol. Sci. 2018, 19, 2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, B.; Cuntz, U.; Fruehauf, E.; Wawarta, R.; Folwaczny, C.; Riepl, R.L.; Heiman, M.L.; Lehnert, P.; Fichter, M.; Tschop, M. Weight gain decreases elevated plasma ghrelin concentrations of patients with anorexia nervosa. Eur. J. Endocrinol. 2001, 145, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiiya, T.; Nakazato, M.; Mizuta, M.; Date, Y.; Mondal, M.S.; Tanaka, M.; Nozoe, S.; Hosoda, H.; Kangawa, K.; Matsukura, S. Plasma ghrelin levels in lean and obese humans and the effect of glucose on ghrelin secretion. J. Clin. Endocrinol. Metab. 2002, 87, 240–244. [Google Scholar] [CrossRef]
- Rigamonti, A.E.; Pincelli, A.I.; Corra, B.; Viarengo, R.; Bonomo, S.M.; Galimberti, D.; Scacchi, M.; Scarpini, E.; Cavagnini, F.; Muller, E.E. Plasma ghrelin concentrations in elderly subjects: Comparison with anorexic and obese patients. J. Endocrinol. 2002, 175, R1–R5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Dulawa, S.C. The Utility of Animal Models for Studying the Metabo-Psychiatric Origins of Anorexia Nervosa. Front. Psychiatry 2021, 12, 711181. [Google Scholar] [CrossRef] [PubMed]
- Tolle, V.; Kadem, M.; Bluet-Pajot, M.T.; Frere, D.; Foulon, C.; Bossu, C.; Dardennes, R.; Mounier, C.; Zizzari, P.; Lang, F.; et al. Balance in ghrelin and leptin plasma levels in anorexia nervosa patients and constitutionally thin women. J. Clin. Endocrinol. Metab. 2003, 88, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mequinion, M.; Caron, E.; Zgheib, S.; Stievenard, A.; Zizzari, P.; Tolle, V.; Cortet, B.; Lucas, S.; Prevot, V.; Chauveau, C.; et al. Physical activity: Benefit or weakness in metabolic adaptations in a mouse model of chronic food restriction? Am. J. Physiol. Endocrinol. Metab. 2015, 308, E241–E255. [Google Scholar] [CrossRef] [Green Version]
- Verhagen, L.A.; Egecioglu, E.; Luijendijk, M.C.; Hillebrand, J.J.; Adan, R.A.; Dickson, S.L. Acute and chronic suppression of the central ghrelin signaling system reveals a role in food anticipatory activity. Eur. Neuropsychopharmacol. 2011, 21, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Soriano-Guillen, L.; Barrios, V.; Campos-Barros, A.; Argente, J. Ghrelin levels in obesity and anorexia nervosa: Effect of weight reduction or recuperation. J. Pediatr. 2004, 144, 36–42. [Google Scholar] [CrossRef]
- Janas-Kozik, M.; Krupka-Matuszczyk, I.; Malinowska-Kolodziej, I.; Lewin-Kowalik, J. Total ghrelin plasma level in patients with the restrictive type of anorexia nervosa. Regul. Pept. 2007, 140, 43–46. [Google Scholar] [CrossRef]
- Harada, T.; Nakahara, T.; Yasuhara, D.; Kojima, S.; Sagiyama, K.-i.; Amitani, H.; Laviano, A.; Naruo, T.; Inui, A. Obestatin, Acyl Ghrelin, and Des-acyl Ghrelin Responses to an Oral Glucose Tolerance Test in the Restricting Type of Anorexia Nervosa. Biol. Psychiatry 2008, 63, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Sedlackova, D.; Kopeckova, J.; Papezova, H.; Vybiral, S.; Kvasnickova, H.; Hill, M.; Nedvidkova, J. Changes of plasma obestatin, ghrelin and NPY in anorexia and bulimia nervosa patients before and after a high-carbohydrate breakfast. Physiol. Res. 2011, 60, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Germain, N.; Galusca, B.; Grouselle, D.; Frere, D.; Billard, S.; Epelbaum, J.; Estour, B. Ghrelin and obestatin circadian levels differentiate bingeing-purging from restrictive anorexia nervosa. J. Clin. Endocrinol. Metab. 2010, 95, 3057–3062. [Google Scholar] [CrossRef] [PubMed]
- Beasley, J.M.; Ange, B.A.; Anderson, C.A.M.; Miller, E.R.; Holbrook, J.T.; Appel, L.J. Characteristics associated with fasting appetite hormones (obestatin, ghrelin, and leptin). Obesity 2009, 17, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Hassouna, R.; Zizzari, P.; Tolle, V. The ghrelin/obestatin balance in the physiological and pathological control of growth hormone secretion, body composition and food intake. J. Neuroendocrinol. 2010, 22, 793–804. [Google Scholar] [CrossRef]
- Koca, S.S.; Kara, M.; Ozgen, M.; Dayanan, R.; Demir, C.F.; Aksoy, K.; Ilhan, N.; Donder, E.; Isik, A. Low prevalence of obesity in Behcet’s disease is associated with high obestatin level. Eur. J. Rheumatol. 2017, 4, 113–117. [Google Scholar] [CrossRef]
- Gao, X.Y.; Kuang, H.Y.; Liu, X.M.; Ma, Z.B. Decreased gastric body mucosa obestatin expression in overweight and obese patients. Peptides 2010, 31, 291–296. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Li, F.; Wang, J.P. Correlation analysis of serum obestatin expression with insulin resistance in childhood obesity. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Guo, Z.F.; Zheng, X.; Qin, Y.W.; Hu, J.Q.; Chen, S.P.; Zhang, Z. Circulating preprandial ghrelin to obestatin ratio is increased in human obesity. J. Clin. Endocrinol. Metab. 2007, 92, 1875–1880. [Google Scholar] [CrossRef] [Green Version]
- Vicennati, V.; Genghini, S.; De Iasio, R.; Pasqui, F.; Pagotto, U.; Pasquali, R. Circulating obestatin levels and the ghrelin/obestatin ratio in obese women. Eur. J. Endocrinol. 2007, 157, 295–301. [Google Scholar] [CrossRef]
- Zhang, N.; Yuan, C.; Li, Z.; Li, J.; Li, X.; Li, C.; Li, R.; Wang, S.R. Meta-analysis of the relationship between obestatin and ghrelin levels and the ghrelin/obestatin ratio with respect to obesity. Am. J. Med. Sci. 2011, 341, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Anderwald-Stadler, M.; Krebs, M.; Promintzer, M.; Mandl, M.; Bischof, M.G.; Nowotny, P.; Kastenbauer, T.; Luger, A.; Prager, R.; Anderwald, C. Plasma obestatin is lower at fasting and not suppressed by insulin in insulin-resistant humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1393–E1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.F.; Guo, Z.F.; Cao, J.; Hu, J.Q.; Zhao, X.X.; Xu, R.L.; Huang, X.M.; Qin, Y.W.; Zheng, X. Plasma ghrelin and obestatin levels are increased in spontaneously hypertensive rats. Peptides 2010, 31, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.F.; Song, S.W.; Qin, Y.W.; Zhang, J.L.; Zhao, X.X.; Zhang, B.L.; Ren, A.J.; Guo, Z.F.; Zheng, X. Bolus intravenous injection of obestatin does not change blood pressure level of spontaneously hypertensive rat. Peptides 2009, 30, 1928–1930. [Google Scholar] [CrossRef]
- Ren, A.J.; He, Q.; Shi, J.S.; Guo, Z.F.; Zheng, X.; Lin, L.; Wang, Y.K.; Xia, S.Y.; Sun, L.L.; Du, X.; et al. Association of obestatin with blood pressure in the third trimesters of pregnancy. Peptides 2009, 30, 1742–1745. [Google Scholar] [CrossRef]
- Li, Z.F.; Guo, Z.F.; Yang, S.G.; Zheng, X.; Cao, J.; Qin, Y.W. Circulating ghrelin and ghrelin to obestatin ratio are low in patients with untreated mild-to-moderate hypertension. Regul. Pept. 2010, 165, 206–209. [Google Scholar] [CrossRef]
- Schinzari, F.; Veneziani, A.; Mores, N.; Barini, A.; Di Daniele, N.; Cardillo, C.; Tesauro, M. Vascular Effects of Obestatin in Lean and Obese Subjects. Diabetes 2017, 66, 1214–1221. [Google Scholar] [CrossRef] [Green Version]
- Poykko, S.M.; Kellokoski, E.; Horkko, S.; Kauma, H.; Kesaniemi, Y.A.; Ukkola, O. Low plasma ghrelin is associated with insulin resistance, hypertension, and the prevalence of type 2 diabetes. Diabetes 2003, 52, 2546–2553. [Google Scholar] [CrossRef]
- Matsumura, K.; Tsuchihashi, T.; Fujii, K.; Abe, I.; Iida, M. Central ghrelin modulates sympathetic activity in conscious rabbits. Hypertension 2002, 40, 694–699. [Google Scholar] [CrossRef] [Green Version]
- Nagaya, N.; Kojima, M.; Uematsu, M.; Yamagishi, M.; Hosoda, H.; Oya, H.; Hayashi, Y.; Kangawa, K. Hemodynamic and hormonal effects of human ghrelin in healthy volunteers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1483–R1487. [Google Scholar] [CrossRef] [Green Version]
- Nagaya, N.; Miyatake, K.; Uematsu, M.; Oya, H.; Shimizu, W.; Hosoda, H.; Kojima, M.; Nakanishi, N.; Mori, H.; Kangawa, K. Hemodynamic, renal, and hormonal effects of ghrelin infusion in patients with chronic heart failure. J. Clin. Endocrinol. Metab. 2001, 86, 5854–5859. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, N.; Uematsu, M.; Kojima, M.; Ikeda, Y.; Yoshihara, F.; Shimizu, W.; Hosoda, H.; Hirota, Y.; Ishida, H.; Mori, H.; et al. Chronic administration of ghrelin improves left ventricular dysfunction and attenuates development of cardiac cachexia in rats with heart failure. Circulation 2001, 104, 1430–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, H.; Nakata, M.; Dezaki, K.; Lu, M.; Gantulga, D.; Yamamoto, K.; Shimada, K.; Kario, K.; Yada, T. Ghrelin counteracts salt-induced hypertension via promoting diuresis and renal nitric oxide production in Dahl rats. Endocr. J. 2013, 60, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.J.; Bidlingmaier, M.; Altaye, M.; Page, L.C.; D’Alessio, D.; Tschop, M.H.; Tong, J. Acute administration of acyl, but not desacyl ghrelin, decreases blood pressure in healthy humans. Eur. J. Endocrinol. 2017, 176, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alloatti, G.; Arnoletti, E.; Bassino, E.; Penna, C.; Perrelli, M.G.; Ghe, C.; Muccioli, G. Obestatin affords cardioprotection to the ischemic-reperfused isolated rat heart and inhibits apoptosis in cultures of similarly stressed cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H470–H481. [Google Scholar] [CrossRef]
- Beberashvili, I.; Katkov, A.; Sinuani, I.; Azar, A.; Shapiro, G.; Feldman, L.; Gorelik, O.; Stav, K.; Efrati, S. Serum obestatin: A biomarker of cardiovascular and all-cause mortality in hemodialysis patients. Am. J. Nephrol. 2018, 47, 254–265. [Google Scholar] [CrossRef]
- Beberashvili, I.; Sinuani, I.; Azar, A.; Kadoshi, H.; Shapiro, G.; Feldman, L.; Sandbank, J.; Averbukh, Z. Low serum concentration of obestatin as a predictor of mortality in maintenance hemodialysis patients. BioMed Res. Int. 2013, 2013, 796586. [Google Scholar] [CrossRef] [Green Version]
- Zorlu, M.; Kiskac, M.; Guler, E.M.; Gultepe, I.; Yavuz, E.; Celik, K.; Kocyigit, A. Serum obestatin and omentin levels in patients with diabetic nephropathy. Niger. J. Clin. Pract. 2017, 20, 182–187. [Google Scholar] [CrossRef]
- Stehouwer, C.D.; Smulders, Y.M. Microalbuminuria and risk for cardiovascular disease: Analysis of potential mechanisms. J. Am. Soc. Nephrol. 2006, 17, 2106–2111. [Google Scholar] [CrossRef] [Green Version]
- Baragli, A.; Grande, C.; Gesmundo, I.; Settanni, F.; Taliano, M.; Gallo, D.; Gargantini, E.; Ghigo, E.; Granata, R. Obestatin enhances in vitro generation of pancreatic islets through regulation of developmental pathways. PLoS ONE 2013, 8, e64374. [Google Scholar] [CrossRef]
- Favaro, E.; Granata, R.; Miceli, I.; Baragli, A.; Settanni, F.; Cavallo Perin, P.; Ghigo, E.; Camussi, G.; Zanone, M.M. The ghrelin gene products and exendin-4 promote survival of human pancreatic islet endothelial cells in hyperglycaemic conditions, through phosphoinositide 3-kinase/Akt, extracellular signal-related kinase (ERK)1/2 and cAMP/protein kinase A (PKA) signalling pathways. Diabetologia 2012, 55, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Kanat, B.H.; Ayten, R.; Aydin, S.; Girgin, M.; Cetinkaya, Z.; Ilhan, Y.S.; Yur, M.; Catak, Z. Significance of appetite hormone ghrelin and obestatin levels in the assessment of the severity of acute pancreatitis. Turk. J. Gastroenterol. 2014, 25, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Kellokoski, E.; Kunnari, A.; Jokela, M.; Makela, S.; Kesaniemi, Y.A.; Horkko, S. Ghrelin and obestatin modulate early atherogenic processes on cells: Enhancement of monocyte adhesion and oxidized low-density lipoprotein binding. Metabolism 2009, 58, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yin, X.; Qi, Y.; Pendyala, L.; Chen, J.; Hou, D.; Tang, C. Ghrelin and cardiovascular diseases. Curr. Cardiol. Rev. 2010, 6, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Tokudome, T.; Kishimoto, I. Ghrelin as a treatment for cardiovascular diseases. Hypertension 2014, 64, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virdis, A.; Lerman, L.O.; Regoli, F.; Ghiadoni, L.; Lerman, A.; Taddei, S. Human Ghrelin: A Gastric Hormone with Cardiovascular Properties. Curr. Pharm. Des. 2016, 22, 52–58. [Google Scholar] [CrossRef]
- Katare, R.; Rawal, S.; Munasinghe, P.E.; Tsuchimochi, H.; Inagaki, T.; Fujii, Y.; Dixit, P.; Umetani, K.; Kangawa, K.; Shirai, M.; et al. Ghrelin Promotes Functional Angiogenesis in a Mouse Model of Critical Limb Ischemia Through Activation of Proangiogenic MicroRNAs. Endocrinology 2016, 157, 432–445. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Tang, J.; Yang, T.; Ma, H.; Yi, D.; Gu, C.; Yu, S. Cardioprotective effect of ghrelin in cardiopulmonary bypass involves a reduction in inflammatory response. PLoS ONE 2013, 8, e55021. [Google Scholar] [CrossRef]
- Yuan, M.J.; He, H.; Hu, H.Y.; Li, Q.; Hong, J.; Huang, C.X. Myocardial angiogenesis after chronic ghrelin treatment in a rat myocardial infarction model. Regul. Pept. 2012, 179, 39–42. [Google Scholar] [CrossRef]
- Khazaei, M.; Tahergorabi, Z. Systemic ghrelin administration alters serum biomarkers of angiogenesis in diet-induced obese mice. Int. J. Pept. 2013, 2013, 249565. [Google Scholar] [CrossRef] [Green Version]
- Egido, E.M.; Hernandez, R.; Marco, J.; Silvestre, R.A. Effect of obestatin on insulin, glucagon and somatostatin secretion in the perfused rat pancreas. Regul. Pept. 2009, 152, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Qader, S.S.; Hakanson, R.; Rehfeld, J.F.; Lundquist, I.; Salehi, A. Proghrelin-derived peptides influence the secretion of insulin, glucagon, pancreatic polypeptide and somatostatin: A study on isolated islets from mouse and rat pancreas. Regul. Pept. 2008, 146, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezaki, K.; Hosoda, H.; Kakei, M.; Hashiguchi, S.; Watanabe, M.; Kangawa, K.; Yada, T. Endogenous ghrelin in pancreatic islets restricts insulin release by attenuating Ca2+ signaling in beta-cells: Implication in the glycemic control in rodents. Diabetes 2004, 53, 3142–3151. [Google Scholar] [CrossRef] [Green Version]
- Salehi, A.; Dornonville de la Cour, C.; Hakanson, R.; Lundquist, I. Effects of ghrelin on insulin and glucagon secretion: A study of isolated pancreatic islets and intact mice. Regul. Pept. 2004, 118, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Qader, S.S.; Lundquist, I.; Ekelund, M.; Hakanson, R.; Salehi, A. Ghrelin activates neuronal constitutive nitric oxide synthase in pancreatic islet cells while inhibiting insulin release and stimulating glucagon release. Regul. Pept. 2005, 128, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezaki, K.; Sone, H.; Koizumi, M.; Nakata, M.; Kakei, M.; Nagai, H.; Hosoda, H.; Kangawa, K.; Yada, T. Blockade of pancreatic islet-derived ghrelin enhances insulin secretion to prevent high-fat diet-induced glucose intolerance. Diabetes 2006, 55, 3486–3493. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Asnicar, M.; Saha, P.K.; Chan, L.; Smith, R.G. Ablation of ghrelin improves the diabetic but not obese phenotype of ob/ob mice. Cell Metab. 2006, 3, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Granata, R.; Baragli, A.; Settanni, F.; Scarlatti, F.; Ghigo, E. Unraveling the role of the ghrelin gene peptides in the endocrine pancreas. J. Mol. Endocrinol. 2010, 45, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Doyle, M.E.; Egan, J.M. Mechanisms of action of glucagon-like peptide 1 in the pancreas. Pharmacol. Ther. 2007, 113, 546–593. [Google Scholar] [CrossRef] [Green Version]
- Terauchi, Y.; Takamoto, I.; Kubota, N.; Matsui, J.; Suzuki, R.; Komeda, K.; Hara, A.; Toyoda, Y.; Miwa, I.; Aizawa, S.; et al. Glucokinase and IRS-2 are required for compensatory beta cell hyperplasia in response to high-fat diet-induced insulin resistance. J. Clin. Investig. 2007, 117, 246–257. [Google Scholar] [CrossRef]
- Jhala, U.S.; Canettieri, G.; Screaton, R.A.; Kulkarni, R.N.; Krajewski, S.; Reed, J.; Walker, J.; Lin, X.; White, M.; Montminy, M. cAMP promotes pancreatic beta-cell survival via CREB-mediated induction of IRS2. Genes Dev. 2003, 17, 1575–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukowczan, J.; Warzecha, Z.; Ceranowicz, P.; Kusnierz-Cabala, B.; Tomaszewska, R.; Dembinski, A. Corrigendum to “Pretreatment with obestatin reduces the severity of ischemia/reperfusion-induced acute pancreatitis in rats” [Eur. J. Pharmacol. 760 (2015) 113-121]. Eur. J. Pharmacol. 2019, 842, 384–385. [Google Scholar] [CrossRef]
- Ceranowicz, P.; Warzecha, Z.; Dembinski, A.; Cieszkowski, J.; Dembinski, M.; Sendur, R.; Kusnierz-Cabala, B.; Tomaszewska, R.; Kuwahara, A.; Kato, I. Pretreatment with obestatin inhibits the development of cerulein-induced pancreatitis. J. Physiol. Pharmacol. 2009, 60, 95–101. [Google Scholar] [PubMed]
- Granata, R.; Volante, M.; Settanni, F.; Gauna, C.; Ghe, C.; Annunziata, M.; Deidda, B.; Gesmundo, I.; Abribat, T.; van der Lely, A.J.; et al. Unacylated ghrelin and obestatin increase islet cell mass and prevent diabetes in streptozotocin-treated newborn rats. J. Mol. Endocrinol. 2010, 45, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Irako, T.; Akamizu, T.; Hosoda, H.; Iwakura, H.; Ariyasu, H.; Tojo, K.; Tajima, N.; Kangawa, K. Ghrelin prevents development of diabetes at adult age in streptozotocin-treated newborn rats. Diabetologia 2006, 49, 1264–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baena-Nieto, G.; Lomas-Romero, I.M.; Mateos, R.M.; Leal-Cosme, N.; Perez-Arana, G.; Aguilar-Diosdado, M.; Segundo, C.; Lechuga-Sancho, A.M. Ghrelin mitigates beta-cell mass loss during insulitis in an animal model of autoimmune diabetes mellitus, the BioBreeding/Worcester rat. Diabetes Metab. Res. Rev. 2017, 33. [Google Scholar] [CrossRef] [Green Version]
- DeMarco, V.G.; Sowers, J.R. Ghrelin: A new incretin enhancer therapy? Diabetes 2015, 64, 1500–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reano, S.; Angelino, E.; Ferrara, M.; Malacarne, V.; Sustova, H.; Sabry, O.; Agosti, E.; Clerici, S.; Ruozi, G.; Zentilin, L.; et al. Unacylated Ghrelin Enhances Satellite Cell Function and Relieves the Dystrophic Phenotype in Duchenne Muscular Dystrophy mdx Model. Stem Cells 2017, 35, 1733–1746. [Google Scholar] [CrossRef] [Green Version]
- Gurriaran-Rodriguez, U.; Santos-Zas, I.; Gonzalez-Sanchez, J.; Beiroa, D.; Moresi, V.; Mosteiro, C.S.; Lin, W.; Vinuela, J.E.; Senaris, J.; Garcia-Caballero, T.; et al. Action of obestatin in skeletal muscle repair: Stem cell expansion, muscle growth, and microenvironment remodeling. Mol. Ther. 2015, 23, 1003–1021. [Google Scholar] [CrossRef] [Green Version]
- Ruozi, G.; Bortolotti, F.; Falcione, A.; Dal Ferro, M.; Ukovich, L.; Macedo, A.; Zentilin, L.; Filigheddu, N.; Gortan Cappellari, G.; Baldini, G.; et al. AAV-mediated in vivo functional selection of tissue-protective factors against ischaemia. Nat. Commun. 2015, 6, 7388. [Google Scholar] [CrossRef] [Green Version]
- Santos-Zas, I.; Cid-Diaz, T.; Gonzalez-Sanchez, J.; Gurriaran-Rodriguez, U.; Seoane-Mosteiro, C.; Porteiro, B.; Nogueiras, R.; Casabiell, X.; Luis Relova, J.; Gallego, R.; et al. Obestatin controls skeletal muscle fiber-type determination. Sci. Rep. 2017, 7, 2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontan-Lozano, A.; Saez-Cassanelli, J.L.; Inda, M.C.; de los Santos-Arteaga, M.; Sierra-Dominguez, S.A.; Lopez-Lluch, G.; Delgado-Garcia, J.M.; Carrion, A.M. Caloric restriction increases learning consolidation and facilitates synaptic plasticity through mechanisms dependent on NR2B subunits of the NMDA receptor. J. Neurosci. 2007, 27, 10185–10195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, M.; Yamaki, A.; Furuya, M.; Inomata, N.; Minamitake, Y.; Ohsuye, K.; Kangawa, K. Ghrelin improves body weight loss and skeletal muscle catabolism associated with angiotensin II-induced cachexia in mice. Regul. Pept. 2012, 178, 21–28. [Google Scholar] [CrossRef]
- Nagaya, N.; Moriya, J.; Yasumura, Y.; Uematsu, M.; Ono, F.; Shimizu, W.; Ueno, K.; Kitakaze, M.; Miyatake, K.; Kangawa, K. Effects of ghrelin administration on left ventricular function, exercise capacity, and muscle wasting in patients with chronic heart failure. Circulation 2004, 110, 3674–3679. [Google Scholar] [CrossRef] [Green Version]
- Nagaya, N.; Itoh, T.; Murakami, S.; Oya, H.; Uematsu, M.; Miyatake, K.; Kangawa, K. Treatment of cachexia with ghrelin in patients with COPD. Chest 2005, 128, 1187–1193. [Google Scholar] [CrossRef]
- Balasubramaniam, A.; Joshi, R.; Su, C.; Friend, L.A.; Sheriff, S.; Kagan, R.J.; James, J.H. Ghrelin inhibits skeletal muscle protein breakdown in rats with thermal injury through normalizing elevated expression of E3 ubiquitin ligases MuRF1 and MAFbx. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R893–R901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjögren, M.; Duarte, A.I.; McCourt, A.C.; Shcherbina, L.; Wierup, N.; Björkqvist, M. Ghrelin rescues skeletal muscle catabolic profile in the R6/2 mouse model of Huntington’s disease. Sci. Rep. 2017, 7, 13896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filigheddu, N.; Gnocchi, V.F.; Coscia, M.; Cappelli, M.; Porporato, P.E.; Taulli, R.; Traini, S.; Baldanzi, G.; Chianale, F.; Cutrupi, S.; et al. Ghrelin and des-acyl ghrelin promote differentiation and fusion of C2C12 skeletal muscle cells. Mol. Biol. Cell 2007, 18, 986–994. [Google Scholar] [CrossRef]
- Porporato, P.E.; Filigheddu, N.; Reano, S.; Ferrara, M.; Angelino, E.; Gnocchi, V.F.; Prodam, F.; Ronchi, G.; Fagoonee, S.; Fornaro, M.; et al. Acylated and unacylated ghrelin impair skeletal muscle atrophy in mice. J. Clin. Investig. 2013, 123, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.S.; Wei, Q.; Wang, H.; Kim, D.M.; Balderas, M.; Wu, G.; Lawler, J.; Safe, S.; Guo, S.; Devaraj, S.; et al. Protective Effects of Ghrelin on Fasting-Induced Muscle Atrophy in Aging Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 621–630. [Google Scholar] [CrossRef]
- Angelino, E.; Reano, S.; Bollo, A.; Ferrara, M.; De Feudis, M.; Sustova, H.; Agosti, E.; Clerici, S.; Prodam, F.; Tomasetto, C.-L.; et al. Ghrelin knockout mice display defective skeletal muscle regeneration and impaired satellite cell self-renewal. Endocrine 2018, 62, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Stoyanova, I.I. Ghrelin: A link between ageing, metabolism and neurodegenerative disorders. Neurobiol. Dis. 2014, 72, 72–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diano, S.; Farr, S.A.; Benoit, S.C.; McNay, E.C.; da Silva, I.; Horvath, B.; Gaskin, F.S.; Nonaka, N.; Jaeger, L.B.; Banks, W.A.; et al. Ghrelin controls hippocampal spine synapse density and memory performance. Nat. Neurosci. 2006, 9, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Lacquaniti, A.; Donato, V.; Chirico, V.; Buemi, A.; Buemi, M. Obestatin: An interesting but controversial gut hormone. Ann. Nut. Metab. 2011, 59, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Ku, J.M.; Andrews, Z.B.; Barsby, T.; Reichenbach, A.; Lemus, M.B.; Drummond, G.R.; Sleeman, M.W.; Spencer, S.J.; Sobey, C.G.; Miller, A.A. Ghrelin-related peptides exert protective effects in the cerebral circulation of male mice through a nonclassical ghrelin receptor(s). Endocrinology 2015, 156, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Agnew, A.J.; Robinson, E.; McVicar, C.M.; Harvey, A.P.; Ali, I.H.; Lindsay, J.E.; McDonald, D.M.; Green, B.D.; Grieve, D.J. The gastrointestinal peptide obestatin induces vascular relaxation via specific activation of endothelium-dependent NO signalling. Br. J. Pharmacol. 2012, 166, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyuncuoglu, T.; Vizdiklar, C.; Uren, D.; Yilmaz, H.; Yildirim, C.; Atal, S.S.; Akakin, D.; Kervancioglu Demirci, E.; Yuksel, M.; Yegen, B.C. Obestatin improves oxidative brain damage and memory dysfunction in rats induced with an epileptic seizure. Peptides 2017, 90, 37–47. [Google Scholar] [CrossRef]
- Obay, B.D.; Tasdemir, E.; Tümer, C.; Bilgin, H.M.; Sermet, A. Antiepileptic effects of ghrelin on pentylenetetrazole-induced seizures in rats. Peptides 2007, 28, 1214–1219. [Google Scholar] [CrossRef]
- Weng, M.; Xie, X.; Liu, C.; Lim, K.L.; Zhang, C.W.; Li, L. The Sources of Reactive Oxygen Species and Its Possible Role in the Pathogenesis of Parkinson’s Disease. Parkinsons Dis. 2018, 2018, 9163040. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Song, N.; Xie, J.; Jiang, H. Ghrelin antagonized 1-methyl-4-phenylpyridinium (MPP(+))-induced apoptosis in MES23.5 cells. J. Mol. Neurosci. 2009, 37, 182–189. [Google Scholar] [CrossRef]
- Liu, Y.; Xing, Y.-X.; Gao, X.-Y.; Kuang, H.-Y.; Zhang, J.; Liu, R. Obestatin prevents H2O2-induced damage through activation of TrkB in RGC-5 cells. Biomed. Pharmacother. 2018, 97, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, S.; Ren, J.; Li, B.; Qin, B. Ghrelin protects retinal ganglion cells against rotenone via inhibiting apoptosis, restoring mitochondrial function, and activating AKT-mTOR signaling. Neuropeptides 2018, 67, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Pazos, Y.; Alvarez, C.J.P.; Camiña, J.P.; Casanueva, F.F. Stimulation of extracellular signal-regulated kinases and proliferation in the human gastric cancer cells KATO-III by obestatin. Growth Factors 2007, 25, 373–381. [Google Scholar] [CrossRef]
- Álvarez, C.J.P.; Lodeiro, M.; Theodoropoulou, M.; Camiña, J.P.; Casanueva, F.F.; Pazos, Y. Obestatin stimulates Akt signalling in gastric cancer cells through β-arrestin-mediated epidermal growth factor receptor transactivation. Endocrine Relat. Cancer 2009, 16, 599–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alén, B.O.; Leal-López, S.; Alén, M.O.; Viaño, P.; García-Castro, V.; Mosteiro, C.S.; Beiras, A.; Casanueva, F.F.; Gallego, R.; García-Caballero, T.; et al. The role of the obestatin/GPR39 system in human gastric adenocarcinomas. Oncotarget 2016, 7, 5957–5971. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Liu, Y.P.; Chan, Y.C.; Su, C.Y.; Lin, Y.F.; Hsu, S.L.; Yang, C.S.; Hsiao, M. Ghrelin promotes renal cell carcinoma metastasis via Snail activation and is associated with poor prognosis. J. Pathol. 2015, 237, 50–61. [Google Scholar] [CrossRef]
- Kraus, D.; Reckenbeil, J.; Wenghoefer, M.; Stark, H.; Frentzen, M.; Allam, J.-P.; Novak, N.; Frede, S.; Götz, W.; Probstmeier, R. Ghrelin promotes oral tumor cell proliferation by modifying GLUT1 expression. Cell. Mol. Life Sci. 2016, 73, 1287–1299. [Google Scholar] [CrossRef]
- Majchrzak, K.; Szyszko, K.; Pawłowski, K.; Motyl, T.; Król, M. A role of ghrelin in cancerogenesis. Pol. J. Vet. Sci. 2012, 15, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Majchrzak, K.; Pawłowski, K.M.; Orzechowska, E.J.; Dolka, I.; Mucha, J.; Motyl, T.; Król, M. A role of ghrelin in canine mammary carcinoma cells proliferation, apoptosis and migration. BMC Vet. Res. 2012, 8, 170. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Arai, K.; Sanno, N.; Osamura, R.Y.; Teramoto, A.; Shibasaki, T. Ghrelin and growth hormone (GH) secretagogue receptor (GHSR) mRNA expression in human pituitary adenomas. Clin. Endocrinol. 2001, 54, 759–768. [Google Scholar] [CrossRef]
- Miura, T.; Mitsunaga, S.; Ikeda, M.; Ohno, I.; Takahashi, H.; Suzuki, H.; Irisawa, A.; Kuwata, T.; Ochiai, A. Characterization of low active ghrelin ratio in patients with advanced pancreatic cancer. Support. Care Cancer 2018, 26, 3811–3817. [Google Scholar] [CrossRef] [PubMed]
- Cassoni, P.; Ghe, C.; Marrocco, T.; Tarabra, E.; Allia, E.; Catapano, F.; Deghenghi, R.; Ghigo, E.; Papotti, M.; Muccioli, G. Expression of ghrelin and biological activity of specific receptors for ghrelin and des-acyl ghrelin in human prostate neoplasms and related cell lines. Eur. J. Endocrinol. 2004, 150, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, A.H.; Jeffery, P.L.; Duncan, R.P.; Herington, A.C.; Chopin, L.K. Ghrelin and a novel preproghrelin isoform are highly expressed in prostate cancer and ghrelin activates mitogen-activated protein kinase in prostate cancer. Clin. Cancer Res. 2005, 11, 8295–8303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, P.L.; Herington, A.C.; Chopin, L.K. The potential autocrine/paracrine roles of ghrelin and its receptor in hormone-dependent cancer. Cytokine Growth Factor Rev. 2003, 14, 113–122. [Google Scholar] [CrossRef]
- Hormaechea-Agulla, D.; Gómez-Gómez, E.; Ibáñez-Costa, A.; Carrasco-Valiente, J.; Rivero-Cortés, E.; L-López, F.; Pedraza-Arevalo, S.; Valero-Rosa, J.; Sánchez-Sánchez, R.; Ortega-Salas, R.; et al. Ghrelin O-acyltransferase (GOAT) enzyme is overexpressed in prostate cancer, and its levels are associated with patient’s metabolic status: Potential value as a non-invasive biomarker. Cancer Lett. 2016, 383, 125–134. [Google Scholar] [CrossRef]
- Zhu, J.; Yao, J.; Huang, R.; Wang, Y.; Jia, M.; Huang, Y. Ghrelin promotes human non-small cell lung cancer A549 cell proliferation through PI3K/Akt/mTOR/P70S6K and ERK signaling pathways. Biochem. Biophys. Res. Commun. 2018, 498, 616–620. [Google Scholar] [CrossRef]
- Lien, G.-S.; Lin, C.-H.; Yang, Y.-L.; Wu, M.-S.; Chen, B.-C. Ghrelin induces colon cancer cell proliferation through the GHS-R, Ras, PI3K, Akt, and mTOR signaling pathways. Eur. J. Pharmacol. 2016, 776, 124–131. [Google Scholar] [CrossRef]
- Ławnicka, H.; Mełeń-Mucha, G.; Motylewska, E.; Mucha, S.; Stępień, H. Modulation of ghrelin axis influences the growth of colonic and prostatic cancer cells in vitro. Pharmacol. Rep. 2012, 64, 951–959. [Google Scholar] [CrossRef]
- Yu, H.; Xu, G.; Fan, X. The effect of ghrelin on cell proliferation in small intestinal IEC-6 cells. Biomed. Pharmacother. 2013, 67, 235–239. [Google Scholar] [CrossRef]
- Baldanzi, G.; Filigheddu, N.; Cutrupi, S.; Catapano, F.; Bonissoni, S.; Fubini, A.; Malan, D.; Baj, G.; Granata, R.; Broglio, F.; et al. Ghrelin and des-acyl ghrelin inhibit cell death in cardiomyocytes and endothelial cells through ERK1/2 and PI 3-kinase/AKT. J. Cell Biol. 2002, 159, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Waseem, T.; Duxbury, M.; Ashley, S.W.; Robinson, M.K. Ghrelin promotes intestinal epithelial cell proliferation through PI3K/Akt pathway and EGFR trans-activation both converging to ERK 1/2 phosphorylation. Peptides 2014, 52, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Li, B.B.; Chen, Z.B.; Li, B.C.; Lin, Q.; Li, X.X.; Li, S.L.; Ding, C.; Wu, L.L.; Yu, G.Y. Expression of ghrelin in human salivary glands and its levels in saliva and serum in Chinese obese children and adolescents. Arch. Oral Biol. 2011, 56, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Ozercan, I.H.; Geckil, H.; Dagli, F.; Aydin, S.; Kumru, S.; Kilic, N.; Sahin, I.; Ozercan, M.R. Ghrelin is present in teeth. J. Biochem. Mol. Biol. 2007, 40, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Laborde, N.J.; Kajiya, M.; Shin, J.; Zhu, T.; Thondukolam, A.K.; Min, C.; Kamata, N.; Karimbux, N.Y.; Stashenko, P.; et al. Expression and possible immune-regulatory function of ghrelin in oral epithelium. J. Dent. Res. 2011, 90, 1286–1292. [Google Scholar] [CrossRef] [Green Version]
- Nokhbehsaim, M.; Memmert, S.; Damanaki, A.; Nanayakkara, S.; Zhou, X.; Jager, A.; Deschner, J. Effect of interleukin-1beta on ghrelin receptor in periodontal cells. Clin. Oral Investig. 2019, 23, 113–122. [Google Scholar] [CrossRef]
- Stempniewicz, A.; Ceranowicz, P.; Warzecha, Z. Potential Therapeutic Effects of Gut Hormones, Ghrelin and Obestatin in Oral Mucositis. Int. J. Mol. Sci. 2019, 20, 1534. [Google Scholar] [CrossRef] [Green Version]
- Cieszkowski, J.; Warzecha, Z.; Ceranowicz, P.; Ceranowicz, D.; Kusnierz-Cabala, B.; Pedziwiatr, M.; Dembinski, M.; Ambrozy, T.; Kaczmarzyk, T.; Pihut, M.; et al. Therapeutic effect of exogenous ghrelin in the healing of gingival ulcers is mediated by the release of endogenous growth hormone and insulin-like growth factor-1. J. Physiol. Pharmacol. 2017, 68, 609–617. [Google Scholar]
- Thomas, S.J.; Almers, L.; Schneider, J.; Graham, J.E.; Havel, P.J.; Corley, D.A. Ghrelin and Leptin Have a Complex Relationship with Risk of Barrett’s Esophagus. Dig. Dis. Sci. 2016, 61, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Takata, A.; Takiguchi, S.; Miyazaki, Y.; Miyata, H.; Takahashi, T.; Kurokawa, Y.; Yamasaki, M.; Nakajima, K.; Mori, M.; Kangawa, K.; et al. Randomized Phase II Study of the Anti-inflammatory Effect of Ghrelin During the Postoperative Period of Esophagectomy. Ann. Surg. 2015, 262, 230–236. [Google Scholar] [CrossRef]
- Yamamoto, K.; Takiguchi, S.; Miyata, H.; Miyazaki, Y.; Hiura, Y.; Yamasaki, M.; Nakajima, K.; Fujiwara, Y.; Mori, M.; Kangawa, K.; et al. Reduced plasma ghrelin levels on day 1 after esophagectomy: A new predictor of prolonged systemic inflammatory response syndrome. Surg. Today 2013, 43, 48–54. [Google Scholar] [CrossRef]
- Sibilia, V.; Rindi, G.; Pagani, F.; Rapetti, D.; Locatelli, V.; Torsello, A.; Campanini, N.; Deghenghi, R.; Netti, C. Ghrelin protects against ethanol-induced gastric ulcers in rats: Studies on the mechanisms of action. Endocrinology 2003, 144, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konturek, P.C.; Brzozowski, T.; Pajdo, R.; Nikiforuk, A.; Kwiecien, S.; Harsch, I.; Drozdowicz, D.; Hahn, E.G.; Konturek, S.J. Ghrelin-a new gastroprotective factor in gastric mucosa. J. Physiol. Pharmacol. 2004, 55, 325–336. [Google Scholar] [PubMed]
- Ginter, G.; Ceranowicz, P.; Warzecha, Z. Protective and Healing Effects of Ghrelin and Risk of Cancer in the Digestive System. Int. J. Mol. Sci. 2021, 22, 10571. [Google Scholar] [CrossRef] [PubMed]
- Warzecha, Z.; Ceranowicz, P.; Dembinski, M.; Cieszkowski, J.; Ginter, G.; Ptak-Belowska, A.; Dembinski, A. Involvement of cyclooxygenase-1 and cyclooxygenase-2 activity in the therapeutic effect of ghrelin in the course of ethanol-induced gastric ulcers in rats. J. Physiol. Pharmacol. 2014, 65, 95–106. [Google Scholar] [PubMed]
- Ceranowicz, P.; Warzecha, Z.; Dembinski, A.; Sendur, R.; Cieszkowski, J.; Ceranowicz, D.; Pawlik, W.W.; Kuwahara, A.; Kato, I.; Konturek, P.C. Treatment with ghrelin accelerates the healing of acetic acid-induced gastric and duodenal ulcers in rats. J. Physiol. Pharmacol. 2009, 60, 87–98. [Google Scholar] [PubMed]
- Wu, R.; Dong, W.; Ji, Y.; Zhou, M.; Marini, C.P.; Ravikumar, T.S.; Wang, P. Orexigenic hormone ghrelin attenuates local and remote organ injury after intestinal ischemia-reperfusion. PLoS ONE 2008, 3, e2026. [Google Scholar] [CrossRef]
- Pawlik, M.W.; Obuchowicz, R.; Biernat, J.; Szczepanski, W.; Pajdo, R.; Kwiecien, S.; Brzozowski, T.; Konturek, S.J.; Pawlik, W.W. Effects of peripherally and centrally applied ghrelin in the pathogenesis of ischemia-reperfusion induced injury of the small intestine. J. Physiol. Pharmacol. 2011, 62, 429–439. [Google Scholar]
- Wang, Z.; Yang, W.L.; Jacob, A.; Aziz, M.; Wang, P. Human ghrelin mitigates intestinal injury and mortality after whole body irradiation in rats. PLoS ONE 2015, 10, e0118213. [Google Scholar] [CrossRef]
- Cheng, Y.; Wei, Y.; Yang, W.; Cai, Y.; Chen, B.; Yang, G.; Shang, H.; Zhao, W. Ghrelin Attenuates Intestinal Barrier Dysfunction Following Intracerebral Hemorrhage in Mice. Int. J. Mol. Sci. 2016, 17, 2032. [Google Scholar] [CrossRef] [Green Version]
- Warzecha, Z.; Ceranowicz, D.; Dembinski, A.; Ceranowicz, P.; Cieszkowski, J.; Kuwahara, A.; Kato, I.; Dembinski, M.; Konturek, P.C. Ghrelin accelerates the healing of cysteamine-induced duodenal ulcers in rats. Med. Sci. Monit. 2012, 18, BR181–BR187. [Google Scholar] [CrossRef]
- Onishi, S.; Kaji, T.; Yamada, W.; Nakame, K.; Machigashira, S.; Kawano, M.; Yano, K.; Harumatsu, T.; Yamada, K.; Masuya, R.; et al. Ghrelin stimulates intestinal adaptation following massive small bowel resection in parenterally fed rats. Peptides 2018, 106, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Sanchez, N.; Ponciano-Rodriguez, G.; Bermejo-Martinez, L.; Villa, A.R.; Chavez-Tapia, N.C.; Zamora-Valdes, D.; Pichardo-Bahena, R.; Barredo-Prieto, B.; Uribe-Ramos, M.H.; Ramos, M.H.; et al. Low serum levels of ghrelin are associated with gallstone disease. World J. Gastroenterol. 2006, 12, 3096–3100. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Grobe, Y.; Villalobos-Blasquez, I.; Sanchez-Lara, K.; Villa, A.R.; Ponciano-Rodriguez, G.; Ramos, M.H.; Chavez-Tapia, N.C.; Uribe, M.; Mendez-Sanchez, N. High ghrelin and obestatin levels and low risk of developing fatty liver. Ann. Hepatol. 2010, 9, 52–57. [Google Scholar] [CrossRef]
- Ezquerro, S.; Mocha, F.; Fruhbeck, G.; Guzman-Ruiz, R.; Valenti, V.; Mugueta, C.; Becerril, S.; Catalan, V.; Gomez-Ambrosi, J.; Silva, C.; et al. Ghrelin Reduces TNF-alpha-Induced Human Hepatocyte Apoptosis, Autophagy, and Pyroptosis: Role in Obesity-Associated NAFLD. J. Clin. Endocrinol. Metab. 2019, 104, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Nagoya, T.; Kamimura, K.; Inoue, R.; Ko, M.; Owaki, T.; Niwa, Y.; Sakai, N.; Setsu, T.; Sakamaki, A.; Yokoo, T.; et al. Ghrelin-insulin-like growth factor-1 axis is activated via autonomic neural circuits in the non-alcoholic fatty liver disease. Neurogastroenterol. Motil. 2020, 32, e13799. [Google Scholar] [CrossRef]
- Li, Y.; Hai, J.; Li, L.; Chen, X.; Peng, H.; Cao, M.; Zhang, Q. Administration of ghrelin improves inflammation, oxidative stress, and apoptosis during and after non-alcoholic fatty liver disease development. Endocrine 2013, 43, 376–386. [Google Scholar] [CrossRef]
- Dembinski, A.; Warzecha, Z.; Ceranowicz, P.; Tomaszewska, R.; Stachura, J.; Konturek, S.J.; Konturek, P.C. Ghrelin attenuates the development of acute pancreatitis in rat. J. Physiol. Pharmacol. 2003, 54, 561–573. [Google Scholar]
- Dembinski, A.; Warzecha, Z.; Ceranowicz, P.; Cieszkowski, J.; Pawlik, W.W.; Tomaszewska, R.; Kusnierz-Cabala, B.; Naskalski, J.W.; Kuwahara, A.; Kato, I. Role of growth hormone and insulin-like growth factor-1 in the protective effect of ghrelin in ischemia/reperfusion-induced acute pancreatitis. Growth Horm. IGF Res. 2006, 16, 348–356. [Google Scholar] [CrossRef]
- Zhou, X.; Xue, C. Ghrelin inhibits the development of acute pancreatitis and nuclear factor kappaB activation in pancreas and liver. Pancreas 2009, 38, 752–757. [Google Scholar] [CrossRef]
- Warzecha, Z.; Ceranowicz, P.; Dembinski, A.; Cieszkowski, J.; Kusnierz-Cabala, B.; Tomaszewska, R.; Kuwahara, A.; Kato, I. Therapeutic effect of ghrelin in the course of cerulein-induced acute pancreatitis in rats. J. Physiol. Pharmacol. 2010, 61, 419–427. [Google Scholar]
- Bukowczan, J.; Warzecha, Z.; Ceranowicz, P.; Kusnierz-Cabala, B.; Tomaszewska, R.; Dembinski, A. Therapeutic effect of ghrelin in the course of ischemia/reperfusion-induced acute pancreatitis. Curr. Pharm. Des. 2015, 21, 2284–2290. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, S.; Oshitani, N.; Kamata, N.; Sogawa, M.; Yamagami, H.; Watanabe, K.; Tominaga, K.; Watanabe, T.; Fujiwara, Y.; Maeda, K.; et al. Phenotypical and functional study of ghrelin and its receptor in the pathogenesis of Crohn’s disease. Inflamm. Bowel Dis. 2008, 14, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Engel, M.; Burnat, G.; Gaca, P.; Kwiecien, S.; Pajdo, R.; Konturek, S.J. Ghrelin ameliorates colonic inflammation. Role of nitric oxide and sensory nerves. J. Physiol. Pharmacol. 2009, 60, 41–47. [Google Scholar] [PubMed]
- Gonzalez-Rey, E.; Chorny, A.; Delgado, M. Therapeutic action of ghrelin in a mouse model of colitis. Gastroenterology 2006, 130, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Matuszyk, A.; Ceranowicz, P.; Warzecha, Z.; Cieszkowski, J.; Ceranowicz, D.; Galazka, K.; Bonior, J.; Jaworek, J.; Bartus, K.; Gil, K.; et al. Exogenous Ghrelin Accelerates the Healing of Acetic Acid-Induced Colitis in Rats. Int. J. Mol. Sci. 2016, 17, 1455. [Google Scholar] [CrossRef] [Green Version]
- Ceranowicz, P.; Warzecha, Z.; Cieszkowski, J.; Ceranowicz, D.; Kusnierz-Cabala, B.; Bonior, J.; Jaworek, J.; Ambrozy, T.; Gil, K.; Olszanecki, R.; et al. Essential Role of Growth Hormone and IGF-1 in Therapeutic Effect of Ghrelin in the Course of Acetic Acid-Induced Colitis. Int. J. Mol. Sci. 2017, 18, 1118. [Google Scholar] [CrossRef] [Green Version]
- Noh, J.Y.; Wu, C.S.; DeLuca, J.A.A.; Devaraj, S.; Jayaraman, A.; Alaniz, R.C.; Tan, X.D.; Allred, C.D.; Sun, Y. Novel Role of Ghrelin Receptor in Gut Dysbiosis and Experimental Colitis in Aging. Int. J. Mol. Sci. 2022, 23, 2219. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, J.; Shen, J.; Wang, S.; Guo, C.; Fan, X. Ghrelin Inhibits Intestinal Epithelial Cell Apoptosis Through the Unfolded Protein Response Pathway in Ulcerative Colitis. Front. Pharmacol. 2021, 12, 661853. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, L.; Dai, W.; Mao, Y.; Li, S.; Wang, J.; Li, H.; Guo, C.; Fan, X. Ghrelin ameliorates intestinal barrier dysfunction in experimental colitis by inhibiting the activation of nuclear factor-kappa B. Biochem. Biophys. Res. Commun. 2015, 458, 140–147. [Google Scholar] [CrossRef]
- De Smet, B.; Thijs, T.; Moechars, D.; Colsoul, B.; Polders, L.; Ver Donck, L.; Coulie, B.; Peeters, T.L.; Depoortere, I. Endogenous and exogenous ghrelin enhance the colonic and gastric manifestations of dextran sodium sulphate-induced colitis in mice. Neurogastroenterol. Motil. 2009, 21, 59–70. [Google Scholar] [CrossRef]
- Liu, Z.Z.; Wang, W.G.; Li, Q.; Tang, M.; Li, J.; Wu, W.T.; Wan, Y.H.; Wang, Z.G.; Bao, S.S.; Fei, J. Growth hormone secretagogue receptor is important in the development of experimental colitis. Cell Biosci. 2015, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, P.; Lu, X.; Jin, N.; Shi, J. Knockdown of ghrelin-O-acyltransferase attenuates colitis through the modulation of inflammatory factors and tight junction proteins in the intestinal epithelium. Cell. Biol. Int. 2020, 44, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Mathur, N.; Mehdi, S.F.; Anipindi, M.; Aziz, M.; Khan, S.A.; Kondakindi, H.; Lowell, B.; Wang, P.; Roth, J. Ghrelin as an Anti-Sepsis Peptide: Review. Front. Immunol. 2020, 11, 610363. [Google Scholar] [CrossRef] [PubMed]
- Waseem, T.; Duxbury, M.; Ito, H.; Ashley, S.W.; Robinson, M.K. Exogenous ghrelin modulates release of pro-inflammatory and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Surgery 2008, 143, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Jacob, A.; Rajan, D.; Pathickal, B.; Balouch, I.; Hartman, A.; Wu, R.; Zhou, M.; Wang, P. The inhibitory effect of ghrelin on sepsis-induced inflammation is mediated by the MAPK phosphatase-1. Int. J. Mol. Med. 2010, 25, 159–164. [Google Scholar]
- Dembinski, A.; Warzecha, Z.; Ceranowicz, P.; Cieszkowski, J.; Dembinski, M.; Ptak-Belowska, A.; Kuwahara, A.; Kato, I. Administration of obestatin accelerates the healing of chronic gastric ulcers in rats. Med. Sci. Monit. 2011, 17, BR196–BR200. [Google Scholar] [CrossRef] [Green Version]
- Matuszyk, A.; Ceranowicz, P.; Warzecha, Z.; Cieszkowski, J.; Galazka, K.; Bonior, J.; Jaworek, J.; Konturek, P.C.; Gil, K.; Dembinski, A. Pretreatment with obestatin inhibits the development of acetic acid-induced colitis in rats. Arch. Med. Sci. 2018, 14, 920–929. [Google Scholar] [CrossRef] [Green Version]
- Matuszyk, A.; Ceranowicz, P.; Warzecha, Z.; Cieszkowski, J.; Bonior, J.; Jaworek, J.; Kusnierz-Cabala, B.; Konturek, P.; Ambrozy, T.; Dembinski, A. Obestatin Accelerates the Healing of Acetic Acid-Induced Colitis in Rats. Oxid. Med. Cell. Longev. 2016, 2016, 2834386. [Google Scholar] [CrossRef] [Green Version]
- Konarska, K.; Cieszkowski, J.; Warzecha, Z.; Ceranowicz, P.; Chmura, A.; Kusnierz-Cabala, B.; Galazka, K.; Kowalczyk, P.; Miskiewicz, A.; Konturek, T.J.; et al. Treatment with Obestatin-A Ghrelin Gene-Encoded Peptide-Reduces the Severity of Experimental Colitis Evoked by Trinitrobenzene Sulfonic Acid. Int. J. Mol. Sci. 2018, 19, 1643. [Google Scholar] [CrossRef] [Green Version]
- Ulasoglu, C.; Isbilen, B.; Doganay, L.; Ozen, F.; Kiziltas, S.; Tuncer, I. Effect of Helicobacter pylori eradication on serum ghrelin and obestatin levels. World J. Gastroenterol. 2013, 19, 2388–2394. [Google Scholar] [CrossRef]
- Bukowczan, J.; Warzecha, Z.; Ceranowicz, P.; Kusnierz-Cabala, B.; Tomaszewska, R.; Dembinski, A. Pretreatment with obestatin reduces the severity of ischemia/reperfusion-induced acute pancreatitis in rats. Eur. J. Pharmacol. 2015, 760, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Bukowczan, J.; Cieszkowski, J.; Warzecha, Z.; Ceranowicz, P.; Kusnierz-Cabala, B.; Tomaszewska, R.; Dembinski, A. Therapeutic Effect of Obestatin in the Course of Cerulein-Induced Acute Pancreatitis. Pancreas 2016, 45, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Bukowczan, J.; Warzecha, Z.; Ceranowicz, P.; Kusnierz-Cabala, B.; Tomaszewska, R. Obestatin Accelerates the Recovery in the Course of Ischemia/Reperfusion-Induced Acute Pancreatitis in Rats. PLoS ONE 2015, 10, e0134380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gohary, O.A. Obestatin improves hepatic injury induced by ischemia/reperfusion in rats: Role of nitric oxide. Gen. Physiol. Biophys. 2017, 36, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Granata, R.; Gallo, D.; Luque, R.M.; Baragli, A.; Scarlatti, F.; Grande, C.; Gesmundo, I.; Cordoba-Chacon, J.; Bergandi, L.; Settanni, F.; et al. Obestatin regulates adipocyte function and protects against diet-induced insulin resistance and inflammation. FASEB J. 2012, 26, 3393–3411. [Google Scholar] [CrossRef] [Green Version]
- Tschop, M.; Smiley, D.L.; Heiman, M.L. Ghrelin induces adiposity in rodents. Nature 2000, 407, 908–913. [Google Scholar] [CrossRef]
- Theander-Carrillo, C.; Wiedmer, P.; Cettour-Rose, P.; Nogueiras, R.; Perez-Tilve, D.; Pfluger, P.; Castaneda, T.R.; Muzzin, P.; Schurmann, A.; Szanto, I.; et al. Ghrelin action in the brain controls adipocyte metabolism. J. Clin. Investig. 2006, 116, 1983–1993. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Roh, S.G.; Hong, Y.H.; Shrestha, Y.B.; Hishikawa, D.; Chen, C.; Kojima, M.; Kangawa, K.; Sasaki, S. The role of ghrelin and growth hormone secretagogues receptor on rat adipogenesis. Endocrinology 2003, 144, 754–759. [Google Scholar] [CrossRef] [Green Version]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.H.; Doria, A.; et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Kozak, L.P. Brown fat and the myth of diet-induced thermogenesis. Cell Metab. 2010, 11, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerback, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.I.; Middelbeek, R.J.; Townsend, K.L.; An, D.; Nygaard, E.B.; Hitchcox, K.M.; Markan, K.R.; Nakano, K.; Hirshman, M.F.; Tseng, Y.H.; et al. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity. J. Clin. Investig. 2013, 123, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Chondronikola, M.; Volpi, E.; Borsheim, E.; Porter, C.; Annamalai, P.; Enerback, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M.; et al. Brown adipose tissue improves whole-body glucose homeostasis and insulin sensitivity in humans. Diabetes 2014, 63, 4089–4099. [Google Scholar] [CrossRef] [Green Version]
- Mano-Otagiri, A.; Iwasaki-Sekino, A.; Nemoto, T.; Ohata, H.; Shuto, Y.; Nakabayashi, H.; Sugihara, H.; Oikawa, S.; Shibasaki, T. Genetic suppression of ghrelin receptors activates brown adipocyte function and decreases fat storage in rats. Regul. Pept. 2010, 160, 81–90. [Google Scholar] [CrossRef]
- Mano-Otagiri, A.; Ohata, H.; Iwasaki-Sekino, A.; Nemoto, T.; Shibasaki, T. Ghrelin suppresses noradrenaline release in the brown adipose tissue of rats. J. Endocrinol. 2009, 201, 341–349. [Google Scholar] [CrossRef]
- Lin, L.; Saha, P.K.; Ma, X.; Henshaw, I.O.; Shao, L.; Chang, B.H.; Buras, E.D.; Tong, Q.; Chan, L.; McGuinness, O.P.; et al. Ablation of ghrelin receptor reduces adiposity and improves insulin sensitivity during aging by regulating fat metabolism in white and brown adipose tissues. Aging Cell 2011, 10, 996–1010. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, T.; Masaki, T.; Kakuma, T.; Yoshimatsu, H. Centrally administered ghrelin suppresses sympathetic nerve activity in brown adipose tissue of rats. Neurosci. Lett. 2003, 349, 75–78. [Google Scholar] [CrossRef]
- Tsubone, T.; Masaki, T.; Katsuragi, I.; Tanaka, K.; Kakuma, T.; Yoshimatsu, H. Ghrelin regulates adiposity in white adipose tissue and UCP1 mRNA expression in brown adipose tissue in mice. Regul. Pept. 2005, 130, 97–103. [Google Scholar] [CrossRef]
- Ma, X.; Lin, L.; Qin, G.; Lu, X.; Fiorotto, M.; Dixit, V.D.; Sun, Y. Ablations of ghrelin and ghrelin receptor exhibit differential metabolic phenotypes and thermogenic capacity during aging. PLoS ONE 2011, 6, e16391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szentirmai, E.; Hajdu, I.; Obal, F., Jr.; Krueger, J.M. Ghrelin-induced sleep responses in ad libitum fed and food-restricted rats. Brain Res. 2006, 1088, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Szentirmai, E.; Krueger, J.M. Obestatin alters sleep in rats. Neurosci. Lett. 2006, 404, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Szentirmai, E.; Kapas, L.; Sun, Y.; Smith, R.G.; Krueger, J.M. The preproghrelin gene is required for the normal integration of thermoregulation and sleep in mice. Proc. Natl. Acad. Sci. USA 2009, 106, 14069–14074. [Google Scholar] [CrossRef] [Green Version]
- Abel, T.; Havekes, R.; Saletin, J.M.; Walker, M.P. Sleep, Plasticity and Memory from Molecules to Whole-Brain Networks. Curr. Biol. 2013, 23, R774–R788. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Acylated Ghrelin (AG) | Obestatin | |
|---|---|---|
| (1) Food intake | GH release via GHSR activation [12,13,14] | Down-regulation of NPY and NPY-R [37,42] |
| (2) Insulin signaling | AMPK signaling mediated G-protein subunit activation [101] | Regulation of pScr, Akt, ERK1/2 via GLP-1R binding [27] |
| (3) Thermogenesis | Suppression of NE release of BAT [170,171] | Up-regulation of UCP1 mRNA expression [8] |
| (4) Neuronal Function | Neuronal survival of epileptic mice via attenuation of lipid peroxidation [134] | Attenuation of epileptic seizure duration/onset via interaction with neuropeptide Y (NPY) or GABA neurons [135] |
| (5) Muscle function | Activation of GH/IGF-1 via mTOR/Akt signaling [120,126] | Regulation of myogenic differentiation through involvement of GPR39 [23] |
| (6) Cardiovascular function | Cardiac pumping function via GHSR and Akt signaling [95] | Activation of PI3K, PKCs and ERK1/2 pathway [82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villarreal, D.; Pradhan, G.; Zhou, Y.; Xue, B.; Sun, Y. Diverse and Complementary Effects of Ghrelin and Obestatin. Biomolecules 2022, 12, 517. https://doi.org/10.3390/biom12040517
Villarreal D, Pradhan G, Zhou Y, Xue B, Sun Y. Diverse and Complementary Effects of Ghrelin and Obestatin. Biomolecules. 2022; 12(4):517. https://doi.org/10.3390/biom12040517
Chicago/Turabian StyleVillarreal, Daniel, Geetali Pradhan, Yu Zhou, Bingzhong Xue, and Yuxiang Sun. 2022. "Diverse and Complementary Effects of Ghrelin and Obestatin" Biomolecules 12, no. 4: 517. https://doi.org/10.3390/biom12040517
APA StyleVillarreal, D., Pradhan, G., Zhou, Y., Xue, B., & Sun, Y. (2022). Diverse and Complementary Effects of Ghrelin and Obestatin. Biomolecules, 12(4), 517. https://doi.org/10.3390/biom12040517