
Effect of Gum Acacia on the Intestinal Bioavailability of n-3 Polyunsaturated Fatty Acids in Rats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Lipid Formulations
2.3. Experimental Design: Animal and Surgical Procedures
2.4. Fatty Acid Profile and N-3 LC PUFA Composition in Lymph
2.5. Statistical Analysis
3. Results
3.1. Influence of Using Gum Acacia as Emulsifier on the Intestinal Absorption of N-3 LC-PUFA
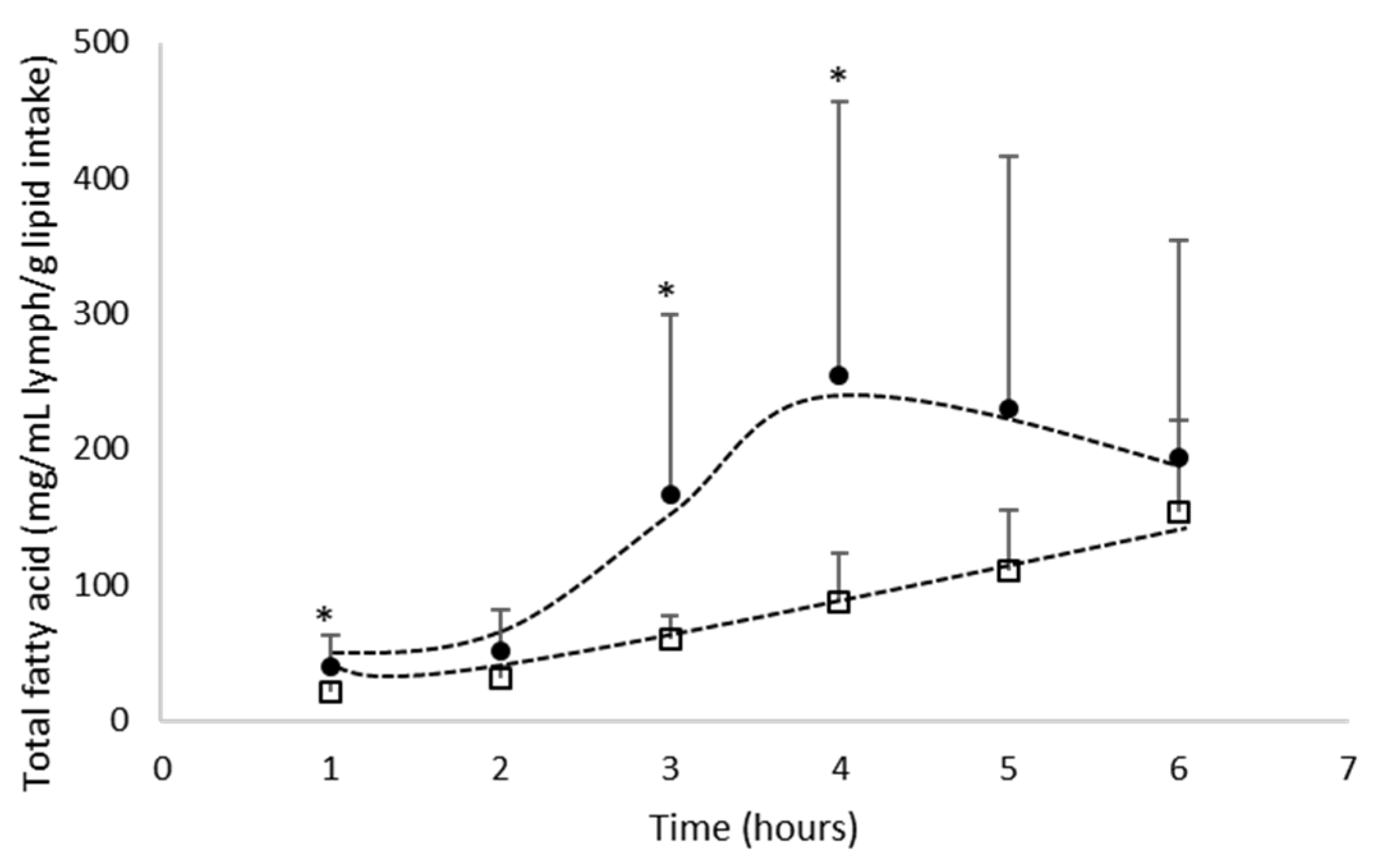
3.1.1. Total Fatty Acid Absorption
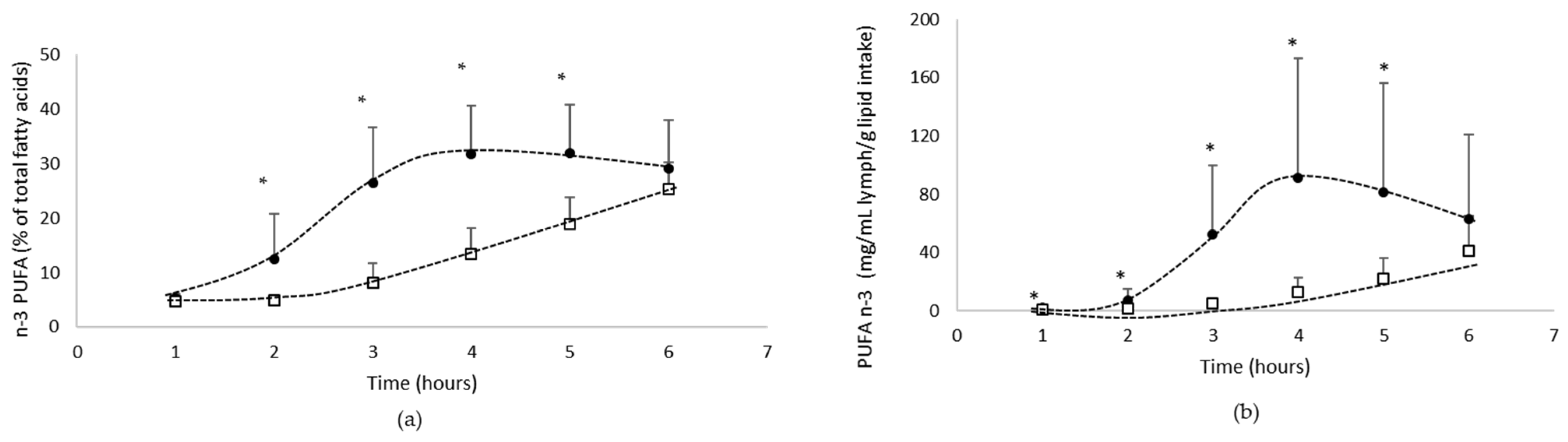
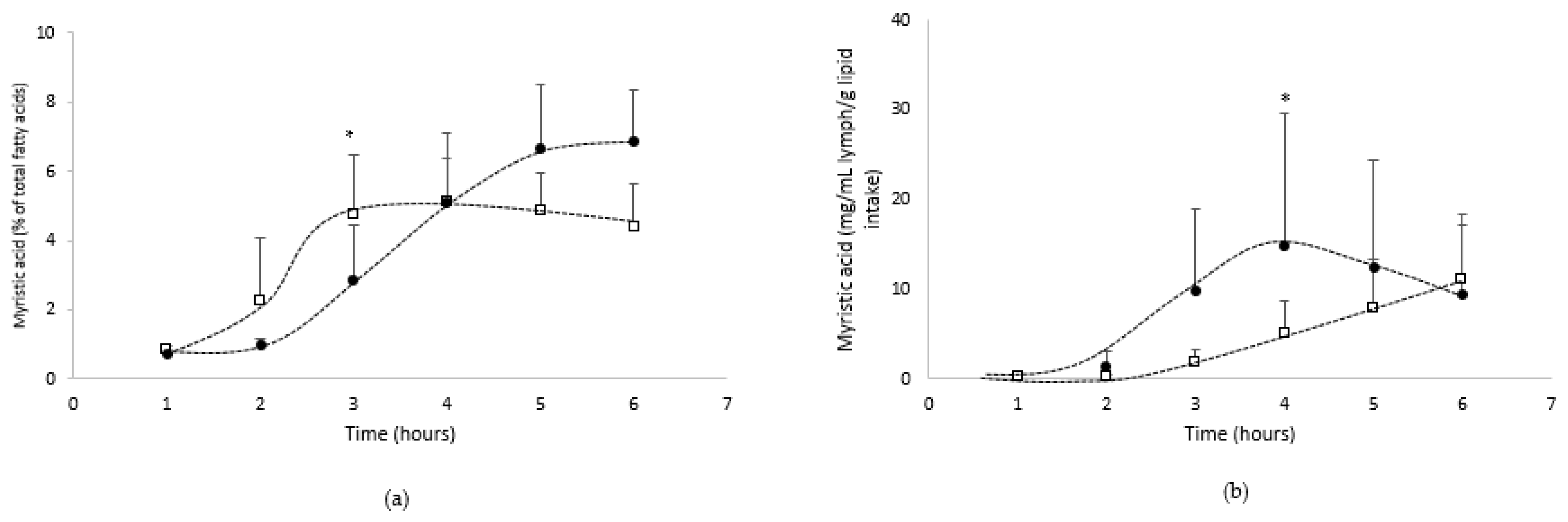
3.1.2. Lymphatic Recovery of N-3 PUFA
- Total n-3 PUFA absorption
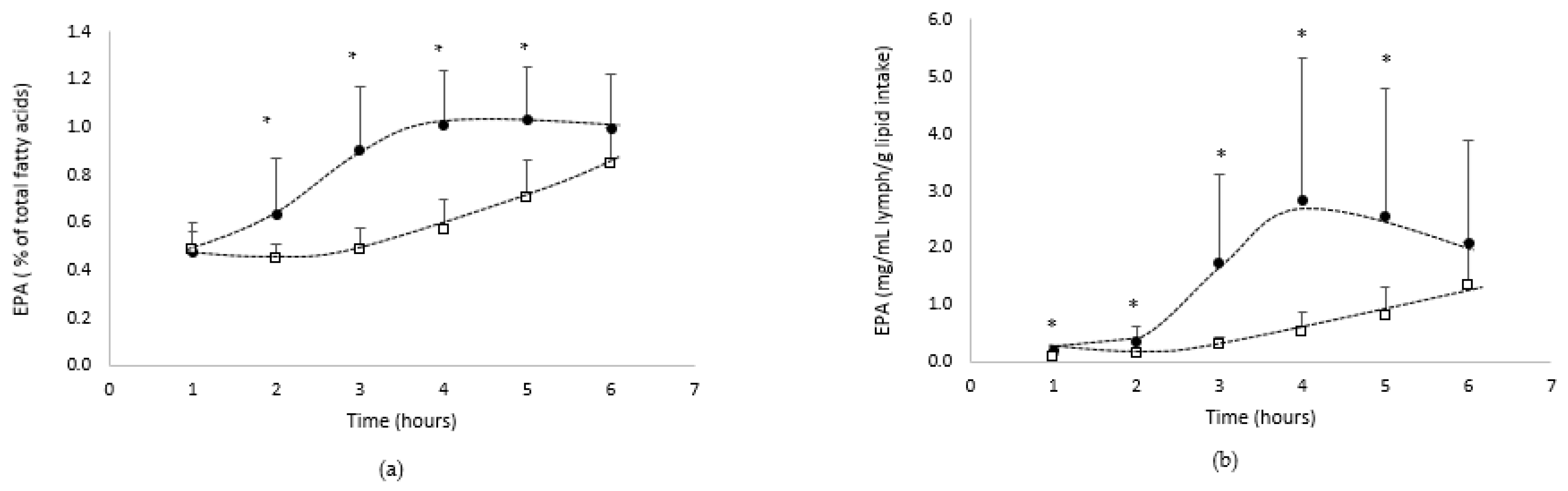
- EPA absorption
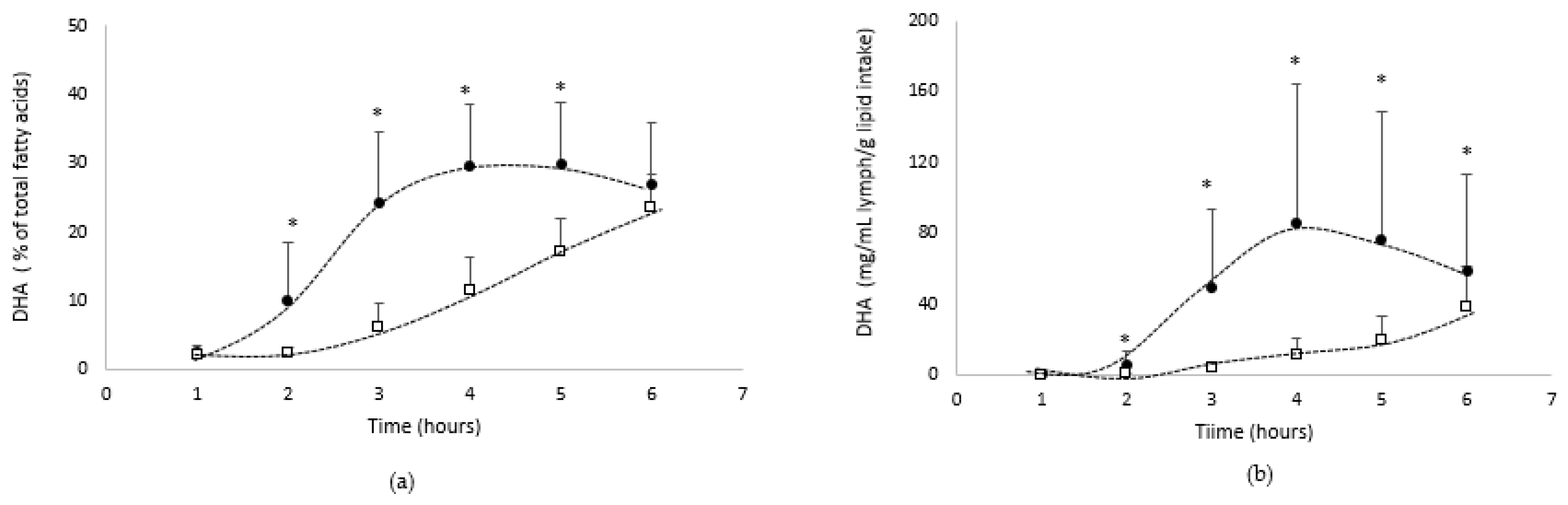
- DHA absorption
- Absorption of the other main fatty acids from the oil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubuisson, C.; Carrillo, S.; Dufour, A.; Havard, S.; Pinard, P.; Volatier, J.-L. The French Dietary Survey on the General Population (INCA3). EFSA Supporting Publ. 2017, 14, 1351E. [Google Scholar] [CrossRef] [Green Version]
- ANSES. Actualisation des Apports Nutritionnels Conseillés Pour les Acides Gras. 2011. Available online: NUT2006sa0359Ra.pdf (accessed on 12 December 2021).
- Avis de L’Anses Rapport D’expertise Collective Étude Individuelle Nationale des Consommations Alimentaires 3 (INCA 3). 2017. INCA 3: Evolution des Habitudes et Modes de Consommation, de Nouveaux Enjeux en Matière de Sécurité Sanitaire et de Nutrition|Anses—Agence Nationale de Sécurité Sanitaire de L’alimentation, de L’environnement et du Travail. Available online: https://www.vie-publique.fr/rapport/36772-etude-individuelle-nationale-des-consommations-alimentaires-3-inca-3 (accessed on 12 December 2021).
- Saini, R.K.; Keum, Y.-S. Omega-3 and Omega-6 Polyunsaturated Fatty Acids: Dietary Sources, Metabolism, and Significance—A Review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Moghadasian, M.H. A Systemic Review of the Roles of n-3 Fatty Acids in Health and Disease. J. Am. Diet. Assoc. 2009, 109, 668–679. [Google Scholar]
- Dyall, S.C.; Michael-Titus, A.T. Neurological Benefits of Omega-3 Fatty Acids. Neuromolecular Med. 2008, 10, 219–235. [Google Scholar] [CrossRef]
- Calder, P.C. Very Long-Chain n-3 Fatty Acids and Human Health: Fact, Fiction and the Future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Marine Omega-3 Fatty Acids and Inflammatory Processes: Effects, Mechanisms and Clinical Relevance. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Zárate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of Long Chain Polyunsaturated Fatty Acids in Human Health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Cansell, M. Marine Phospholipids as Dietary Carriers of Long-Chain Polyunsaturated Fatty Acids. Lipid Technol. 2010, 22, 223–226. [Google Scholar] [CrossRef]
- Couëdelo, L.; Amara, S.; Lecomte, M.; Meugnier, E.; Monteil, J.; Fonseca, L.; Pineau, G.; Cansell, M.; Carrière, F.; Michalski, M.C.; et al. Impact of Various Emulsifiers on ALA Bioavailability and Chylomicron Synthesis through Changes in Gastrointestinal Lipolysis. Food Funct. 2015, 6, 1726–1735. [Google Scholar] [CrossRef]
- Couëdelo, L.; Boué-Vaysse, C.; Fonseca, L.; Montesinos, E.; Djoukitch, S.; Combe, N.; Cansell, M. Lymphatic Absorption of α-Linolenic Acid in Rats Fed Flaxseed Oil-Based Emulsion. Br. J. Nutr. 2011, 105, 1026–1035. [Google Scholar] [CrossRef] [Green Version]
- Garaiova, I.; Guschina, I.A.; Plummer, S.F.; Tang, J.; Wang, D.; Plummer, N.T. A Randomised Cross-over Trial in Healthy Adults Indicating Improved Absorption of Omega-3 Fatty Acids by Pre-Emulsification. Nutr. J. 2007, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalski, M.C.; Genot, C.; Gayet, C.; Lopez, C.; Fine, F.; Joffre, F.; Vendeuvre, J.L.; Bouvier, J.; Chardigny, J.M.; Raynal-Ljutovac, K. Multiscale Structures of Lipids in Foods as Parameters Affecting Fatty Acid Bioavailability and Lipid Metabolism. Prog. Lipid Res. 2013, 52, 354–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, O.; Meraihi, Z.; Mura, J.L.; Frey, A.; Riess, G.H.; Bach, A.C. Fat Emulsion Particle Size: Influence on the Clearance Rate and the Tissue Lipolytic Activity. Am. J. Clin. Nutr. 1989, 50, 1370–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugasini, D.; Devaraj, V.C.; Ramesh, M.; Lokesh, B.R. Lymphatic Transport of α-Linolenic Acid and Its Conversion to Long Chain n-3 Fatty Acids in Rats Fed Microemulsions of Linseed Oil. Lipids 2014, 49, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Sehl, A.; Couëdelo, L.; Vaysse, C.; Cansell, M. Intestinal Bioavailability of N-3 Long-Chain Polyunsaturated Fatty Acids Influenced by the Supramolecular Form of Phospholipids. Food Funct. 2020, 11, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Lane, K.E.; Li, W.; Smith, C.; Derbyshire, E. The Bioavailability of an Omega-3-Rich Algal Oil Is Improved by Nanoemulsion Technology Using Yogurt as a Food Vehicle. Int. J. Food Sci. Technol. 2014, 49, 1264–1271. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Wang, Q.; Li, W.; Wright, A.J. Emulsification of Algal Oil with Soy Lecithin Improved DHA Bioaccessibility but Did Not Change Overall in Vitro Digestibility. Food Funct. 2014, 5, 2913–2921. [Google Scholar] [CrossRef]
- Austria, J.A.; Richard, M.N.; Chahine, M.N.; Edel, A.L.; Malcolmson, L.J.; Dupasquier, C.M.C.; Pierce, G.N. Bioavailability of Alpha-Linolenic Acid in Subjects after Ingestion of Three Different Forms of Flaxseed. J. Am. Coll. Nutr. 2008, 27, 214–221. [Google Scholar] [CrossRef]
- Michalski, M.-C.; Couëdelo, L.; Penhoat, A.; Vaysse, C.; Vors, C. Chapter 2—Bioavailability and Metabolism of Dietary Lipids. In Lipids and Edible Oils; Galanakis, C.M., Ed.; Academic Press: Amsterdam, The Netherlands, 2020; pp. 45–92. ISBN 978-0-12-817105-9. [Google Scholar]
- Borgström, B. On the Interactions between Pancreatic Lipase and Colipase and the Substrate, and the Importance of Bile Salts. J. Lipid Res. 1975, 16, 411–417. [Google Scholar] [CrossRef]
- Armand, M. Lipases and Lipolysis in the Human Digestive Tract: Where Do We Stand? Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 156–164. [Google Scholar] [CrossRef]
- Golding, M.; Wooster, T.J. The Influence of Emulsion Structure and Stability on Lipid Digestion. Curr. Opin. Colloid Interface Sci. 2010, 15, 90–101. [Google Scholar] [CrossRef]
- Porsgaard, T.; Xu, X.; Göttsche, J.; Mu, H. Differences in the Intramolecular Structure of Structured Oils Do Not Affect Pancreatic Lipase Activity in Vitro or the Absorption by Rats of (n-3) Fatty Acids. J. Nutr. 2005, 135, 1705–1711. [Google Scholar] [CrossRef]
- Couëdelo, L.; Termon, A.; Vaysse, C. Matrice lipidique et biodisponibilité de l’acide alpha-linolénique. OCL 2017, 24, D204. [Google Scholar] [CrossRef] [Green Version]
- Bottino, N.R.; Vandenburg, G.A.; Reiser, R. Resistance of Certain Long-Chain Polyunsaturated Fatty Acids of Marine Oils to Pancreatic Lipase Hydrolysis. Lipids 1967, 2, 489–493. [Google Scholar] [CrossRef]
- Lamothe, S.; Jolibois, É.; Britten, M. Effect of Emulsifiers on Linseed Oil Emulsion Structure, Lipolysis and Oxidation during in Vitro Digestion. Food Funct. 2020, 11, 10126–10136. [Google Scholar] [CrossRef]
- Singh, H.; Ye, A.; Horne, D. Structuring Food Emulsions in the Gastrointestinal Tract to Modify Lipid Digestion. Prog. Lipid Res. 2009, 48, 92–100. [Google Scholar] [CrossRef]
- Armand, M.; Pasquier, B.; André, M.; Borel, P.; Senft, M.; Peyrot, J.; Salducci, J.; Portugal, H.; Jaussan, V.; Lairon, D. Digestion and Absorption of 2 Fat Emulsions with Different Droplet Sizes in the Human Digestive Tract. Am. J. Clin. Nutr. 1999, 70, 1096–1106. [Google Scholar] [CrossRef]
- McClements, D.J.; Saliva-Trujillo, L.; Zhang, R.; Zhang, Z.; Zou, L.; Yao, M.; Xiao, H. Boosting the Bioavailability of Hydrophobic Nutrients, Vitamins, and Nutraceuticals in Natural Products Using Excipient Emulsions. Food Res. Int. 2016, 88, 140–152. [Google Scholar] [CrossRef] [Green Version]
- Vors, C.; Lecomte, M.; Michalski, M.-C. Impact de la structure émulsionnée des lipides sur le devenir métabolique des acides gras alimentaires. Cah. Nutr. Diététique 2016, 51, 238–247. [Google Scholar] [CrossRef]
- Dapčević Hadnađev, T.; Dokić, P.; Krstonošić, V.; Hadnađev, M. Influence of Oil Phase Concentration on Droplet Size Distribution and Stability of Oil-in-Water Emulsions. Eur. J. Lipid Sci. Technol. 2013, 115, 313–321. [Google Scholar] [CrossRef]
- O’Driscoll, C.M. Lipid-Based Formulations for Intestinal Lymphatic Delivery. Eur. J. Pharm. Sci. 2002, 15, 405–415. [Google Scholar] [CrossRef]
- Parthasarathi, S.; Muthukumar, S.P.; Anandharamakrishnan, C. The Influence of Droplet Size on the Stability, in Vivo Digestion, and Oral Bioavailability of Vitamin E Emulsions. Food Funct. 2016, 7, 2294–2302. [Google Scholar] [CrossRef]
- Michalski, M.-C.; Soares, A.F.; Lopez, C.; Leconte, N.; Briard, V.; Geloen, A. The Supramolecular Structure of Milk Fat Influences Plasma Triacylglycerols and Fatty Acid Profile in the Rat. Eur. J. Nutr. 2006, 45, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Rhazi, L.; Lakahal, L.; Andrieux, O.; Niamba, N.; Depeint, F.; Guillemet, D. Relationship between the Molecular Characteristics of Acacia Gum and Its Functional Properties. Food Chem. 2020, 328, 126860. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Katayama, T.; Ogasawara, T.; Sasaki, Y.; Al-Assaf, S.; Phillips, G.O. Characterization and Properties of Acacia Senegal (L.) Willd. Var. Senegal with Enhanced Properties (Acacia (Sen) SUPER GUM TM ): Part 5. Factors Affecting the Emulsification of Acacia Senegal and Acacia (Sen) SUPER GUMTM. Food Hydrocoll. 2007, 21, 353–358. [Google Scholar] [CrossRef]
- Buffo, R.; Reineccius, G. Beverage Emulsions and the Utilization of Gum Acacia as Emulsifier/Stabilizer. Perfum. Flavorist 2000, 25, 24–44. [Google Scholar]
- Atgié, M.; Masbernat, O.; Roger, K. Emulsions Stabilized by Gum Arabic: Composition and Packing within Interfacial Films. Langmuir 2019, 35, 962–972. [Google Scholar] [CrossRef]
- Dag, D.; Guner, S.; Oztop, M.H. Physicochemical Mechanisms of Different Biopolymers’ (Lysozyme, Gum Arabic, Whey Protein, Chitosan) Adsorption on Green Tea Extract Loaded Liposomes. Int. J. Biol. Macromol. 2019, 138, 473–482. [Google Scholar] [CrossRef]
- Fang, X.; Shima, M.; Adachi, S. Effects of Drying Conditions on the Oxidation of Linoleic Acid Encapsulated with Gum Arabic by Spray-Drying. Food Sci. Technol. Res. 2005, 11, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Omar, K.A.; Shan, L.; Zou, X.; Song, Z.; Wang, X. Effects of Two Emulsifiers on Yield and Storage of Flaxseed Oil Powder by Response Surface Methodology. Pak. J. Nutr. 2009, 8, 1316–1324. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, L.; Li, D.; Adhikari, B.; Shi, J. Effect of Gum Arabic on Stability of Oil-in-Water Emulsion Stabilized by Flaxseed and Soybean Protein. Carbohydr. Polym. 2011, 86, 343–351. [Google Scholar] [CrossRef]
- Akram, S.; Bao, Y.; Butt, M.S.; Shukat, R.; Afzal, A.; Huang, J.-Y. Fabrication and Characterization of Gum Arabic- and Maltodextrin-Based Microcapsules Containing Polyunsaturated Oils. J. Sci. Food Agric. 2021, 101, 6384–6394. [Google Scholar] [CrossRef]
- Berton-Carabin, C.; Schröder, A.; Schroën, K.; Laguerre, M. Chapter 14—Lipid Oxidation in Pickering Emulsions. In Omega-3 Delivery Systems; García-Moreno, P.J., Jacobsen, C., Moltke Sørensen, A.-D., Yesiltas, B., Eds.; Academic Press: Amsterdam, The Netherlands, 2021; pp. 275–293. ISBN 978-0-12-821391-9. [Google Scholar]
- Wyatt, G.M.; Bayliss, C.E.; Holcroft, J.D. A Change in Human Faecal Flora in Response to Inclusion of Gum Arabic in the Diet. Br. J. Nutr. 1986, 55, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Michel, C.; Kravtchenko, T.P.; David, A.; Gueneau, S.; Kozlowski, F.; Cherbut, C. In Vitro Prebiotic Effects of Acacia Gums onto the Human Intestinal Microbiota Depends on Both Botanical Origin and Environmental PH. Anaerobe 1998, 4, 257–266. [Google Scholar] [CrossRef]
- Cherbut, C.; Michel, C.; Raison, V.; Kravtchenko, T.; Severine, M. Acacia Gum Is a Bifidogenic Dietary Fibre with High Digestive Tolerance in Healthy Humans. Microb. Ecol. Health Dis. 2003, 15, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Terpend, K.; Possemiers, S.; Daguet, D.; Marzorati, M. Arabinogalactan and Fructo-Oligosaccharides Have a Different Fermentation Profile in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). Environ. Microbiol. Rep. 2013, 5, 595–603. [Google Scholar] [CrossRef]
- Tan, Y.; Zhang, Z.; Muriel Mundo, J.; McClements, D.J. Factors Impacting Lipid Digestion and Nutraceutical Bioaccessibility Assessed by Standardized Gastrointestinal Model (INFOGEST): Emulsifier Type. Food Res. Int. 2020, 137, 109739. [Google Scholar] [CrossRef]
- Tamargo, A.; Martin, D.; Navarro del Hierro, J.; Moreno-Arribas, M.V.; Muñoz, L.A. Intake of Soluble Fibre from Chia Seed Reduces Bioaccessibility of Lipids, Cholesterol and Glucose in the Dynamic Gastrointestinal Model Simgi®. Food Res. Int. 2020, 137, 109364. [Google Scholar] [CrossRef]
- Uebelhack, R.; Bongartz, U.; Seibt, S.; Bothe, G.; Chong, P.W.; De Costa, P.; Wszelaki, N. Double-Blind, Randomized, Three-Armed, Placebo-Controlled, Clinical Investigation to Evaluate the Benefit and Tolerability of Two Dosages of IQP-AE-103 in Reducing Body Weight in Overweight and Moderately Obese Subjects. J. Obes. 2019, 2019, 3412952. [Google Scholar] [CrossRef]
- Bollman, J.L.; Cain, J.C.; Grindlay, J.H. Techniques for the Collection of Lymph from the Liver, Small Intestine, or Thoracic Duct of the Rat. J. Lab. Clin. Med. 1948, 33, 1349–1352. [Google Scholar]
- Lepage, G.; Roy, C.C. Specific Methylation of Plasma Nonesterified Fatty Acids in a One-Step Reaction. J. Lipid Res. 1988, 29, 227–235. [Google Scholar] [CrossRef]
- Ikeda, I.; Sasaki, E.; Yasunami, H.; Nomiyama, S.; Nakayama, M.; Sugano, M.; Imaizumi, K.; Yazawa, K. Digestion and Lymphatic Transport of Eicosapentaenoic and Docosahexaenoic Acids given in the Form of Triacylglycerol, Free Acid and Ethyl Ester in Rats. Biochim. Biophys. Acta 1995, 1259, 297–304. [Google Scholar] [CrossRef]
- Ikeda, I.; Yoshida, H.; Imaizumi, K. Effects of Triolein or Oleic Acid on Lymphatic Recovery of Docosahexaenoic Acid given as Ethyl Ester and Their Intramolecular Distribution in Lymph Triglyceride of Rats. Lipids 1997, 32, 949–952. [Google Scholar] [CrossRef]
- Lambert, M.S.; Botham, K.M.; Mayes, P.A. Modification of the Fatty Acid Composition of Dietary Oils and Fats on Incorporation into Chylomicrons and Chylomicron Remnants. Br. J. Nutr. 1996, 76, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Carrière, F.; Verger, R.; Lookene, A.; Olivecrona, G. Lipase Structures at the Interface between Chemistry and Biochemistry. EXS 1995, 73, 3–26. [Google Scholar] [PubMed]
- Bonnet, M.; Cansell, M.; Placin, F.; Monteil, J.; Anton, M.; Leal-Calderon, F. Influence of the Oil Globule Fraction on the Release Rate Profiles from Multiple W/O/W Emulsions. Colloids Surf. B Biointerfaces 2010, 78, 44–52. [Google Scholar] [CrossRef]
- Nishimukai, M.; Hara, H.; Aoyama, Y. The Addition of Soybean Phosphatidylcholine to Triglyceride Increases Suppressive Effects on Food Intake and Gastric Emptying in Rats. J. Nutr. 2003, 133, 1255–1258. [Google Scholar] [CrossRef]
- Nishimukai, M.; Hara, H. Soybean Phosphatidylcholine-Induced Enhancement of Lymphatic Absorption of Triglyceride Depends on Chylomicron Formation in Rats. Biosci. Biotechnol. Biochem. 2007, 71, 1192–1197. [Google Scholar] [CrossRef]
- Raatz, S.K.; Redmon, J.B.; Wimmergren, N.; Donadio, J.V.; Bibus, D.M. Enhanced Absorption of N-3 Fatty Acids from Emulsified Compared with Encapsulated Fish Oil. J. Am. Diet. Assoc. 2009, 109, 1076–1081. [Google Scholar] [CrossRef] [Green Version]
- Christensen, M.S.; Müllertz, A.; Høy, C.E. Absorption of Triglycerides with Defined or Random Structure by Rats with Biliary and Pancreatic Diversion. Lipids 1995, 30, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Laguerre, M.; Tenon, M.; Schroën, K.; Berton-Carabin, C.C. Natural Particles Can Armor Emulsions against Lipid Oxidation and Coalescence. Food Chem. 2021, 347, 129003. [Google Scholar] [CrossRef]
- Adams, S.; Sello, C.T.; Qin, G.-X.; Che, D.; Han, R. Does Dietary Fiber Affect the Levels of Nutritional Components after Feed Formulation? Fibers 2018, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, I.A. The Physiological Roles of Dietary Fibre. Food Hydrocoll. 2011, 25, 238–250. [Google Scholar] [CrossRef]
- Desnuelle, P. Pancreatic Lipase. Adv. Enzym. Relat. Subj. Biochem. 1961, 23, 129–161. [Google Scholar]
- Akanbi, T.O.; Sinclair, A.J.; Barrow, C.J. Pancreatic Lipase Selectively Hydrolyses DPA over EPA and DHA Due to Location of Double Bonds in the Fatty Acid Rather than Regioselectivity. Food Chem. 2014, 160, 61–66. [Google Scholar] [CrossRef]
- Carrière, F.; Renou, C.; Lopez, V.; De Caro, J.; Ferrato, F.; Lengsfeld, H.; De Caro, A.; Laugier, R.; Verger, R. The Specific Activities of Human Digestive Lipases Measured from the in Vivo and in Vitro Lipolysis of Test Meals. Gastroenterology 2000, 119, 949–960. [Google Scholar] [CrossRef]
- Bonnaire, L.; Sandra, S.; Helgason, T.; Decker, E.A.; Weiss, J.; McClements, D.J. Influence of Lipid Physical State on the in Vitro Digestibility of Emulsified Lipids. J. Agric. Food Chem. 2008, 56, 3791–3797. [Google Scholar] [CrossRef]
- Dille, M.J.; Baydin, T.; Kristiansen, K.A.; Draget, K.I. The Impact of Emulsion Droplet Size on in Vitro Lipolysis Rate and in Vivo Plasma Uptake Kinetics of Triglycerides and Vitamin D3 in Rats. Food Funct. 2021, 12, 3219–3232. [Google Scholar] [CrossRef]
- McClements, D.J.; Li, Y. Structured Emulsion-Based Delivery Systems: Controlling the Digestion and Release of Lipophilic Food Components. Adv. Colloid Interface Sci. 2010, 159, 213–228. [Google Scholar] [CrossRef]
- Ramírez, M.; Amate, L.; Gil, A. Absorption and Distribution of Dietary Fatty Acids from Different Sources. Early Hum. Dev. 2001, 65, S95–S101. [Google Scholar] [CrossRef]
- Embleton, J.K.; Pouton, C.W. Structure and Function of Gastro-Intestinal Lipases. Adv. Drug Deliv. Rev. 1997, 25, 15–32. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oil | Emulsion | |
|---|---|---|
| Total fatty acid (%) | ||
| SFA * | 30.9 | 33.4 |
| 12:0 | 0.7 | 0.8 |
| 14:0 | 8.6 | 9.1 |
| 16:0 | 20.2 | 21.4 |
| 18:0 | 0.7 | 0.7 |
| MUFA * | 9.8 | 10.5 |
| 14:1 | 0.1 | 0.1 |
| 16:1 | 4.0 | 4.3 |
| 18:1 | 5.8 | 6.0 |
| PUFA * | 59.0 | 55.5 |
| PUFA n-6 * | 9.7 | 9.4 |
| 18:2(n-6) | 1.0 | 1.1 |
| 20:4(n-6) | 0.2 | 0.2 |
| 22:5(n-6) | 8.2 | 7.9 |
| PUFA n-3 * | 49.3 | 46.1 |
| 18:4(n-3) | 0.2 | 0.3 |
| 20:5(n-3) | 1.5 | 1.5 |
| 22:5(n-3) | 0.1 | 0.3 |
| 22:6(n-3) | 47.3 | 43.9 |
| Parameters | |
|---|---|
| % of Total Lipids | |
| Polymer | 0.2 |
| Triglyceride | 97.2 |
| Diglyceride | <0.1 |
| Monoglyceride | 0.9 |
| Free fatty acids and others (sterol, FAME *) | 1.6 |
| Parameters | Oil | Emulsion |
|---|---|---|
| Peroxyde value (mEq O2/kg) | 1.5 ± 1.0 | 5.3 ± 2.1 |
| P-anisidine value | 5.1 | 7.1 |
| Oil | GA-Emulsion | |||
|---|---|---|---|---|
| Total FA * | AUC * | 375.1 | 820.9 | $ |
| Cmax * | 153.6 | 254.9 | £ | |
| Tmax * | 6 h | 4 h | ¤ | |
| n-3 PUFA * | AUC | 62.9 | 265.2 | $ |
| Cmax | 41 | 91.1 | £ | |
| Tmax | 6 h | 4 h | ¤ | |
| EPA * | AUC | 2.5 | 8.6 | $ |
| Cmax | 1.3 | 2.8 | £ | |
| Tmax | 6 h | 4 h | ¤ | |
| DHA * | AUC | 55.8 | 248.2 | $ |
| Cmax | 38.4 | 86 | £ | |
| Tmax | 6 h | 4 h | ¤ | |
| Myristic acid | AUC | 20.8 | 43.4 | $ |
| Cmax | 11.2 | 14.9 | £ | |
| Tmax | 6 h | 4 h | ¤ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Couëdelo, L.; Joseph, C.; Abrous, H.; Chamekh-Coelho, I.; Vaysse, C.; Baury, A.; Guillemet, D. Effect of Gum Acacia on the Intestinal Bioavailability of n-3 Polyunsaturated Fatty Acids in Rats. Biomolecules 2022, 12, 975. https://doi.org/10.3390/biom12070975
Couëdelo L, Joseph C, Abrous H, Chamekh-Coelho I, Vaysse C, Baury A, Guillemet D. Effect of Gum Acacia on the Intestinal Bioavailability of n-3 Polyunsaturated Fatty Acids in Rats. Biomolecules. 2022; 12(7):975. https://doi.org/10.3390/biom12070975
Chicago/Turabian StyleCouëdelo, Leslie, Cécile Joseph, Hélène Abrous, Ikram Chamekh-Coelho, Carole Vaysse, Aurore Baury, and Damien Guillemet. 2022. "Effect of Gum Acacia on the Intestinal Bioavailability of n-3 Polyunsaturated Fatty Acids in Rats" Biomolecules 12, no. 7: 975. https://doi.org/10.3390/biom12070975
APA StyleCouëdelo, L., Joseph, C., Abrous, H., Chamekh-Coelho, I., Vaysse, C., Baury, A., & Guillemet, D. (2022). Effect of Gum Acacia on the Intestinal Bioavailability of n-3 Polyunsaturated Fatty Acids in Rats. Biomolecules, 12(7), 975. https://doi.org/10.3390/biom12070975