
Impact of Primary RPE Cells in a Porcine Organotypic Co-Cultivation Model

 ,
,  and
and 
Abstract
:1. Introduction
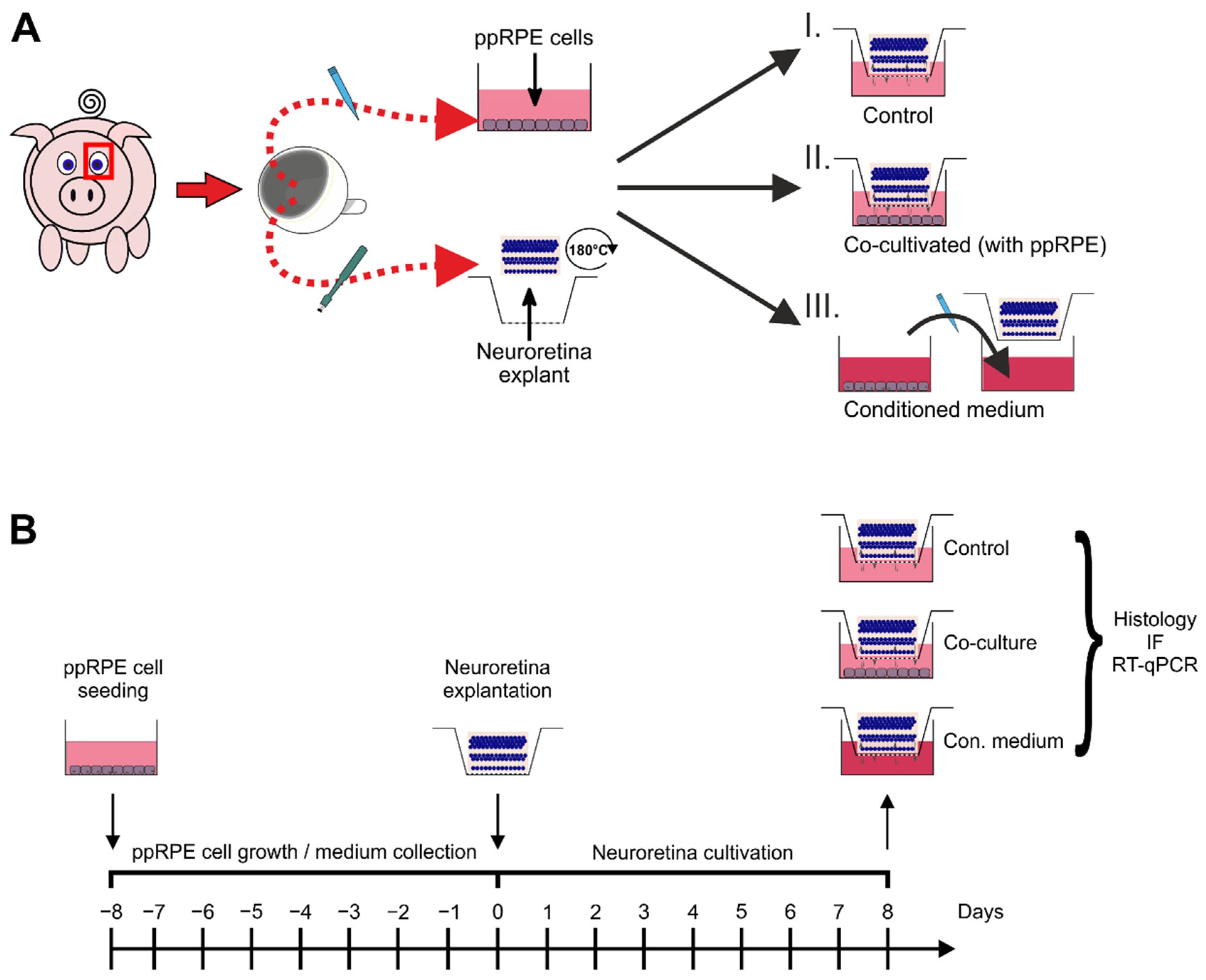
2. Materials and Methods
2.1. Dissection and Isolation of Primary Porcine RPE Cells
2.2. Preparation and Cultivation of Neuroretina Explants
2.3. Co-Cultivation of ppRPE Cells and Neuroretina Explants
2.4. Tissue Processing and Preparation for Histology and RT-qPCR
2.5. (Immuno-) Histology
2.6. Microscopy and Image Assessment
2.7. Quantitative Real-Time PCR (RT-qPCR)
2.8. Statistical Analysis
3. Results
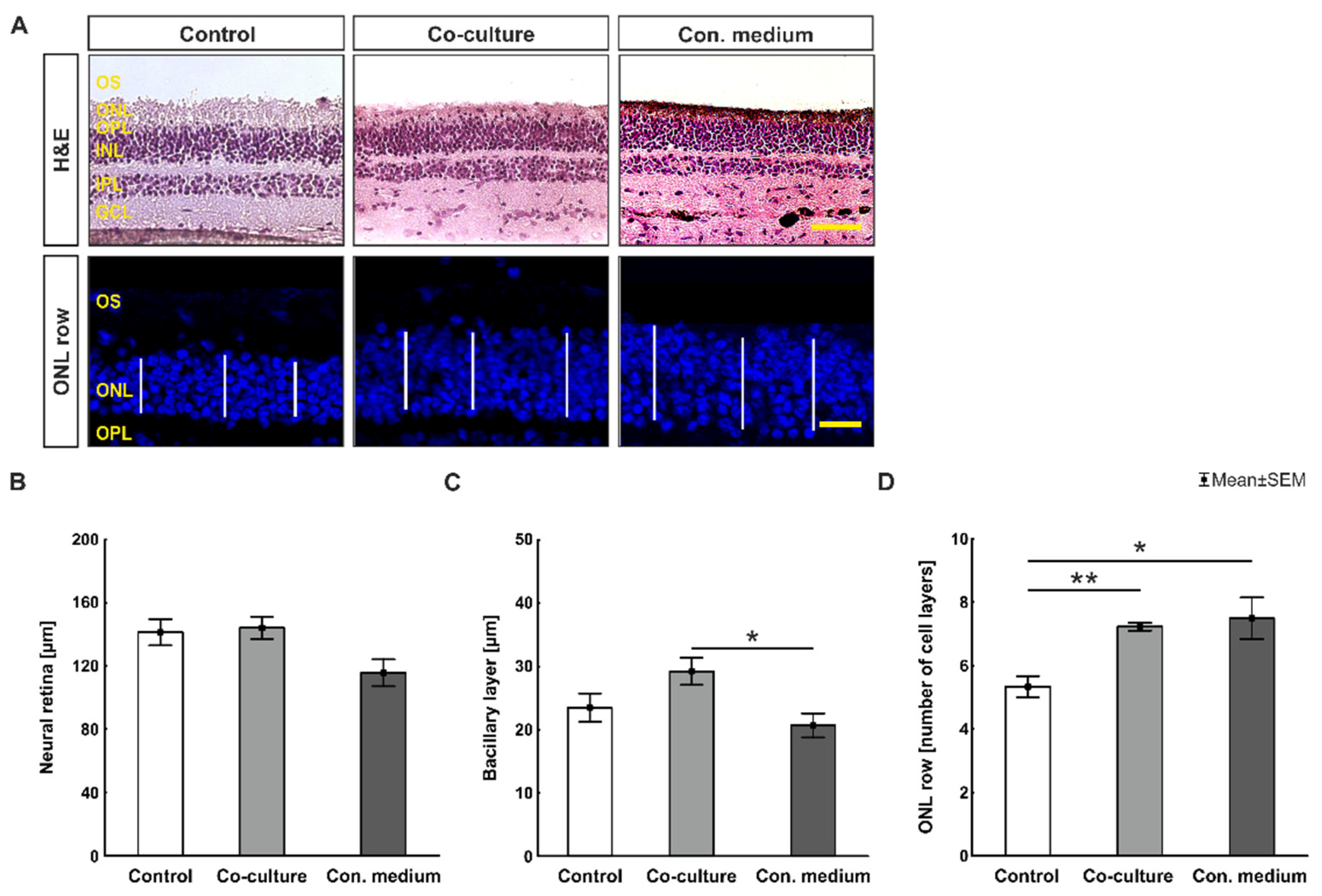
3.1. The Presence of ppRPE Cells Preserved the Neuroretina Most Efficiently in Culture
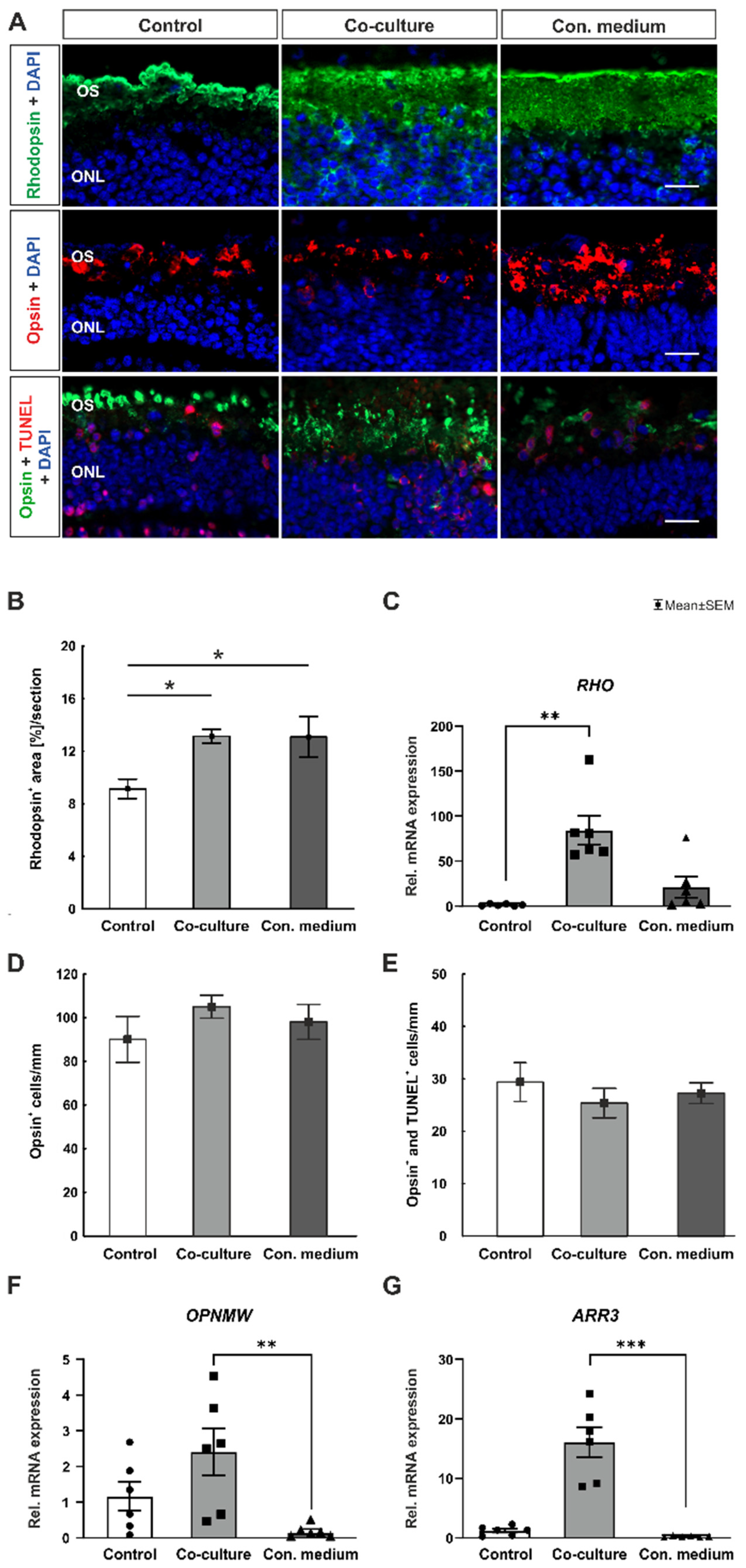
3.2. Presence of ppRPE Cells Maintains Rods
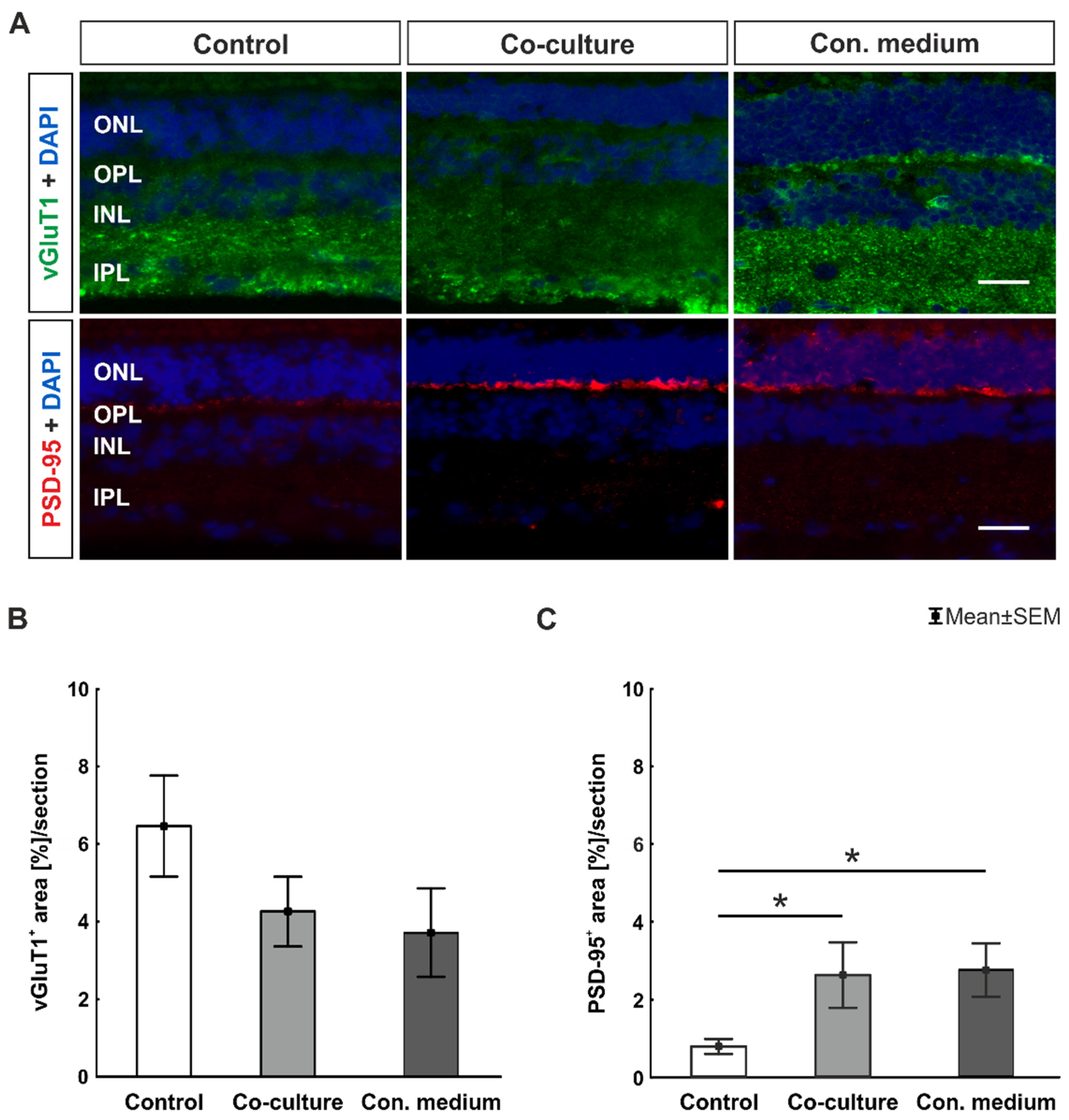
3.3. Co-Cultivation with ppRPE or Conditioned Medium Led to Unaltered Synaptic Density
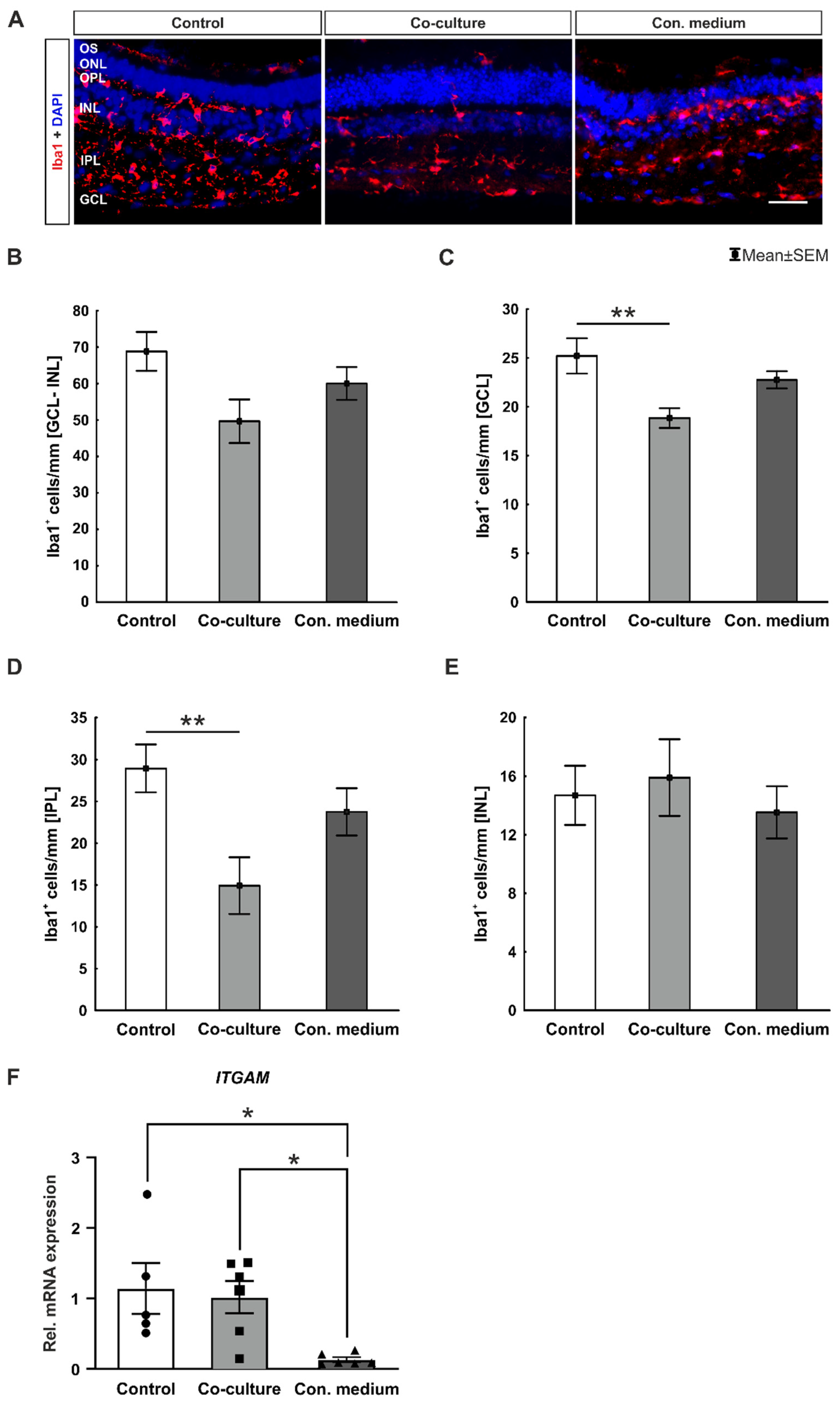
3.4. No Microglia Activation
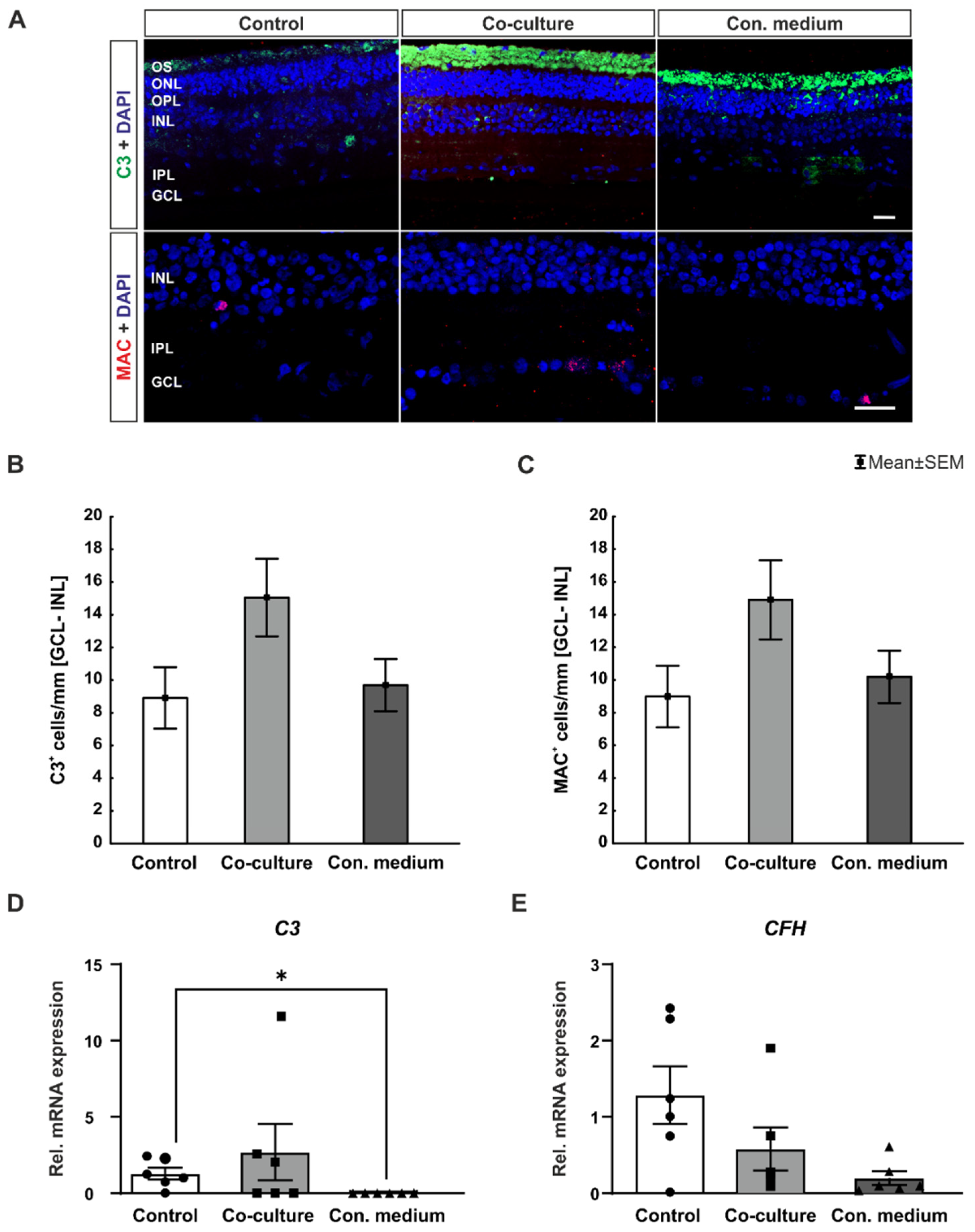
3.5. No Complement System Activation In Vitro
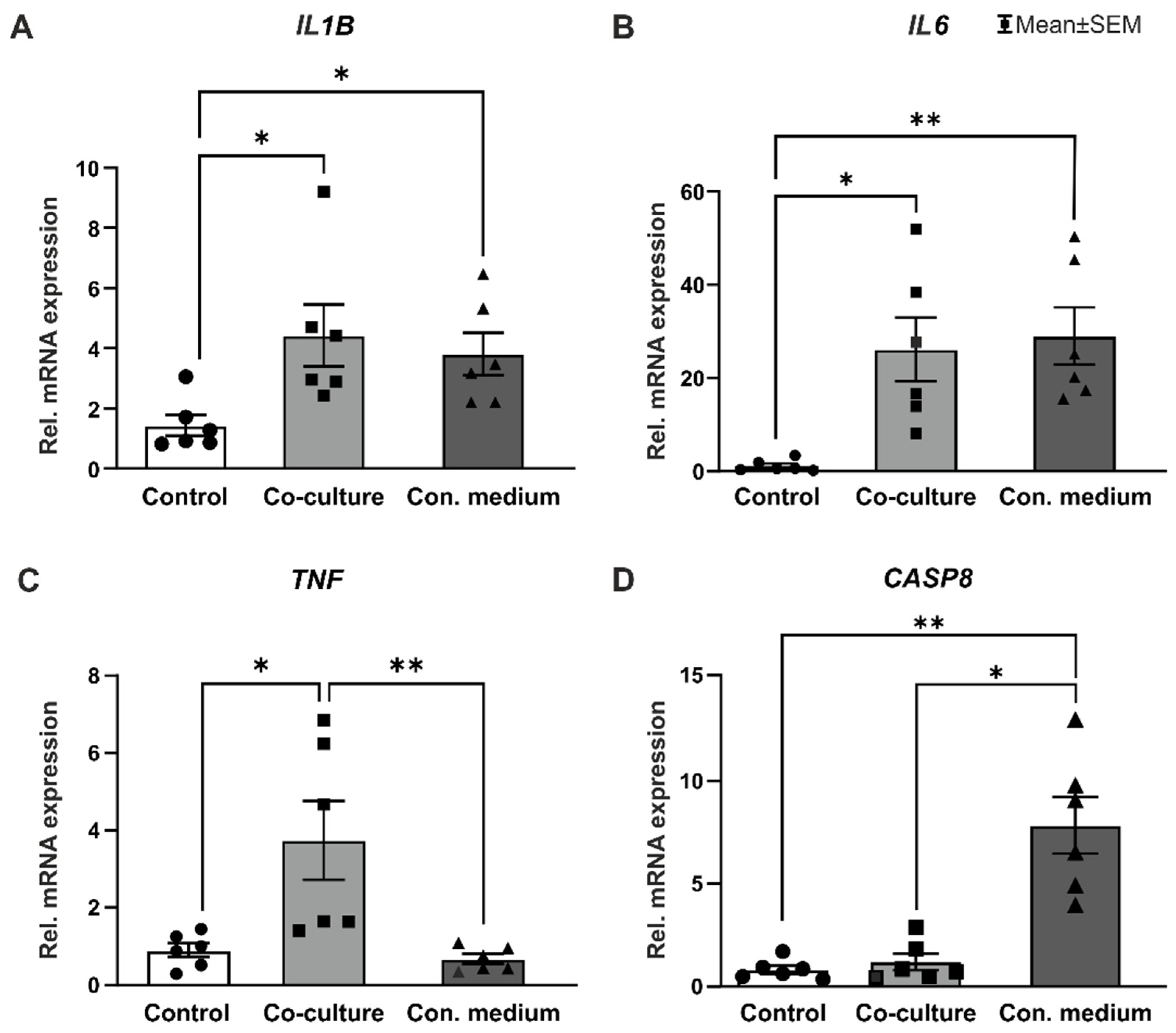
3.6. Pro-Inflammatory Cytokines were Upregulated in Co-Culture and Conditioned Medium Samples
4. Discussion
4.1. Best Preservation of Neuroretina in ppRPE Co-Cultivation
4.2. Altered Postsynaptic Transmission in Co-Culture and Conditioned Medium Conditions
4.3. Lower Microglia Numbers in Co-Culture Retinae
4.4. No Detectable Complement System Activation
4.5. Pro-Inflammatory Cytokines Are Upregulated in Co-Cultivation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchie, H.; Roser, M. Age Structure. Our World Data, 2019; in press. [Google Scholar]
- Klein, R.; Klein, B.E.; Linton, K.L. Prevalence of age-related maculopathy. The Beaver Dam Eye Study. Ophthalmology 1992, 99, 933–943. [Google Scholar] [CrossRef]
- Snow, K.K.; Seddon, J.M. Do age-related macular degeneration and cardiovascular disease share common antecedents? Ophthal. Epidemiol. 1999, 6, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.P.; Edwards, R.; Elton, P.; Mitchell, P. Age related macular degeneration: Smoking entails major risk of blindness. BMJ 2003, 326, 1458–1459, author reply 1459–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopdar, A.; Chakravarthy, U.; Verma, D. Age related macular degeneration. BMJ 2003, 326, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, S. Retinitis pigmentosa: A brief overview. Indian J. Ophthalmol. 2011, 59, 343–346. [Google Scholar] [CrossRef]
- Tanna, P.; Strauss, R.W.; Fujinami, K.; Michaelides, M. Stargardt disease: Clinical features, molecular genetics, animal models and therapeutic options. Br. J. Ophthalmol. 2017, 101, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Ricca, A.M.; Han, I.C.; Sohn, E.H. Stargardt disease masquerades. Curr. Opin. Ophthalmol. 2021, 32, 214–224. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, J.; Li, D. Functions and Diseases of the Retinal Pigment Epithelium. Front. Pharmacol. 2021, 12, 727870. [Google Scholar] [CrossRef]
- Zarbin, M. Cell-Based Therapy for Degenerative Retinal Disease. Trends Mol. Med. 2016, 22, 115–134. [Google Scholar] [CrossRef]
- Pennesi, M.E.; Neuringer, M.; Courtney, R.J. Animal models of age related macular degeneration. Mol. Asp. Med. 2012, 33, 487–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, M.J.; Smith, P.J.; Samuelson, D.A.; MacKay, E.O. Photoreceptor density of the domestic pig retina. Vet. Ophthalmol. 1999, 2, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Middleton, S. Porcine ophthalmology. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, I.; Martin, R.; Ussa, F.; Fernandez-Bueno, I. The parameters of the porcine eyeball. Graefes Arch. Clin. Exp. Ophthalmol. 2011, 249, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.; Reinehr, S.; Gammel, M.R.; Greulich, A.; Hurst, J.; Dick, H.B.; Schnichels, S.; Joachim, S.C. Novel Porcine Retina Cultivation Techniques Provide Improved Photoreceptor Preservation. Front. Neurosci. 2020, 14, 556700. [Google Scholar] [CrossRef]
- Steinberg, R.H. Interactions between the retinal pigment epithelium and the neural retina. Doc. Ophthalmol. 1985, 60, 327–346. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.S.; Edelman, J.L. Active ion transport pathways in the bovine retinal pigment epithelium. J. Physiol. 1990, 424, 283–300. [Google Scholar] [CrossRef]
- Hamann, S. Molecular mechanisms of water transport in the eye. Int. Rev. Cytol. 2002, 215, 395–431. [Google Scholar] [CrossRef]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [Green Version]
- Baehr, W.; Wu, S.M.; Bird, A.C.; Palczewski, K. The retinoid cycle and retina disease. Vis. Res. 2003, 43, 2957–2958. [Google Scholar] [CrossRef]
- Schnichels, S.; Kiebler, T.; Hurst, J.; Maliha, A.M.; Loscher, M.; Dick, H.B.; Bartz-Schmidt, K.U.; Joachim, S.C. Retinal Organ Cultures as Alternative Research Models. Altern. Lab. Anim. 2019, 47, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, S.; Hurst, J.; Jashari, A.; Ahrens, K.; Tsai, T.; Wunderlich, I.M.; Dick, H.B.; Joachim, S.C.; Schnichels, S. The novel induction of retinal ganglion cell apoptosis in porcine organ culture by NMDA—An opportunity for the replacement of animals in experiments. Altern. Lab. Anim. 2016, 44, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Hurst, J.; Kuehn, S.; Jashari, A.; Tsai, T.; Bartz-Schmidt, K.U.; Schnichels, S.; Joachim, S.C. A novel porcine ex vivo retina culture model for oxidative stress induced by H2O2. Altern. Lab. Anim. 2017, 45, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.; Safaei, A.; Vogt, P.A.; Gammel, M.R.; Dick, H.B.; Schnichels, S.; Joachim, S.C. Coculture of ARPE-19 Cells and Porcine Neural Retina as an Ex Vivo Retinal Model. Altern. Lab. Anim. 2022, 50, 27–44. [Google Scholar] [CrossRef]
- Whelan, J.P.; McGinnis, J.F. Light-dependent subcellular movement of photoreceptor proteins. J. Neurosci. Res. 1988, 20, 263–270. [Google Scholar] [CrossRef]
- Zhu, X.; Li, A.; Brown, B.; Weiss, E.R.; Osawa, S.; Craft, C.M. Mouse cone arrestin expression pattern: Light induced translocation in cone photoreceptors. Mol. Vis. 2002, 8, 462–471. [Google Scholar]
- Mangini, N.J.; Pepperberg, D.R. Immunolocalization of 48K in rod photoreceptors. Light and ATP increase OS labeling. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1221–1234. [Google Scholar]
- Zhang, H.; Cuenca, N.; Ivanova, T.; Church-Kopish, J.; Frederick, J.M.; MacLeish, P.R.; Baehr, W. Identification and light-dependent translocation of a cone-specific antigen, cone arrestin, recognized by monoclonal antibody 7G6. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2858–2867. [Google Scholar] [CrossRef] [Green Version]
- Dosemeci, A.; Weinberg, R.J.; Reese, T.S.; Tao-Cheng, J.H. The Postsynaptic Density: There Is More than Meets the Eye. Front. Synaptic Neurosci. 2016, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.; Fremeau, R.T., Jr.; Duncan, J.L.; Renteria, R.C.; Yang, H.; Hua, Z.; Liu, X.; LaVail, M.M.; Edwards, R.H.; Copenhagen, D.R. Vesicular glutamate transporter 1 is required for photoreceptor synaptic signaling but not for intrinsic visual functions. J. Neurosci. 2007, 27, 7245–7255. [Google Scholar] [CrossRef] [Green Version]
- Kourtzelis, I.; Mitroulis, I.; von Renesse, J.; Hajishengallis, G.; Chavakis, T. From leukocyte recruitment to resolution of inflammation: The cardinal role of integrins. J. Leukoc. Biol. 2017, 102, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarks, S.H. Ageing and degeneration in the macular region: A clinico-pathological study. Br. J. Ophthalmol. 1976, 60, 324–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curcio, C.A. Soft Drusen in Age-Related Macular Degeneration: Biology and Targeting Via the Oil Spill Strategies. Investig. Ophthalmol. Vis. Sci. 2018, 59, AMD160–AMD181. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Zhao, M.; Tsai, S.H.; Burkes, W.L.; Potts, L.B.; Xu, W.; Payne, H.R.; Hein, T.W.; Kuo, L.; Rosa, R.H., Jr. Correlation of spectral domain optical coherence tomography with histology and electron microscopy in the porcine retina. Exp. Eye Res. 2018, 177, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, A.; Hicks, D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Exp. Eye Res. 2002, 74, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef]
- Simoens, P.; De Schaepdrijver, L.; Lauwers, H. Morphologic and clinical study of the retinal circulation in the miniature pig. A: Morphology of the retinal microvasculature. Exp. Eye Res. 1992, 54, 965–973. [Google Scholar] [CrossRef]
- Ghareeb, A.E.; Lako, M.; Steel, D.H. Coculture techniques for modeling retinal development and disease, and enabling regenerative medicine. Stem Cells Transl. Med. 2020, 9, 1531–1548. [Google Scholar] [CrossRef]
- Di Lauro, S.; Rodriguez-Crespo, D.; Gayoso, M.J.; Garcia-Gutierrez, M.T.; Pastor, J.C.; Srivastava, G.K.; Fernandez-Bueno, I. A novel coculture model of porcine central neuroretina explants and retinal pigment epithelium cells. Mol. Vis. 2016, 22, 243–253. [Google Scholar]
- Kaempf, S.; Walter, P.; Salz, A.K.; Thumann, G. Novel organotypic culture model of adult mammalian neurosensory retina in co-culture with retinal pigment epithelium. J. Neurosci. Methods 2008, 173, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Mohlin, C.; Sandholm, K.; Kvanta, A.; Ekdahl, K.N.; Johansson, K. A model to study complement involvement in experimental retinal degeneration. Upsala J. Med. Sci. 2018, 123, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Mohlin, C.; Delbro, D.; Kvanta, A.; Johansson, K. Evaluation of Congo Red Staining in Degenerating Porcine Photoreceptors In Vitro: Protective Effects by Structural and Trophic Support. J. Histochem. Cytochem. 2018, 66, 631–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolomeyer, A.M.; Sugino, I.K.; Zarbin, M.A. Characterization of conditioned media collected from cultured adult versus fetal retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5973–5986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolomeyer, A.M.; Sugino, I.K.; Zarbin, M.A. Characterization of the effects of retinal pigment epithelium-conditioned media on porcine and aged human retina. Graefes Arch. Clin. Exp. Ophthalmol. 2013, 251, 1515–1528. [Google Scholar] [CrossRef]
- Johnson, E.C.; Jia, L.; Cepurna, W.O.; Doser, T.A.; Morrison, J.C. Global changes in optic nerve head gene expression after exposure to elevated intraocular pressure in a rat glaucoma model. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3161–3177. [Google Scholar] [CrossRef]
- Johnson, J.; Tian, N.; Caywood, M.S.; Reimer, R.J.; Edwards, R.H.; Copenhagen, D.R. Vesicular neurotransmitter transporter expression in developing postnatal rodent retina: GABA and glycine precede glutamate. J. Neurosci. 2003, 23, 518–529. [Google Scholar] [CrossRef]
- Sherry, D.M.; Wang, M.M.; Bates, J.; Frishman, L.J. Expression of vesicular glutamate transporter 1 in the mouse retina reveals temporal ordering in development of rod vs. cone and ON vs. OFF circuits. J. Comp. Neurol. 2003, 465, 480–498. [Google Scholar] [CrossRef]
- Koulen, P.; Fletcher, E.L.; Craven, S.E.; Bredt, D.S.; Wassle, H. Immunocytochemical localization of the postsynaptic density protein PSD-95 in the mammalian retina. J. Neurosci. 1998, 18, 10136–10149. [Google Scholar] [CrossRef]
- Blackmon, S.M.; Peng, Y.W.; Hao, Y.; Moon, S.J.; Oliveira, L.B.; Tatebayashi, M.; Petters, R.M.; Wong, F. Early loss of synaptic protein PSD-95 from rod terminals of rhodopsin P347L transgenic porcine retina. Brain Res. 2000, 885, 53–61. [Google Scholar] [CrossRef]
- Mohlin, C.; Taylor, L.; Ghosh, F.; Johansson, K. Autophagy and ER-stress contribute to photoreceptor degeneration in cultured adult porcine retina. Brain Res. 2014, 1585, 167–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, K.; Svensson, L.A.; Mohlin, C. Morphological analyzes of microglia heterogeneity and dynamics during photoreceptor degeneration in vitro: Presumptive dark microglia in porcine retina. Exp. Eye Res. 2020, 200, 108217. [Google Scholar] [CrossRef] [PubMed]
- Wake, H.; Moorhouse, A.J.; Jinno, S.; Kohsaka, S.; Nabekura, J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J. Neurosci. 2009, 29, 3974–3980. [Google Scholar] [CrossRef] [Green Version]
- Stein-Streilein, J. Mechanisms of immune privilege in the posterior eye. Int. Rev. Immunol. 2013, 32, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.V.; Xu, H.; Kuffova, L.; Dick, A.D.; McMenamin, P.G. Dendritic cell physiology and function in the eye. Immunol. Rev. 2010, 234, 282–304. [Google Scholar] [CrossRef] [PubMed]
- Karlstetter, M.; Scholz, R.; Rutar, M.; Wong, W.T.; Provis, J.M.; Langmann, T. Retinal microglia: Just bystander or target for therapy? Prog. Retin. Eye Res. 2015, 45, 30–57. [Google Scholar] [CrossRef]
- Okunuki, Y.; Mukai, R.; Pearsall, E.A.; Klokman, G.; Husain, D.; Park, D.H.; Korobkina, E.; Weiner, H.L.; Butovsky, O.; Ksander, B.R.; et al. Microglia inhibit photoreceptor cell death and regulate immune cell infiltration in response to retinal detachment. Proc. Natl. Acad. Sci. USA 2018, 115, E6264–E6273. [Google Scholar] [CrossRef] [Green Version]
- Silverman, S.M.; Ma, W.; Wang, X.; Zhao, L.; Wong, W.T. C3- and CR3-dependent microglial clearance protects photoreceptors in retinitis pigmentosa. J. Exp. Med. 2019, 216, 1925–1943. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, E.; Delaney, C.; Campbell, M.; Doyle, S.L. Minocycline suppresses disease-associated microglia (DAM) in a model of photoreceptor cell degeneration. Exp. Eye Res. 2022, 217, 108953. [Google Scholar] [CrossRef]
- Sasaki, M.; Ozawa, Y.; Kurihara, T.; Kubota, S.; Yuki, K.; Noda, K.; Kobayashi, S.; Ishida, S.; Tsubota, K. Neurodegenerative influence of oxidative stress in the retina of a murine model of diabetes. Diabetologia 2010, 53, 971–979. [Google Scholar] [CrossRef] [Green Version]
- Hurst, J.; Mueller-Buehl, A.M.; Hofmann, L.; Kuehn, S.; Herms, F.; Schnichels, S.; Joachim, S.C. iNOS-inhibitor driven neuroprotection in a porcine retina organ culture model. J. Cell. Mol. Med. 2020, 24, 4312–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller-Buehl, A.M.; Tsai, T.; Hurst, J.; Theiss, C.; Peters, L.; Hofmann, L.; Herms, F.; Kuehn, S.; Schnichels, S.; Joachim, S.C. Reduced Retinal Degeneration in an Oxidative Stress Organ Culture Model through an iNOS-Inhibitor. Biology 2021, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.N.; Ayyagari, R. Complement factor H: Spatial and temporal expression and localization in the eye. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4091–4097. [Google Scholar] [CrossRef] [Green Version]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoh Kam, J.; Lenassi, E.; Malik, T.H.; Pickering, M.C.; Jeffery, G. Complement component C3 plays a critical role in protecting the aging retina in a murine model of age-related macular degeneration. Am. J. Pathol. 2013, 183, 480–492. [Google Scholar] [CrossRef]
- Maliha, A.M.; Kuehn, S.; Hurst, J.; Herms, F.; Fehr, M.; Bartz-Schmidt, K.U.; Dick, H.B.; Joachim, S.C.; Schnichels, S. Diminished apoptosis in hypoxic porcine retina explant cultures through hypothermia. Sci. Rep. 2019, 9, 4898. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Chen, M. Targeting the complement system for the management of retinal inflammatory and degenerative diseases. Eur. J. Pharmacol. 2016, 787, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A. The retinal Pigment Epithelium in the immune Regulation of the Retina. In Recent Advances in eye Research; Nova Science Publishers: Hauppauge, NY, USA, 2015; pp. 31–52. [Google Scholar]
- AnandBabu, K.; Sen, P.; Angayarkanni, N. Oxidized LDL, homocysteine, homocysteine thiolactone and advanced glycation end products act as pro-oxidant metabolites inducing cytokine release, macrophage infiltration and pro-angiogenic effect in ARPE-19 cells. PLoS ONE 2019, 14, e0216899. [Google Scholar] [CrossRef] [Green Version]
- Demircan, N.; Safran, B.G.; Soylu, M.; Ozcan, A.A.; Sizmaz, S. Determination of vitreous interleukin-1 (IL-1) and tumour necrosis factor (TNF) levels in proliferative diabetic retinopathy. Eye 2006, 20, 1366–1369. [Google Scholar] [CrossRef] [Green Version]
- Pollreisz, A.; Sacu, S.; Eibenberger, K.; Funk, M.; Kivaranovic, D.; Zlabinger, G.J.; Georgopoulos, M.; Schmidt-Erfurth, U. Extent of Detached Retina and Lens Status Influence Intravitreal Protein Expression in Rhegmatogenous Retinal Detachment. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5493–5502. [Google Scholar] [CrossRef] [Green Version]
- Symeonidis, C.; Androudi, S.; Tsaousis, K.T.; Tsinopoulos, I.; Brazitikos, P.; Diza, E.; Dimitrakos, S.A. Comparison of interleukin IL-6 levels in the subretinal fluid and the vitreous during rhegmatogenous retinal detachment. Cytokine 2012, 57, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.; Kuehn, S.; Tsiampalis, N.; Vu, M.K.; Kakkassery, V.; Stute, G.; Dick, H.B.; Joachim, S.C. Anti-inflammatory cytokine and angiogenic factors levels in vitreous samples of diabetic retinopathy patients. PLoS ONE 2018, 13, e0194603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, T.S. Contributions of inflammatory processes to the development of the early stages of diabetic retinopathy. Exp. Diabetes Res. 2007, 2007, 95103. [Google Scholar] [CrossRef]
- Todd, L.; Palazzo, I.; Suarez, L.; Liu, X.; Volkov, L.; Hoang, T.V.; Campbell, W.A.; Blackshaw, S.; Quan, N.; Fischer, A.J. Reactive microglia and IL1beta/IL-1R1-signaling mediate neuroprotection in excitotoxin-damaged mouse retina. J. Neuroinflamm. 2019, 16, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ten Berge, J.C.; Fazil, Z.; van den Born, I.; Wolfs, R.C.W.; Schreurs, M.W.J.; Dik, W.A.; Rothova, A. Intraocular cytokine profile and autoimmune reactions in retinitis pigmentosa, age-related macular degeneration, glaucoma and cataract. Acta Ophthalmol. 2019, 97, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Abcouwer, S.F. Angiogenic Factors and Cytokines in Diabetic Retinopathy. J. Clin. Cell. Immunol. 2013, 11 (Suppl. 1), 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.J.; Zelinka, C.; Milani-Nejad, N. Reactive retinal microglia, neuronal survival, and the formation of retinal folds and detachments. Glia 2015, 63, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Chong, D.Y.; Boehlke, C.S.; Zheng, Q.D.; Zhang, L.; Han, Y.; Zacks, D.N. Interleukin-6 as a photoreceptor neuroprotectant in an experimental model of retinal detachment. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3193–3200. [Google Scholar] [CrossRef]
- Al-Gayyar, M.M.; Elsherbiny, N.M. Contribution of TNF-alpha to the development of retinal neurodegenerative disorders. Eur. Cytokine Netw. 2013, 24, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, L.; Lucius, R.; Roider, J.; Klettner, A. Interaction of inflammatorily activated retinal pigment epithelium with retinal microglia and neuronal cells. Exp. Eye Res. 2020, 199, 108167. [Google Scholar] [CrossRef]
- Hughes, M.A.; Powley, I.R.; Jukes-Jones, R.; Horn, S.; Feoktistova, M.; Fairall, L.; Schwabe, J.W.; Leverkus, M.; Cain, K.; MacFarlane, M. Co-operative and Hierarchical Binding of c-FLIP and Caspase-8: A Unified Model Defines How c-FLIP Isoforms Differentially Control Cell Fate. Mol. Cell 2016, 61, 834–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.; Leung, L.; Carroll, W.L.; Schibler, K.R. Regulation of interleukin-10 gene expression: Possible mechanisms accounting for its upregulation and for maturational differences in its expression by blood mononuclear cells. Blood 1997, 89, 4112–4119. [Google Scholar] [CrossRef] [PubMed]
- Petrilli, V.; Papin, S.; Tschopp, J. The inflammasome. Curr. Biol. 2005, 15, R581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheu, S.; Stetson, D.B.; Reinhardt, R.L.; Leber, J.H.; Mohrs, M.; Locksley, R.M. Activation of the integrated stress response during T helper cell differentiation. Nat. Immunol. 2006, 7, 644–651. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Secondary Antibodies | ||||||
|---|---|---|---|---|---|---|---|
| Antibody | Catalogue Number/Clone ID | Company | Dilution | Antibody | Catalogue Number | Company | Dilution |
| Anti-C3 | CL7334Ap | Cedarlane | 1:500 | Goat anti-rabbit Alexa Fluor 488 | A11008 | Invitrogen | 1:500 |
| Anti-C5b-9 (MAC) | HM3033 | Biozol | 1:100 | Donkey anti-mouse Alexa Fluor 555 | ab150106 | Abcam | 1:500 |
| Anti-Iba1 | 234006 | Synaptic System | 1:400 | Donkey anti-chicken Cy3 | AP194C | Milipore | 1:500 |
| Anti-iNOS | PA1-036 | Thermo Fisher | 1:50 | Donkey anti-rabbit Alexa Fluor 488 | 711-547-003 | Jackson Immuno Research | 1:600 |
| Anti-M/L opsin | AB5405 | Millipore | 1:1200 | Donkey anti-rabbit Alexa Fluor 488 | 711-547-003 | Jackson Immuno Research | 1:600 |
| Anti-PSD-95 | CP35 | Calbiochem | 1:300 | Donkey anti-mouse Alexa Fluor 555 | ab150106 | Abcam | 1:500 |
| Anti-rhodopsin | ab3267 | Abcam | 1:400 | Goat anti-mouse Alexa Fluor 488 | A-11029 | Invitrogen | 1:500 |
| Anti-vGluT1 | 135316 | Synaptic System | 1:100 | Donkey anti-chicken Alexa Fluor 488 | 703-545-155 | Jackson Immuno Research | 1:500 |
| Gene | Oligonucleotides 5′ 3′ | GenBank Accession Number | Amplicon Size |
|---|---|---|---|
| ACTB for ACTB rev | CTCTTCCAGCCTTCCTTC GGGCAGTGATCTCTTTCT | XM_021086047.1 | 178 |
| ARR3 for ARR3 rev | TGACAACTGCGAGAAACAGG CACAGGACACCATCAGGTTG | NM_214345.1 | 157 |
| C3 for C3 rev | ACAAATTGACCCAGCGTAGG GCACGTCCTTGCTGTACTGA | NM_214009.1 | 285 |
| CASP8 for CASP8 rec | GCCCAGATCTCTGCCTACAG CAGGGCCTTGTTGATTTGTT | XM_021074710.1 | 227 |
| CFH for CFH rev | GTGTGTGGTGAAGACGGATG GGGTGGAGCACAGGATTTTA | NM_214281.2 | 248 |
| GFAP for GFAP rev | GGAGAAGCCTTTGCTACACG TCTTCACTCTGCCTGGGTCT | NM_001244397.1 | 170 |
| IL1B for IL1B rev | CCAAAGAGGGACATGGAGAA TTATATCTTGGCGGCCTTTG | XM_021085847.1 | 159 |
| IL6 for IL6 rev | CACCAGGAACGAAAGAGAGC GTTTTGTCCGGAGAGGTGAA | NM_214399.1 | 193 |
| ITGAM for ITGAM rev | AGAAGGAGACACCCAGAGCA GTAGGACAATGGGCGTCACT | XM_021086380.1 | 169 |
| NOS2 for NOS2 rev | TGTTCAGCTGTGCCTTCAAC CAGAACTGGGGGTACATGCT | NM_001143690.1 | 175 |
| OPNMW for OPNMW rev | GGGGAGCATCTTCACCTACA GATGATGGTCTCTGCCAGGT | NM_001011506.1 | 244 |
| RHO for RHO rev | TCCAGGTACATCCCAGAAGG GCTGCCCATAGCAGAAGAAG | NM_214221.1 | 151 |
| RLP4 for RLP4 rev | CAAGAGTAACTACAACCTTC GAACTCTACGATGAATCTTC | XM_005659862.3 | 164 |
| TNF for TNF rev | CCACCAACGTTTTCCTCACT CCAAAATAGACCTGCCCAGA | JF831365.1 | 296 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, N.; Safaei, A.; Hurst, J.; Vogt, P.A.; Dick, H.B.; Joachim, S.C.; Schnichels, S. Impact of Primary RPE Cells in a Porcine Organotypic Co-Cultivation Model. Biomolecules 2022, 12, 990. https://doi.org/10.3390/biom12070990
Wagner N, Safaei A, Hurst J, Vogt PA, Dick HB, Joachim SC, Schnichels S. Impact of Primary RPE Cells in a Porcine Organotypic Co-Cultivation Model. Biomolecules. 2022; 12(7):990. https://doi.org/10.3390/biom12070990
Chicago/Turabian StyleWagner, Natalie, Armin Safaei, José Hurst, Pia A. Vogt, H. Burkhard Dick, Stephanie C. Joachim, and Sven Schnichels. 2022. "Impact of Primary RPE Cells in a Porcine Organotypic Co-Cultivation Model" Biomolecules 12, no. 7: 990. https://doi.org/10.3390/biom12070990
APA StyleWagner, N., Safaei, A., Hurst, J., Vogt, P. A., Dick, H. B., Joachim, S. C., & Schnichels, S. (2022). Impact of Primary RPE Cells in a Porcine Organotypic Co-Cultivation Model. Biomolecules, 12(7), 990. https://doi.org/10.3390/biom12070990