
Biochemical Properties and Physiological Functions of pLG72: Twenty Years of Investigations




Abstract
:1. Introduction
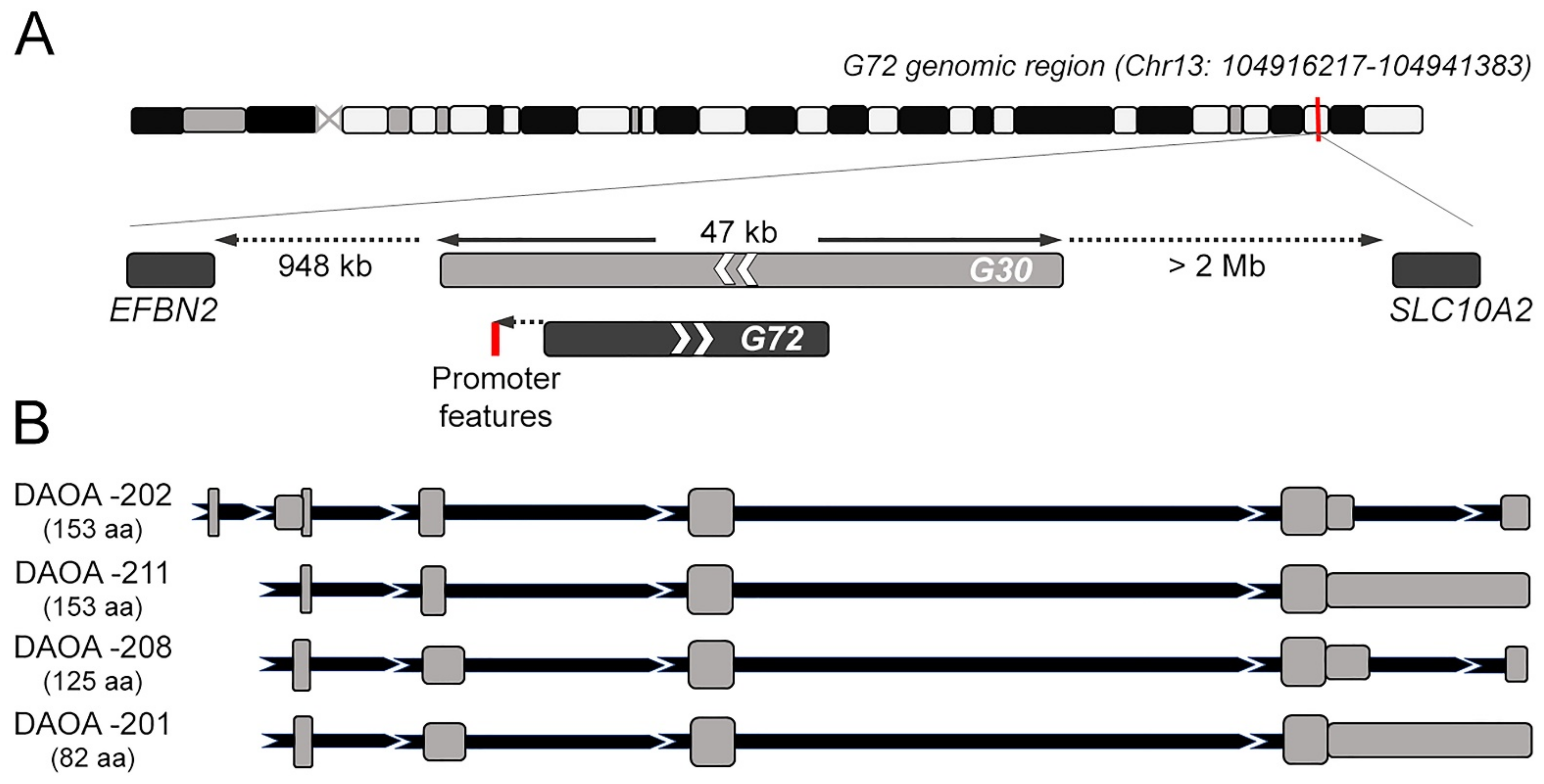
2. G72/G30 Locus and Transcripts
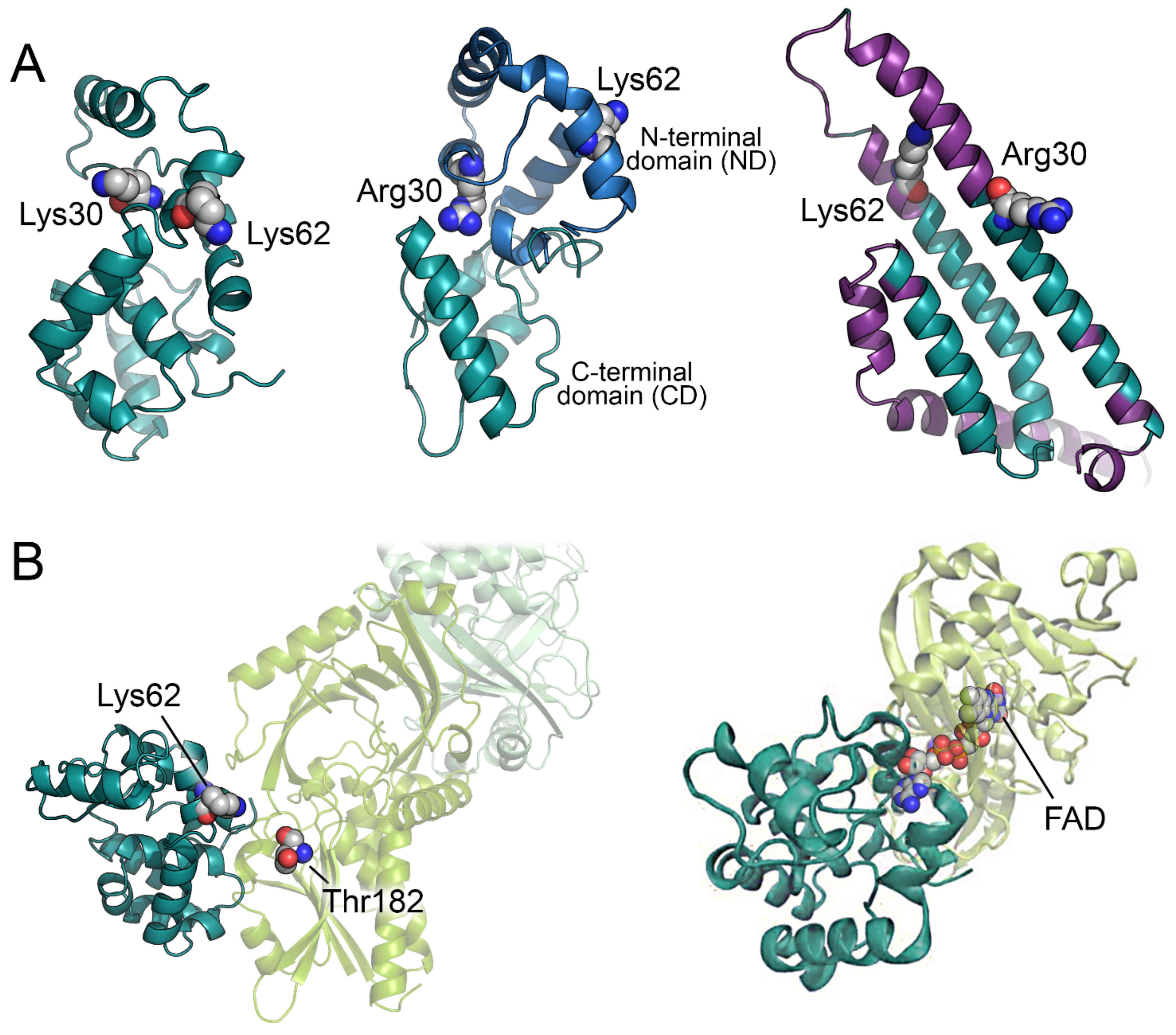
3. Biochemical Properties
4. pLG72 Interactors
5. Putative Biological Roles of pLG72
6. pLG72 Levels in Human Brain
7. Blood pLG72 Levels: A Novel Biomarker?
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chumakov, I.; Blumenfeld, M.; Guerassimenko, O.; Cavarec, L.; Palicio, M.; Abderrahim, H.; Bougueleret, L.; Barry, C.; Tanaka, H.; La Rosa, P.; et al. Genetic and physiological data implicating the new human gene G72 and the gene for D-amino acid oxidase in schizophrenia. Proc. Natl. Acad. Sci. USA 2002, 99, 13675–13680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacchi, S.; Caldinelli, L.; Cappelletti, P.; Pollegioni, L.; Molla, G. Structure–function relationships in human D-amino acid oxidase. Amino Acids 2012, 43, 1833–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollegioni, L.; Sacchi, S. Metabolism of the neuromodulator D-serine. Cell Mol. Life Sci. 2010, 67, 2387–2404. [Google Scholar] [CrossRef] [PubMed]
- Mothet, J.P.; Parent, A.T.; Wolosker, H.; Brady, R.O., Jr.; Linden, D.J.; Ferris, C.D.; Rogawski, M.A.; Snyder, S.H. D-serine is an endogenous ligand for the glycine site of the N-methyl-D-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panatier, A.; Theodosis, D.T.; Mothet, J.P.; Touquet, B.; Pollegioni, L.; Poulain, D.A.; Oliet, S.H. Glia-derived D-serine controls NMDA receptor activity and synaptic memory. Cell 2006, 125, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Papouin, T.; Ladépêche, L.; Ruel, J.; Sacchi, S.; Labasque, M.; Hanini, M.; Groc, L.; Pollegioni, L.; Mothet, J.P.; Oliet, S.H. Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell 2012, 150, 633–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addington, A.M.; Gornick, M.; Sporn, A.L.; Gogtay, N.; Greenstein, D.; Lenane, M.; Gochman, P.; Baker, N.; Balkissoon, R.; Vakkalanka, R.K.; et al. Polymorphisms in the 13q33.2 gene G72/G30 are associated with childhood-onset schizophrenia and psychosis not otherwise specified. Biol. Psychiatry 2004, 55, 976–980. [Google Scholar] [CrossRef]
- Schumacher, J.; Jamra, R.A.; Freudenberg, J.; Becker, T.; Ohlraun, S.; Otte, A.C.; Tullius, M.; Kovalenko, S.; Bogaert, A.V.; Maier, W.; et al. Examination of G72 and D-amino-acid oxidase as genetic risk factors for schizophrenia and bipolar affective disorder. Mol. Psychiatry 2004, 9, 203–207. [Google Scholar] [CrossRef]
- Korostishevsky, M.; Kremer, I.; Kaganovich, M.; Cholostoy, A.; Murad, I.; Muhaheed, M.; Bannoura, I.; Rietschel, M.; Dobrusin, M.; Bening-Abu-Shach, U.; et al. Transmission disequilibrium and haplotype analyses of the G72/G30 locus: Suggestive linkage to schizophrenia in Palestinian Arabs living in the North of Israel. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141B, 91–95. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Zhao, Q.; Huang, K.; Wang, P.; Yang, P.; Li, S.; Feng, G.; Lindpaintner, K.; He, L.; et al. First evidence of association between G72 and bipolar disorder in the Chinese Han population. Psychiatr. Genet. 2009, 19, 151–153. [Google Scholar] [CrossRef]
- Rietschel, M.; Beckmann, L.; Strohmaier, J.; Georgi, A.; Karpushova, A.; Schirmbeck, F.; Boesshenz, K.V.; Schmäl, C.; Bürger, C.; Jamra, R.A.; et al. G72 and its association with major depression and neuroticism in large population-based groups from Germany. Am. J. Psychiatry 2008, 165, 753–762. [Google Scholar] [CrossRef]
- Jansen, A.; Krach, S.; Krug, A.; Markov, V.; Eggermann, T.; Zerres, K.; Stöcker, T.; Shah, N.J.; Nöthen, M.M.; Treutlein, J.; et al. A putative high risk diplotype of the G72 gene is in healthy individuals associated with better performance in working memory functions and altered brain activity in the medial temporal lobe. Neuroimage 2009, 45, 1002–1008. [Google Scholar] [CrossRef]
- Ishiwata, S.; Hattori, K.; Sasayama, D.; Teraishi, T.; Miyakawa, T.; Yokota, Y.; Matsumura, R.; Yoshida, F.; Nishikawa, T.; Kunugi, H. Plasma and cerebrospinal fluid G72 protein levels in schizophrenia and major depressive disorder. Psychiatry Res. 2017, 254, 244–250. [Google Scholar] [CrossRef]
- Sacchi, S.; Binelli, G.; Pollegioni, L. G72 primate-specific gene: A still enigmatic element in psychiatric disorders. Cell Mol. Life Sci. 2016, 73, 2029–2039. [Google Scholar] [CrossRef]
- Detera-Wadleigh, S.D.; McMahon, F.J. G72/G30 in schizophrenia and bipolar disorder: Review and meta-analysis. Biol. Psychiatry 2006, 60, 106–114. [Google Scholar] [CrossRef]
- Li, D.; He, L. G72/G30 genes and schizophrenia: A systematic meta-analysis of association studies. Genetics 2007, 175, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Owen, M.J.; Craddock, N.; Jablensky, A. The genetic deconstruction of psychosis. Schizophr. Bull. 2007, 33, 905–911. [Google Scholar] [CrossRef] [Green Version]
- Müller, D.J.; Zai, C.C.; Shinkai, T.; Strauss, J.; Kennedy, J.L. Association between the DAOA/G72 gene and bipolar disorder and meta-analyses in bipolar disorder and schizophrenia. Bipolar Disord. 2011, 13, 198–207. [Google Scholar] [CrossRef]
- Sacchi, S.; Cappelletti, P.; Pirone, L.; Smaldone, G.; Pedone, E.; Pollegioni, L. Elucidating the role of the pLG72 R30K substitution in schizophrenia susceptibility. FEBS Lett. 2017, 591, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Hartz, S.M.; Ho, B.C.; Andreasen, N.C.; Librant, A.; Rudd, D.; Epping, E.A.; Wassink, T.H. G72 influences longitudinal change in frontal lobe volume in schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153B, 640–647. [Google Scholar] [CrossRef] [Green Version]
- Schultz, C.C.; Nenadic, I.; Koch, K.; Wagner, G.; Roebel, M.; Schachtzabel, C.; Mühleisen, T.W.; Nöthen, M.M.; Cichon, S.; Deufel, T.; et al. Reduced cortical thickness is associated with the glutamatergic regulatory gene risk variant DAOA Arg30Lys in schizophrenia. Neuropsychopharmacology 2011, 36, 1747–1753. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, Y.; Zhang, J.; Liu, Z.; Xu, C.; Zhang, K.; Shen, Y.; Xu, Q. A combined study of genetic association and brain imaging on the DAOA gene in schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2013, 162B, 191–200. [Google Scholar] [CrossRef]
- Su, W.; Zhu, T.; Xu, L.; Wei, Y.; Zeng, B.; Zhang, T.; Cui, H.; Wang, J.; Jia, Y.; Wang, J.; et al. Effect of DAOA genetic variation on white matter alteration in corpus callosum in patients with first-episode schizophrenia. Brain Imaging Behav. 2021, 15, 1748–1759. [Google Scholar] [CrossRef]
- Donohoe, G.; Morris, D.W.; Robertson, I.H.; McGhee, K.A.; Murphy, K.; Kenny, N.; Clarke, S.; Gill, M.; Corvin, A.P. DAOA Arg30Lys and verbal memory function in schizophrenia. Mol. Psychiatry 2007, 12, 795–796. [Google Scholar] [CrossRef]
- Soronen, P.; Silander, K.; Antila, M.; Palo, O.M.; Tuulio-Henriksson, A.; Kieseppä, T.; Ellonen, P.; Wedenoja, J.; Turunen, J.A.; Pietiläinen, O.P.; et al. Association of a nonsynonymous variant of DAOA with visuospatial ability in a bipolar family sample. Biol. Psychiatry 2008, 64, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Britten, R. Transposable elements have contributed to thousands of human proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 1798–1803. [Google Scholar] [CrossRef] [Green Version]
- Benzel, I.; Kew, J.N.; Viknaraja, R.; Kelly, F.; de Belleroche, J.; Hirsch, S.; Sanderson, T.H.; Maycox, P.R. Investigation of G72 (DAOA) expression in the human brain. BMC Psychiatry 2008, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, E.; Liu, C.; Badner, J.A.; Bonner, T.I.; Christian, S.L.; Maheshwari, M.; Detera-Wadleigh, S.D.; Gibbs, R.A.; Gershon, E.S. Polymorphisms at the G72/G30 gene locus, on 13q33, are associated with bipolar disorder in two independent pedigree series. Am. J. Hum. Genet. 2003, 72, 1131–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molla, G.; Bernasconi, M.; Sacchi, S.; Pilone, M.S.; Pollegioni, L. Expression in Escherichia coli and in vitro refolding of the human protein pLG72. Protein Expr. Purif. 2006, 46, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Birolo, L.; Sacchi, S.; Smaldone, G.; Molla, G.; Leo, G.; Caldinelli, L.; Pirone, L.; Eliometri, P.; Di Gaetano, S.; Orefice, I.; et al. Regulating levels of the neuromodulator d-serine in human brain: Structural insight into pLG72 and D-amino acid oxidase interaction. FEBS J. 2016, 283, 3353–3370. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Fukui, K. Structure models of G72, the product of a susceptibility gene to schizophrenia. J. Biochem. 2017, 161, 223–230. [Google Scholar] [CrossRef]
- Jagannath, V.; Brotzakis, Z.F.; Parrinello, M.; Walitza, S.; Grünblatt, E. Controversial effects of D-amino acid oxidase activator (DAOA)/G72 on D-amino acid oxidase (DAO) activity in human neuronal, astrocyte and kidney cell lines: The N-methyl D-aspartate (NMDA) receptor hypofunction point of view. Front. Mol. Neurosci. 2017, 10, 342. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Sacchi, S.; Bernasconi, M.; Martineau, M.; Mothet, J.P.; Ruzzene, M.; Pilone, M.S.; Pollegioni, L.; Molla, G. pLG72 modulates intracellular D-serine levels through its interaction with D-amino acid oxidase: Effect on schizophrenia susceptibility. J. Biol. Chem. 2008, 283, 22244–22256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldinelli, L.; Molla, G.; Bracci, L.; Lelli, B.; Pileri, S.; Cappelletti, P.; Sacchi, S.; Pollegioni, L. Effect of ligand binding on human D-amino acid oxidase: Implications for the development of new drugs for schizophrenia treatment. Protein Sci. 2010, 19, 1500–1512. [Google Scholar] [CrossRef] [Green Version]
- Pollegioni, L.; Piubelli, L.; Molla, G.; Rosini, E. D-Amino acid oxidase-pLG72 interaction and D-serine modulation. Front. Mol. Biosci. 2018, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Kvajo, M.; Dhilla, A.; Swor, D.E.; Karayiorgou, M.; Gogos, J.A. Evidence implicating the candidate schizophrenia/bipolar disorder susceptibility gene G72 in mitochondrial function. Mol. Psychiatry 2008, 13, 685–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollegioni, L.; Molla, G.; Sacchi, S.; Murtas, G. Human D-aspartate oxidase: A key player in D-aspartate metabolism. Front Mol Biosci. 2021, 8, 689719. [Google Scholar] [CrossRef]
- Sacchi, S.; Cappelletti, P.; Giovannardi, S.; Pollegioni, L. Evidence for the interaction of D-amino acid oxidase with pLG72 in a glial cell line. Mol. Cell Neurosci. 2011, 48, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Cappelletti, P.; Campomenosi, P.; Pollegioni, L.; Sacchi, S. The degradation (by distinct pathways) of human D-amino acid oxidase and its interacting partner pLG72-two key proteins in D-serine catabolism in the brain. FEBS J. 2014, 281, 708–723. [Google Scholar] [CrossRef] [Green Version]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.L.; Hsieh, C.H.; Chen, Y.J.; Wang, C.M.; Shih, C.S.; Huang, P.W.; Mir, A.; Lane, H.Y.; Tsai, G.E.; Chang, H.T. The C-terminal region of G72 increases D-amino acid oxidase activity. Int. J. Mol. Sci. 2013, 15, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otte, D.M.; Raskó, T.; Wang, M.; Dreiseidler, M.; Drews, E.; Schrage, H.; Wojtalla, A.; Höhfeld, J.; Wanker, E.; Zimmer, A. Identification of the mitochondrial MSRB2 as a binding partner of LG72. Cell Mol. Neurobiol. 2014, 34, 1123–1130. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Park, S.K. Molecular links between mitochondrial dysfunctions and schizophrenia. Mol. Cells 2012, 33, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Molla, G.; Chaves-Sanjuan, A.; Savinelli, A.; Nardini, M.; Pollegioni, L. Structure and kinetic properties of human D-aspartate oxidase, the enzyme-controlling d-aspartate levels in brain. FASEB J. 2020, 34, 1182–1197. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Saw, H.P.; Cui, F.F.; Lin, S.Y.; Chang, H.T.; Chiu, C.D. pLG72 induces superoxide radicals via interaction and aggregation with SOD1. Free Radic. Res. 2018, 52, 970–976. [Google Scholar] [CrossRef]
- Murtas, G.; Sacchi, S.; Kumar, M.S.; Bosco, D.A.; Pollegioni, L. Is the primate-specific protein pLG72 affecting SOD1 functionality and superoxide formation? Free Radic. Res. 2020, 54, 419–430. [Google Scholar] [CrossRef]
- Korostishevsky, M.; Kaganovich, M.; Cholostoy, A.; Ashkenazi, M.; Ratner, Y.; Dahary, D.; Bernstein, J.; Bening-Abu-Shach, U.; Ben-Asher, E.; Lancet, D.; et al. Is the G72/G30 locus associated with schizophrenia? single nucleotide polymorphisms, haplotypes, and gene expression analysis. Biol. Psychiatry 2004, 56, 169–176. [Google Scholar] [CrossRef]
- Otte, D.M.; Sommersberg, B.; Kudin, A.; Guerrero, C.; Albayram, O.; Filiou, M.D.; Frisch, P.; Yilmaz, O.; Drews, E.; Turck, C.W.; et al. N-acetyl cysteine treatment rescues cognitive deficits induced by mitochondrial dysfunction in G72/G30 transgenic mice. Neuropsychopharmacology 2011, 36, 2233–2243. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chen, H.J.; Zhang, J.; Li, W.; Xie, X.; Chang, H.T. Identification of pLG72-induced oxidative stress using systemic approaches. Biomed Res. Int. 2015, 2015, 429253. [Google Scholar] [CrossRef] [Green Version]
- Korde, A.S.; Maragos, W.F. Identification of an N-methyl-D-aspartate receptor in isolated nervous system mitochondria. J. Biol. Chem. 2012, 287, 35192–35200. [Google Scholar] [CrossRef] [Green Version]
- Filiou, M.D.; Teplytska, L.; Nussbaumer, M.; Otte, D.M.; Zimmer, A.; Turck, C.W. Multi-omics analysis reveals myelin, presynaptic and nicotinate alterations in the hippocampus of G72/G30 transgenic mice. J. Pers. Med. 2022, 12, 244. [Google Scholar] [CrossRef]
- Popiolek, M.; Ross, J.F.; Charych, E.; Chanda, P.; Gundelfinger, E.D.; Moss, S.J.; Brandon, N.J.; Pausch, M.H. D-amino acid oxidase activity is inhibited by an interaction with bassoon protein at the presynaptic active zone. J. Biol. Chem. 2011, 286, 28867–28875. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Jagannath, V.; Marinova, Z.; Monoranu, C.M.; Walitza, S.; Grünblatt, E. Expression of D-amino acid oxidase (DAO/DAAO) and D-amino acid oxidase activator (DAOA/G72) during development and aging in the human post-mortem brain. Front. Neuroanat. 2017, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.; Punzo, D.; Cuomo, M.; Affinito, O.; Coretti, L.; Sacchi, S.; Florio, E.; Lembo, F.; Carella, M.; Copetti, M.; et al. DNA methylation landscape of the genes regulating D-serine and D-aspartate metabolism in post-mortem brain from controls and subjects with schizophrenia. Sci. Rep. 2018, 8, 10163. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.S.; Shin, W.J.; Lee, J.E.; Do, J.T. CpG and Non-CpG methylation in epigenetic gene regulation and brain gunction. Genes 2017, 8, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.H.; Chang, H.T.; Chen, Y.J.; Lin, C.H.; Huang, C.H.; Tun, R.; Tsai, G.E.; Lane, H.Y. Distinctively higher plasma G72 protein levels in patients with schizophrenia than in healthy individuals. Mol. Psychiatry 2014, 19, 636–637. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Chiu, C.C.; Huang, C.H.; Yang, H.T.; Lane, H.Y. pLG72 levels increase in early phase of Alzheimer’s disease but decrease in late phase. Sci. Rep. 2019, 9, 13221. [Google Scholar] [CrossRef] [PubMed]
- Akyol, E.S.; Albayrak, Y.; Aksoy, N.; Şahin, B.; Beyazyüz, M.; Kuloğlu, M.; Hashimoto, K. Increased serum G72 protein levels in patients with schizophrenia: A potential candidate biomarker. Acta Neuropsychiatr. 2017, 29, 80–86. [Google Scholar] [CrossRef]
- Lin, E.; Lin, C.H.; Lai, Y.L.; Huang, C.H.; Huang, Y.J.; Lane, H.Y. Combination of G72 genetic variation and G72 protein level to detect schizophrenia: Machine learning approaches. Front. Psychiatry 2018, 9, 566. [Google Scholar] [CrossRef]
- Lin, E.; Lin, C.H.; Hung, C.C.; Lane, H.Y. An ensemble approach to predict schizophrenia using protein data in the N-methyl-D-aspartate receptor (NMDAR) and tryptophan catabolic pathways. Front. Bioeng. Biotechnol. 2020, 8, 569. [Google Scholar] [CrossRef]
- Piubelli, L.; Pollegioni, L.; Rabattoni, V.; Mauri, M.; Princiotta Cariddi, L.; Versino, M.; Sacchi, S. Serum D-serine levels are altered in early phases of Alzheimer’s disease: Towards a precocious biomarker. Transl. Psychiatry 2021, 11, 77. [Google Scholar] [CrossRef]
- Piubelli, L.; Murtas, G.; Rabattoni, V.; Pollegioni, L. The role of D-amino acids in Alzheimer’s Disease. J. Alzheimers Dis. 2021, 80, 475–492. [Google Scholar] [CrossRef]
- Coyle, J.T. D-serine and the pathophysiology of schizophrenia. In D-Amino Acids; Springer: Tokyo, Japan, 2016; pp. 101–118. [Google Scholar] [CrossRef]
- Terry-Lorenzo, R.T.; Masuda, K.; Sugao, K.; Fang, Q.K.; Orsini, M.A.; Sacchi, S.; Pollegioni, L. High-throughput screening strategy identifies allosteric, covalent human D-amino acid oxidase inhibitor. J. Biomol. Screen. 2015, 20, 1218–1231. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Sample | Healthy Controls | Pathological State | |||
|---|---|---|---|---|---|
| MCI (CDR = 0.5) | Mild AD (CDR = 1) | Moderate AD (CDR = 2) | AD (CDR = 3) | ||
| Plasma a ng/mL | 1.4 ± 0.7 (108) | 1.4 ± 0.7 (81) | 2.3 ± 1.1 (124) | 2.9 ± 1.6 (35) | 1.4 ± 0.7 (28) |
| MDD | SZ | ||||
| Plasma b pg/mL | ~300 (27) ~415 (30) | ~320 (26) | ~280 (27) ~390 (40) | ||
| CSF b pg/mL | ~ 20.5 | ~20 | ~18 | ||
| Plasma c µg/mL | ~1.2 (30) | ~5 (30) medicated ~4 (27) drug free | |||
| Plasma d µg/mL | 1.147 ± 0.574 (60) | 4.057 ± 2.594 (89) unmatched 4.188 ± 2.772 (66) matched | |||
| Serum d ng/mL | 0.346 ± 0.102 (60) | 0.496 ± 0.152 (107) | |||
| Plasma e µg/mL | 1.68 ± 0.81 (86) | 3.24 ± 1.80 (355) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murtas, G.; Pollegioni, L.; Molla, G.; Sacchi, S. Biochemical Properties and Physiological Functions of pLG72: Twenty Years of Investigations. Biomolecules 2022, 12, 858. https://doi.org/10.3390/biom12060858
Murtas G, Pollegioni L, Molla G, Sacchi S. Biochemical Properties and Physiological Functions of pLG72: Twenty Years of Investigations. Biomolecules. 2022; 12(6):858. https://doi.org/10.3390/biom12060858
Chicago/Turabian StyleMurtas, Giulia, Loredano Pollegioni, Gianluca Molla, and Silvia Sacchi. 2022. "Biochemical Properties and Physiological Functions of pLG72: Twenty Years of Investigations" Biomolecules 12, no. 6: 858. https://doi.org/10.3390/biom12060858
APA StyleMurtas, G., Pollegioni, L., Molla, G., & Sacchi, S. (2022). Biochemical Properties and Physiological Functions of pLG72: Twenty Years of Investigations. Biomolecules, 12(6), 858. https://doi.org/10.3390/biom12060858