
Hsp70 and Hsp110 Chaperones Promote Early Steps of Proteasome Assembly



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Media
2.2. Protein Extraction
2.3. Fractionation of Whole-Cell Extracts by Gel Filtration
2.4. Immunoaffinity Purification of the α4 Complex and Mass Spectrometry Analysis
2.5. Electrophoresis and Immunoblotting
3. Results
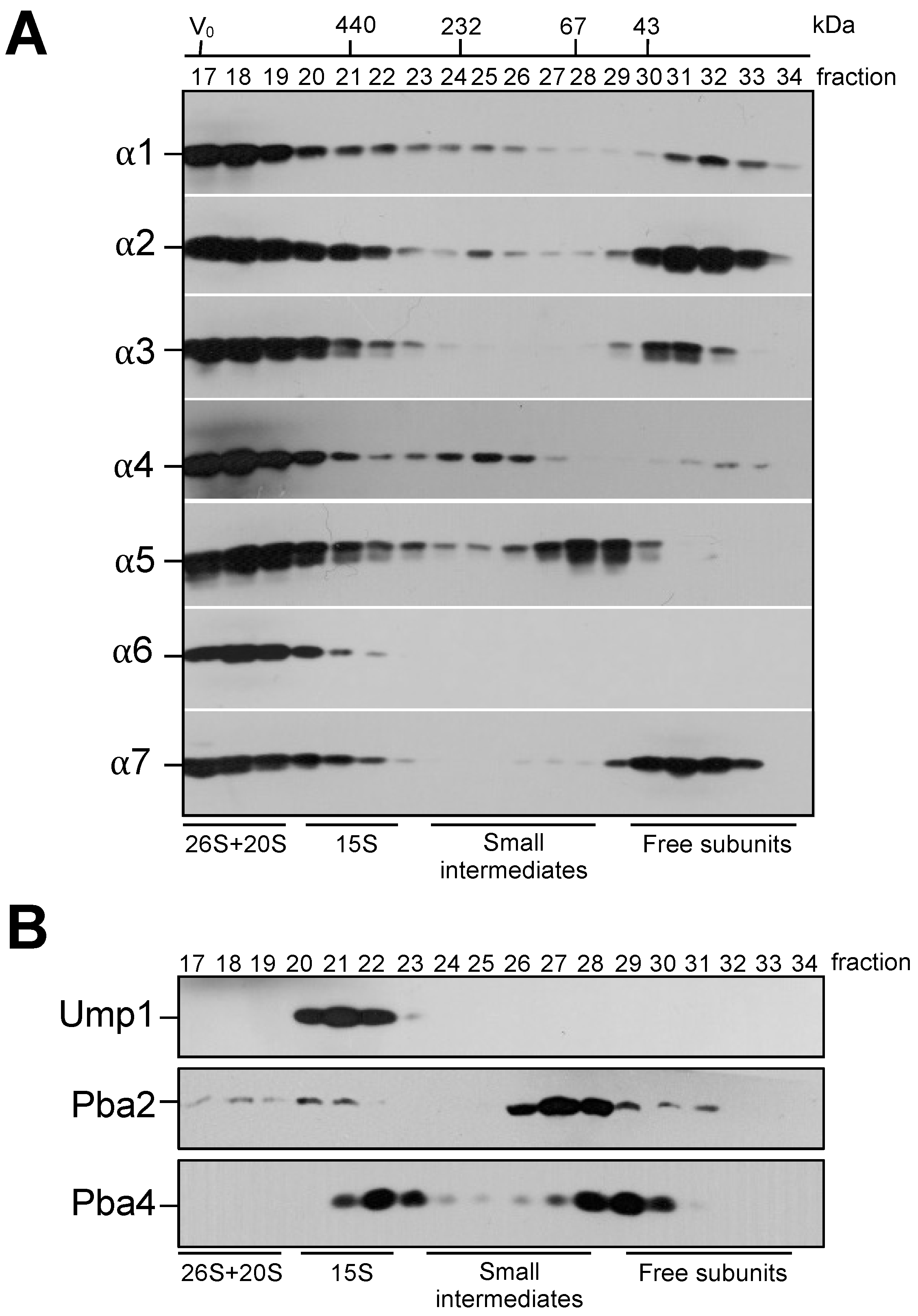
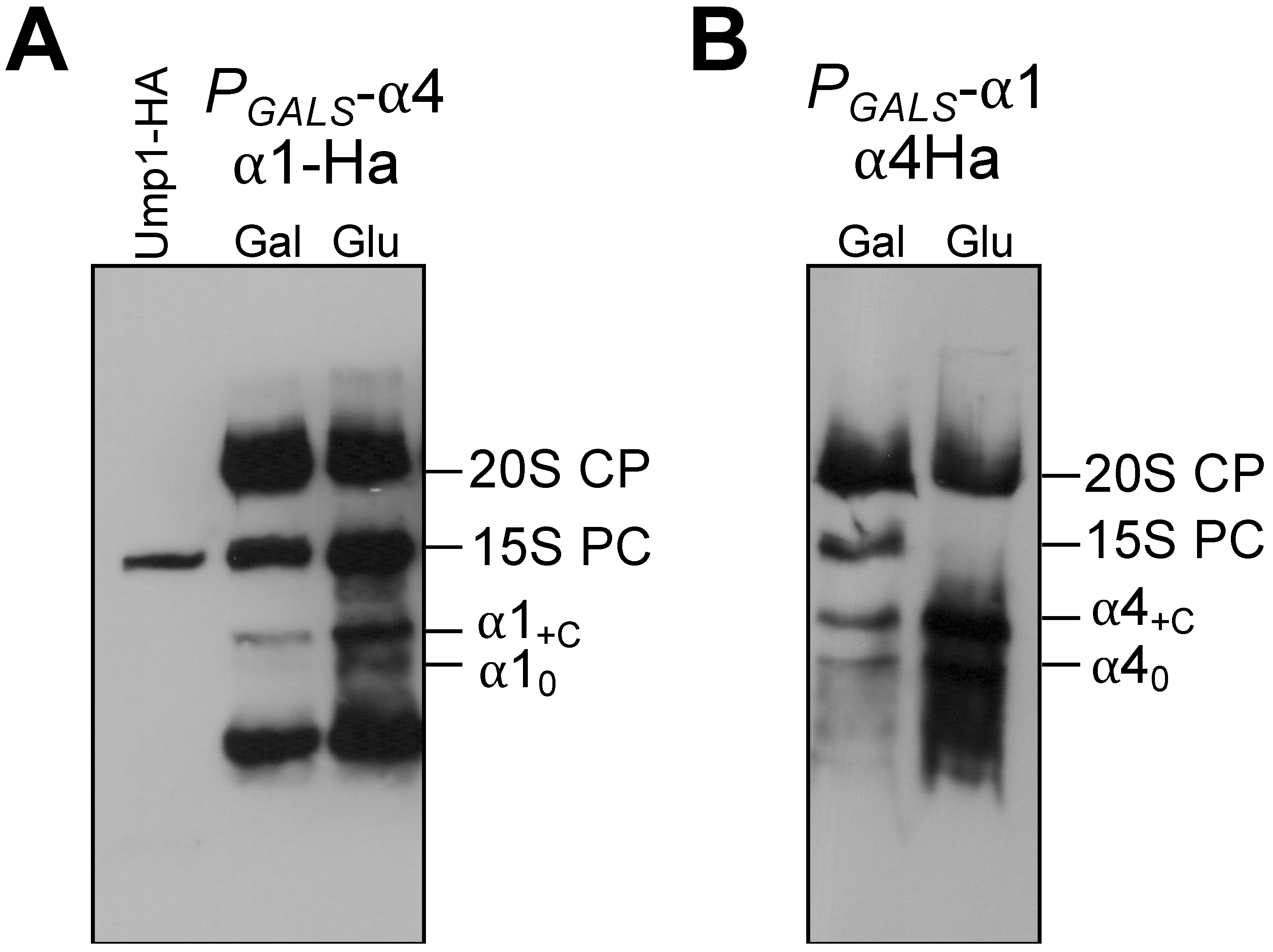
3.1. Identification of Small Intermediates Containing Either α1, α2, or α4 Subunits
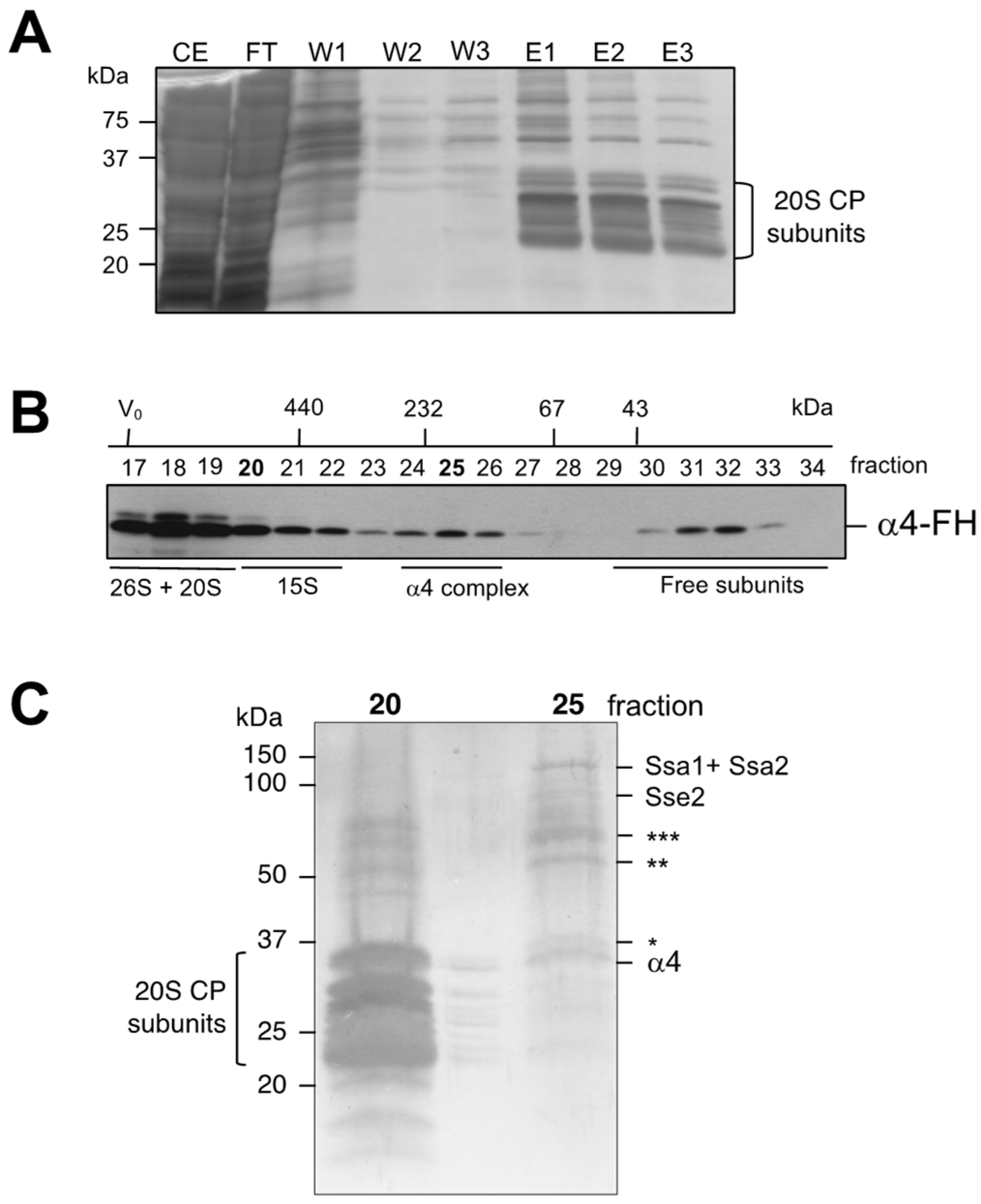
3.2. Isolation and Characterization of Complexes Containing Subunit α4
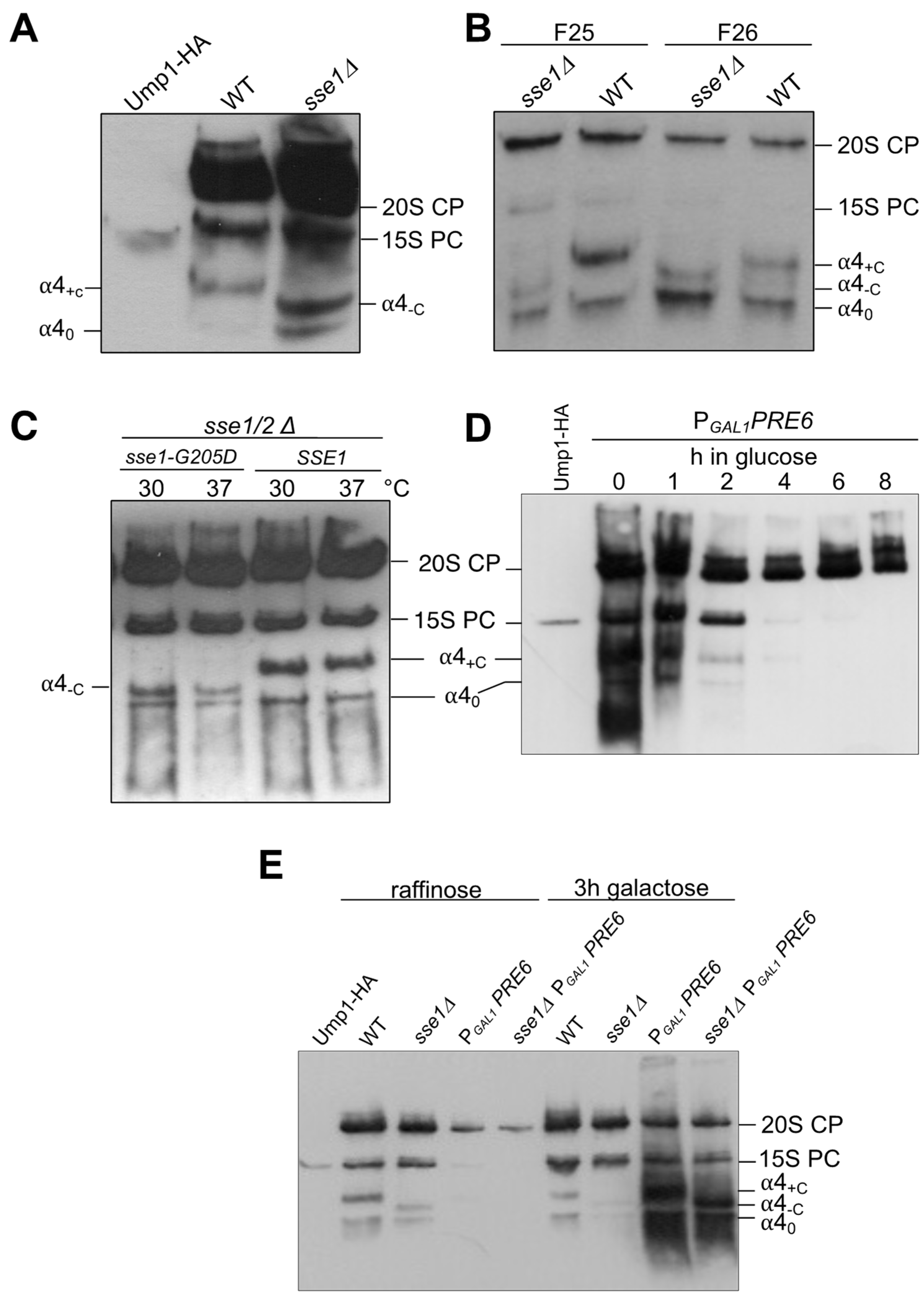
3.3. Effect of SSE1 Deletion on Small Complexes Containing α4 Separated by Native PAGE
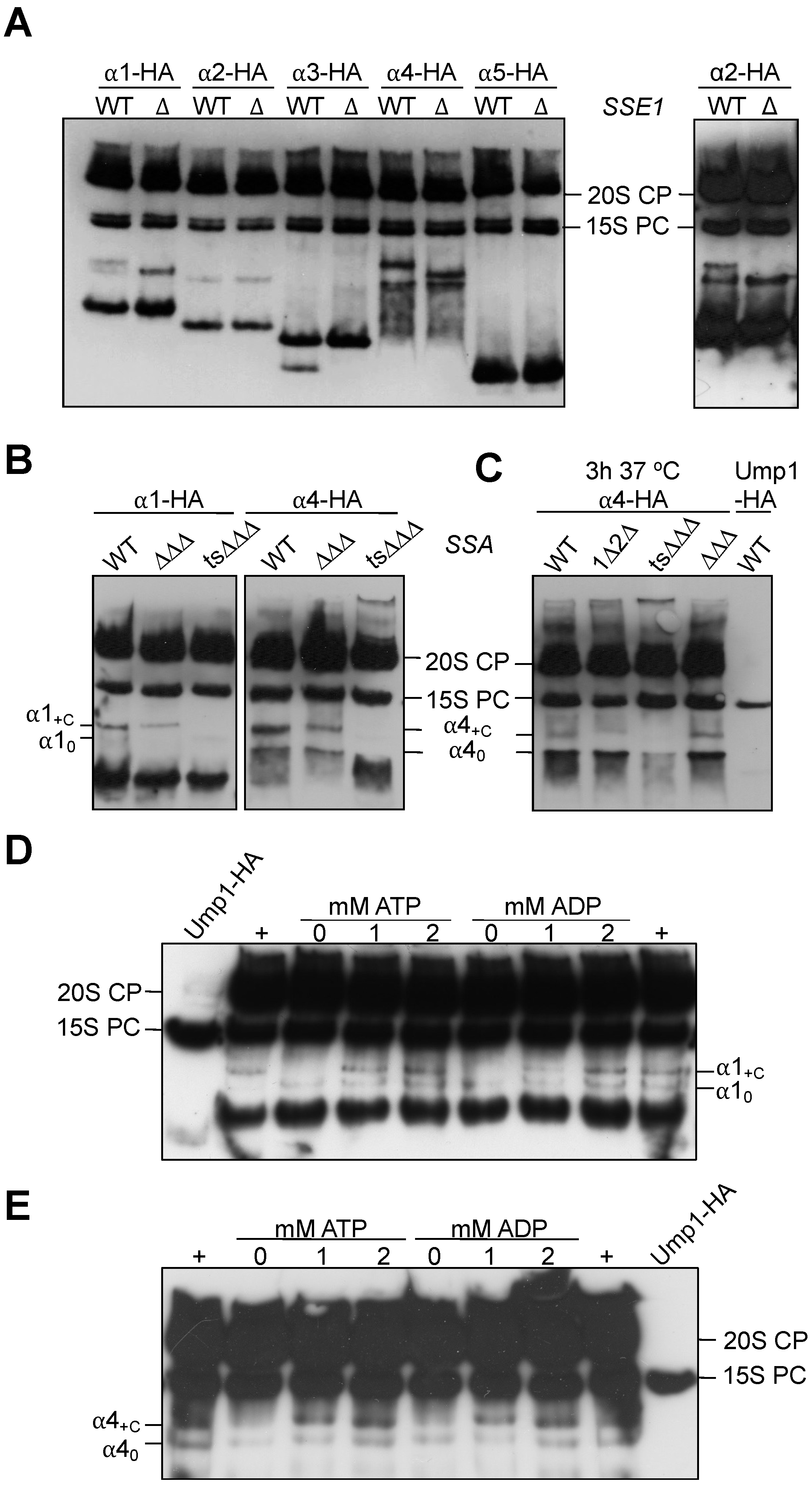
3.4. Effects of sse1 and ssa1-4 Mutations on Complexes Containing Other α Subunits
3.5. Formation of α1 Complexes Is Independent of α4 and Vice Versa
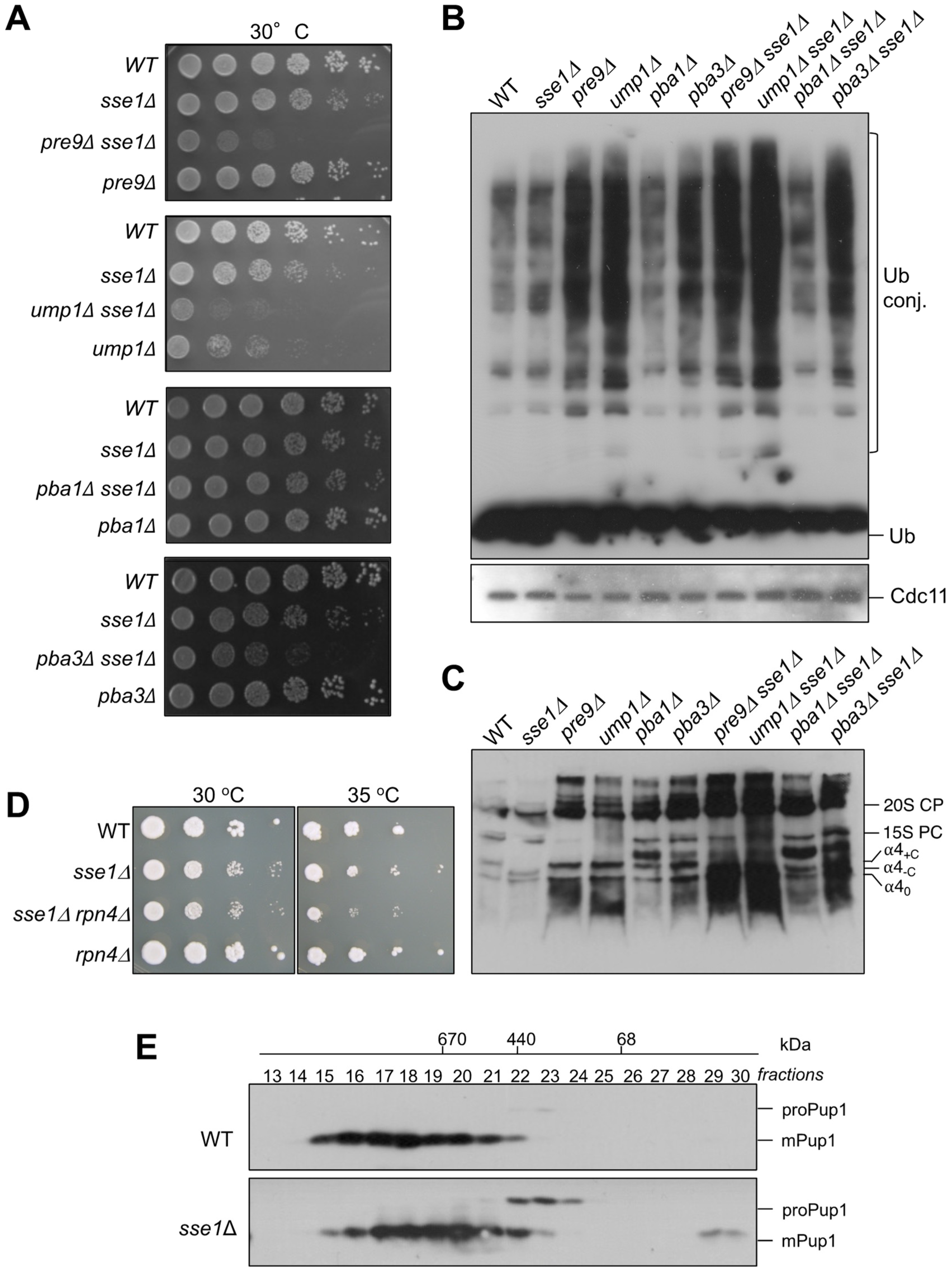
3.6. Genetic Interactions between sse1∆ and Mutations Affecting Proteasome Assembly
4. Discussion
4.1. α Subunits Initiate Assembly of Proteasome Core Particles without Forming a Heptameric α Ring Intermediate
4.2. The Role of Hsp70/Hsp110 Machinery in Early Proteasome Assembly Steps
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popovic, D.; Vucic, D.; Dikic, I. Ubiquitination in disease pathogenesis and treatment. Nat. Med. 2014, 20, 1242–1253. [Google Scholar] [CrossRef]
- Goetzke, C.C.; Ebstein, F.; Kallinich, T. Role of proteasomes in inflammation. J. Clin. Med. 2021, 10, 1783. [Google Scholar] [CrossRef] [PubMed]
- Finley, D.; Ulrich, H.D.; Sommer, T.; Kaiser, P. The ubiquitin-proteasome system of Saccharomyces cerevisiae. Genetics 2012, 192, 319–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Finley, D. Regulation of proteasome activity in health and disease. Biochim. Biophys. Acta 2014, 1843, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Sbardella, D.; Tundo, G.R.; Cunsolo, V.; Grasso, G.; Cascella, R.; Caputo, V.; Santoro, A.M.; Milardi, D.; Pecorelli, A.; Ciaccio, C.; et al. Defective proteasome biogenesis into skin fibroblasts isolated from Rett syndrome subjects with MeCP2 non-sense mutations. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165793. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.; Minis, A.; Lalazar, G.; Rodriguez, J.; Steller, H. PSMD5 Inactivation promotes 26S proteasome assembly during colorectal tumor progression. Cancer Res. 2018, 78, 3458–3468. [Google Scholar] [CrossRef] [Green Version]
- Shim, S.M.; Lee, W.J.; Kim, Y.; Chang, J.W.; Song, S.; Jung, Y.K. Role of S5b/PSMD5 in proteasome inhibition caused by TNF-α/NFκB in higher eukaryotes. Cell Rep. 2012, 2, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Funakoshi, M.; Tomko, R.J., Jr.; Kobayashi, H.; Hochstrasser, M. Multiple assembly chaperones govern biogenesis of the proteasome regulatory particle base. Cell 2009, 137, 887–899. [Google Scholar] [CrossRef] [Green Version]
- Ramos, P.C.; Dohmen, R.J. PACemakers of proteasome core particle assembly. Structure 2008, 16, 1296–1304. [Google Scholar] [CrossRef] [Green Version]
- Murata, S.; Yashiroda, H.; Tanaka, K. Molecular mechanisms of proteasome assembly. Nat. Rev. Mol. Cell Biol. 2009, 10, 104–115. [Google Scholar] [CrossRef]
- Tomko, R.J., Jr.; Hochstrasser, M. Molecular architecture and assembly of the eukaryotic proteasome. Annu. Rev. Biochem. 2013, 82, 415–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groll, M.; Ditzel, L.; Löwe, J.; Stock, D.; Bochtler, M.; Bartunik, H.D.; Huber, R. Structure of 20S proteasome from yeast at 2.4 A resolution. Nature 1997, 386, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Velichutina, I.; Connerly, P.L.; Arendt, C.S.; Li, X.; Hochstrasser, M. Plasticity in eucaryotic 20S proteasome ring assembly revealed by a subunit deletion in yeast. EMBO J. 2004, 23, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmanabhan, A.; Vuong, S.A.; Hochstrasser, M. Assembly of an evolutionarily conserved alternative proteasome isoform in human cells. Cell Rep. 2016, 14, 2962–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kniepert, A.; Groettrup, M. The unique functions of tissue-specific proteasomes. Trends Biochem. Sci. 2014, 39, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, P.C.; Höckendorff, J.; Johnson, E.S.; Varshavsky, A.; Dohmen, R.J. Ump1p is required for proper maturation of the 20S proteasome and becomes its substrate upon completion of the assembly. Cell 1998, 92, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kusmierczyk, A.R.; Wong, P.; Emili, A.; Hochstrasser, M. beta-Subunit appendages promote 20S proteasome assembly by overcoming an Ump1-dependent checkpoint. EMBO J. 2007, 26, 2339–2349. [Google Scholar] [CrossRef] [Green Version]
- Le Tallec, B.; Barrault, M.B.; Courbeyrette, R.; Guérois, R.; Marsolier-Kergoat, M.C.; Peyroche, A. 20S proteasome assembly is orchestrated by two distinct pairs of chaperones in yeast and in mammals. Mol. Cell 2007, 27, 660–674. [Google Scholar] [CrossRef]
- Kock, M.; Nunes, M.M.; Hemann, M.; Kube, S.; Dohmen, R.J.; Herzog, F.; Ramos, P.C.; Wendler, P. Proteasome assembly from 15S precursors involves major conformational changes and recycling of the Pba1-Pba2 chaperone. Nat. Commun. 2015, 6, 6123. [Google Scholar] [CrossRef] [Green Version]
- Schnell, H.M.; Walsh, R.M., Jr.; Rawson, S.; Kaur, M.; Bhanu, M.K.; Tian, G.; Prado, M.A.; Guerra-Moreno, A.; Paulo, J.A.; Gygi, S.P.; et al. Structures of chaperone-associated assembly intermediates reveal coordinated mechanisms of proteasome biogenesis. Nat. Struct. Mol. Biol. 2021, 28, 418–425. [Google Scholar] [CrossRef]
- Marques, A.J.; Glanemann, C.; Ramos, P.C.; Dohmen, R.J. The C-terminal extension of the beta7 subunit and activator complexes stabilize nascent 20 S proteasomes and promote their maturation. J. Biol. Chem. 2007, 282, 34869–34876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, J.; Ramos, P.C.; Dohmen, R.J. Interaction with the assembly chaperone Ump1 promotes incorporation of the β7 subunit into half-proteasome precursor complexes driving their dimerization. Biomolecules 2022, 12, 253. [Google Scholar] [CrossRef] [PubMed]
- Zwickl, P.; Kleinz, J.; Baumeister, W. Critical elements in proteasome assembly. Nat. Struct. Biol. 1994, 1, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Zühl, F.; Seemüller, E.; Golbik, R.; Baumeister, W. Dissecting the assembly pathway of the 20S proteasome. FEBS Lett. 1997, 24, 418189–418194. [Google Scholar] [CrossRef] [Green Version]
- Panfair, D.; Ramamurthy, A.; Kusmierczyk, A.R. Alpha-ring independent assembly of the 20S proteasome. Sci. Rep. 2015, 19, 13130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, Y.; Hendil, K.B.; Yashiroda, H.; Iemura, S.; Nagane, R.; Hioki, Y.; Natsume, T.; Tanaka, K.; Murata, S. A heterodimeric complex that promotes the assembly of mammalian 20S proteasomes. Nature 2005, 437, 1381–1385. [Google Scholar] [CrossRef]
- Hirano, Y.; Hayashi, H.; Iemura, S.; Hendil, K.B.; Niwa, S.; Kishimoto, T.; Kasahara, M.; Natsume, T.; Tanaka, K.; Murata, S. Cooperation of multiple chaperones required for the assembly of mammalian 20S proteasomes. Mol. Cell 2006, 24, 977–984. [Google Scholar] [CrossRef]
- Hammack, L.J.; Panfair, D.; Kusmierczyk, A.R. A novel proteasome assembly intermediate bypasses the need to form α-rings first. Biochem. Biophys. Res. Commun. 2020, 525, 107–112. [Google Scholar] [CrossRef]
- Dohmen, R.J.; Stappen, R.; McGrath, J.P.; Forrová, H.; Kolarov, J.; Goffeau, A.; Varshavsky, A. An essential yeast gene encoding a homolog of ubiquitin-activating enzyme. J. Biol. Chem. 1995, 270, 18099–18109. [Google Scholar] [CrossRef] [Green Version]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Gietz, R.D.; Sugino, A. New yeast-Escherichia coli shuttle vectors constructed with in vitro mutagenized yeast genes lacking six- base pair restriction sites. Gene 1988, 74, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Mumberg, D.; Müller, R.; Funk, M. Regulatable promoters of Saccharomyces cerevisiae: Comparison of transcriptional activity and their use for heterologous expression. Nucleic Acids Res. 1994, 22, 5767–5768. [Google Scholar] [CrossRef] [Green Version]
- Janke, C.; Magiera, M.M.; Rathfelder, N.; Taxis, C.; Reber, S.; Maekawa, H.; Moreno-Borchart, A.; Doenges, G.; Schwob, E.; Schiebel, E.; et al. A versatile toolbox for PCR-based tagging of yeast genes: New fluorescent proteins, more markers and promoter substitution cassettes. Yeast 2004, 21, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Knop, M.; Siegers, K.; Pereira, G.; Zachariae, W.; Winsor, B.; Nasmyth, K.; Schiebel, E. Epitope tagging of yeast genes using a PCR-based strategy: More tags and improved practical routines. Yeast 1999, 15, 963–972. [Google Scholar] [CrossRef]
- Yashiroda, H.; Mizushima, T.; Okamoto, K.; Kameyama, T.; Hayashi, H.; Kishimoto, T.; Niwa, S.; Kasahara, M.; Kurimoto, E.; Sakata, E.; et al. Crystal structure of a chaperone complex that contributes to the assembly of yeast 20S proteasomes. Nat. Struct. Mol. Biol. 2008, 15, 228–236. [Google Scholar] [CrossRef]
- Shaner, L.; Morano, K.A. All in the family: Atypical Hsp70 chaperones are conserved modulators of Hsp70 activity. Cell Stress Chaperones. 2007, 12, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P. Gymnastics of molecular chaperones. Mol. Cell 2010, 39, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Mukai, H.; Kuno, T.; Tanaka, H.; Hirata, D.; Miyakawa, T.; Tanaka, C. Isolation and characterization of SSE1 and SSE2, new members of the yeast HSP70 multigene family. Gene 1993, 132, 57–66. [Google Scholar] [CrossRef]
- Shaner, L.; Wegele, H.; Buchner, J.; Morano, K.A. The yeast Hsp110 Sse1 functionally interacts with the Hsp70 chaperones Ssa and Ssb. J. Biol. Chem. 2005, 280, 41262–41269. [Google Scholar] [CrossRef]
- Dragovic, Z.; Broadley, S.A.; Shomura, Y.; Bracher, A.; Hartl, F.U. Molecular chaperones of the Hsp110 family act as nucleotide exchange factors of Hsp70s. EMBO J. 2006, 25, 2519–2528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raviol, H.; Sadlish, H.; Rodriguez, F.; Mayer, M.P.; Bukau, B. Chaperone network in the yeast cytosol: Hsp110 is revealed as an Hsp70 nucleotide exchange factor. EMBO J. 2006, 25, 2510–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.J.; Chen, X.; Subjeck, J.R. Hsp110 protects heat-denatured proteins and confers cellular thermoresistance. J. Biol. Chem. 1997, 272, 31636–31640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.J.; Easton, D.; Murawski, M.; Kaneko, Y.; Subjeck, J.R. The chaperoning activity of Hsp110. Identification of functional domains by use of targeted deletions. J. Biol. Chem. 1999, 274, 15712–15718. [Google Scholar] [CrossRef] [Green Version]
- Schmidtke, G.; Schmidt, M.; Kloetzel, P.M. Maturation of mammalian 20 S proteasome: Purification and characterization of 13 S and 16 S proteasome precursor complexes. J. Mol. Biol. 1997, 268, 95–106. [Google Scholar] [CrossRef]
- Hammack, L.J.; Firestone, K.; Chang, W.; Kusmierczyk, A.R. Molecular chaperones of the Hsp70 family assist in the assembly of 20S proteasomes. Biochem. Biophys. Res. Commun. 2017, 486, 438–443. [Google Scholar] [CrossRef] [Green Version]
- Hammack, L.J.; Kusmierczyk, A.R. Assembly of proteasome subunits into non-canonical complexes in vivo. Biochem. Biophys. Res. Commun. 2017, 482, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Shaner, L.; Trott, A.; Goeckeler, J.L.; Brodsky, J.L.; Morano, K.A. The function of the yeast molecular chaperone Sse1 is mechanistically distinct from the closely related hsp70 family. J. Biol. Chem. 2004, 279, 21992–22001. [Google Scholar] [CrossRef] [Green Version]
- Werner-Washburne, M.; Stone, D.E.; Craig, E.A. Complex interactions among members of an essential subfamily of hsp70 genes in Saccharomyces cerevisiae. Mol. Cell Biol. 1987, 7, 2568–2577. [Google Scholar] [CrossRef]
- Shaner, L.; Sousa, R.; Morano, K.A. Characterization of Hsp70 binding and nucleotide exchange by the yeast Hsp110 chaperone Sse1. Biochemistry 2006, 45, 15075–15084. [Google Scholar] [CrossRef]
- Schuermann, J.P.; Jiang, J.; Cuellar, J.; Llorca, O.; Wang, L.; Gimenez, L.E.; Jin, S.; Taylor, A.B.; Demeler, B.; Morano, K.A.; et al. Structure of the Hsp110:Hsc70 nucleotide exchange machine. Mol. Cell 2008, 31, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Makhnevych, T.; Houry, W.A. The control of spindle length by Hsp70 and Hsp110 molecular chaperones. FEBS Lett. 2013, 587, 1067–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, K.; Saeki, Y.; Yashiroda, H.; Yagi, H.; Kaiho, A.; Murata, S.; Yamane, T.; Tanaka, K.; Mizushima, T.; Kato, K. Pba3-Pba4 heterodimer acts as a molecular matchmaker in proteasome alpha-ring formation. Biochem. Biophys. Res. Commun. 2014, 450, 1110–1114. [Google Scholar] [CrossRef]
- Yao, Y.; Toth, C.R.; Huang, L.; Wong, M.L.; Dias, P.; Burlingame, A.L.; Coffino, P.; Wang, C.C. alpha5 subunit in Trypanosoma brucei proteasome can self-assemble to form a cylinder of four stacked heptamer rings. Biochem. J. 1999, 344, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Gerards, W.L.; Enzlin, J.; Häner, M.; Hendriks, I.L.; Aebi, U.; Bloemendal, H.; Boelens, W. The human alpha-type proteasomal subunit HsC8 forms a double ring like structure, but does not assemble into proteasome-like particles with the beta-type subunits HsDelta or HsBPROS26. J. Biol. Chem. 1997, 272, 10080–10086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattoo, R.U.H.; Sharma, S.K.; Priya, S.; Finka, A.; Goloubinoff, P. Hsp110 is a Bona Fide chaperone using ATP to unfold stable misfolded polypeptides and reciprocally collaborate with Hsp70 to solubilize protein aggregates. J. Biol. Chem. 2013, 288, 21399–21411. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias, A.C.; Matos, J.; Dohmen, R.J.; Ramos, P.C. Hsp70 and Hsp110 Chaperones Promote Early Steps of Proteasome Assembly. Biomolecules 2023, 13, 11. https://doi.org/10.3390/biom13010011
Matias AC, Matos J, Dohmen RJ, Ramos PC. Hsp70 and Hsp110 Chaperones Promote Early Steps of Proteasome Assembly. Biomolecules. 2023; 13(1):11. https://doi.org/10.3390/biom13010011
Chicago/Turabian StyleMatias, Ana C., Joao Matos, R. Jürgen Dohmen, and Paula C. Ramos. 2023. "Hsp70 and Hsp110 Chaperones Promote Early Steps of Proteasome Assembly" Biomolecules 13, no. 1: 11. https://doi.org/10.3390/biom13010011
APA StyleMatias, A. C., Matos, J., Dohmen, R. J., & Ramos, P. C. (2023). Hsp70 and Hsp110 Chaperones Promote Early Steps of Proteasome Assembly. Biomolecules, 13(1), 11. https://doi.org/10.3390/biom13010011