
Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders

{kind=link}
{kind=link}
Abstract
:1. Introduction
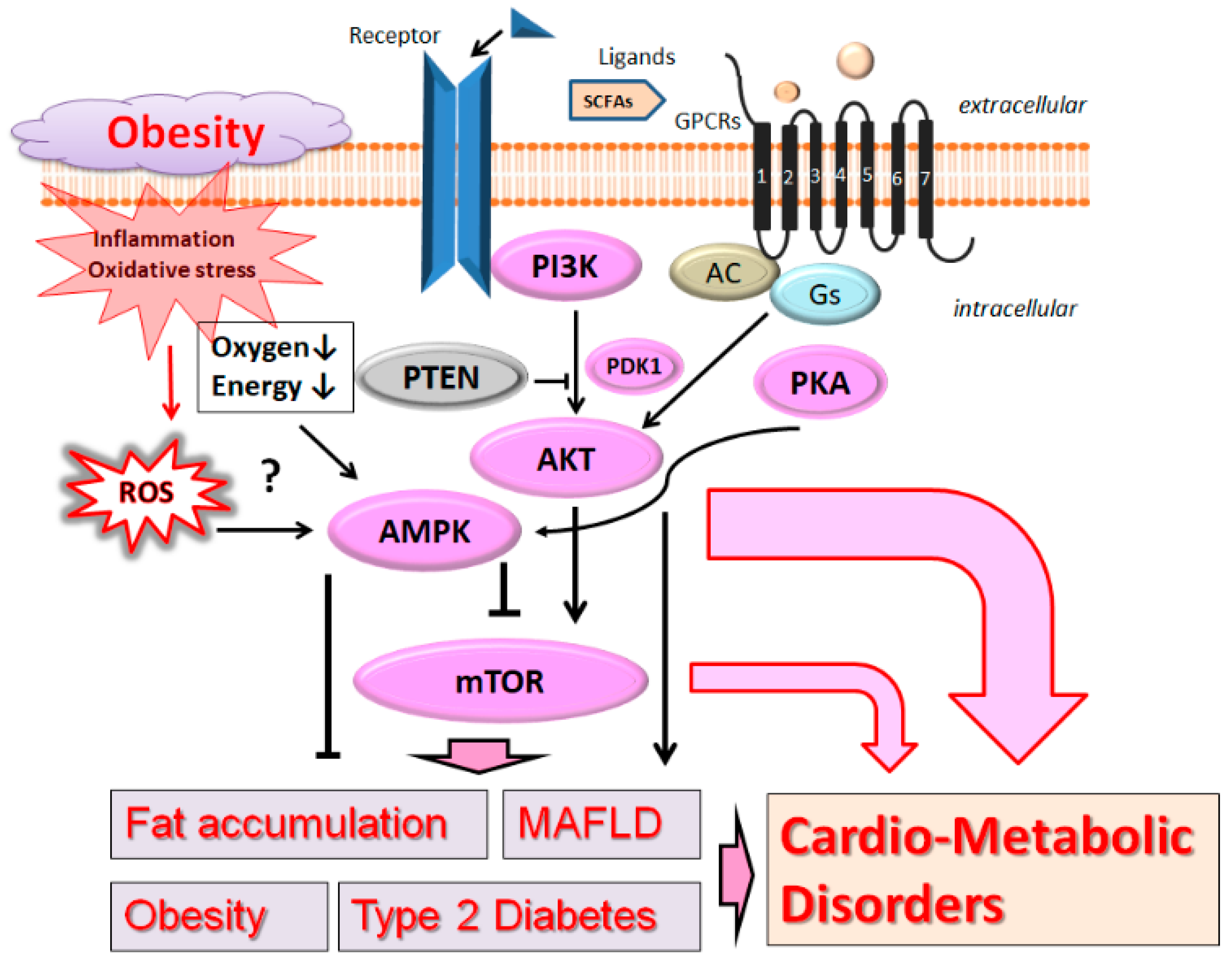
2. PI3K/AKT and AMPK Pathway Involved in Obesity, MAFLD, and Diabetes
3. Probiotics for the Modulation of PI3K/AKT and AMPK Pathway
4. Possible Mechanism behind the Beneficial Effects of Probiotics on Obesity and Diabetes
5. Genetically Modified Probiotics with Lactococci lactis
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACE2 | Angiotensin-converting enzyme 2 |
| AMP | Adenosine monophosphate |
| A. muciniphila | Akkermansia muciniphila |
| AMPK | Activated protein kinase |
| B. lactis | Bifidobacterium animalis subsp. lactis |
| CRISPR | Clustered regularly interspaced palindromic repeat |
| Cas | CRISPR-associated proteins |
| Cas9 | CRISPR-associated proteins 9 |
| DSS | Dextran sodium sulfate |
| GAD | Glutamic acid decarboxylase |
| GLP-1 | Glucagon-like protein-1 |
| HDAC | Histone deacetylase |
| HOMA-IR | Homeostatic model assessment for insulin resistance |
| IBD | Inflammatory bowel disease |
| IL-10 | Interleukin-10 |
| IL-35 | Interleukin-35 |
| LDL | Low density lipoprotein |
| L.lactis | Lactococcus lactis |
| L. plantarum | Lactiplantibacillus plantarum |
| L. rhamnosus | Lactobacillus rhamnosus |
| NAFLD | Non-alcoholic fatty liver disease |
| MAFLD | Metabolic-associated fatty liver disease |
| NASH | Non-alcoholic steatohepatitis |
| mTOR | Mammalian/mechanistic target of rapamycin |
| PI3K | Phosphoinositide-3 kinase |
| PPAR | Peroxisome proliferator-activated receptor |
| QOL | Quality of life |
| ROS | Reactive oxygen species |
| SIRT1 | Sirtuin1 |
| T2D | Type 2 diabetes |
References
- Crisci, G.; De Luca, M.; D’Assante, R.; Ranieri, B.; D’Agostino, A.; Valente, V.; Giardino, F.; Capone, V.; Chianese, S.; Rega, S.; et al. Effects of Exercise on Heart Failure with Preserved Ejection Fraction: An Updated Review of Literature. J. Cardiovasc. Dev. Dis. 2022, 9, 241. [Google Scholar] [CrossRef]
- Schroeder, K.; Kubik, M.Y.; Sirard, J.R.; Lee, J.; Fulkerson, J.A. Sleep is Inversely Associated with Sedentary Time among Youth with Obesity. Am. J. Health Behav. 2020, 44, 756–764. [Google Scholar] [CrossRef]
- Gutiérrez-Cuevas, J.; Santos, A.; Armendariz-Borunda, J. Pathophysiological Molecular Mechanisms of Obesity: A Link between MAFLD and NASH with Cardiovascular Diseases. Int. J. Mol. Sci. 2021, 22, 11629. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; International, Consensus, Panel. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef]
- Juul, F.; Deierlein, A.L.; Vaidean, G.; Quatromoni, P.A.; Parekh, N. Ultra-processed Foods and Cardiometabolic Health Outcomes: From Evidence to Practice. Curr. Atheroscler Rep. 2022, 24, 849–860. [Google Scholar] [CrossRef]
- Vahedi, H.; Atefi, M.; Entezari, M.H.; Hassanzadeh, A. The effect of sesame oil consumption compared to sunflower oil on lipid profile, blood pressure, and anthropometric indices in women with non-alcoholic fatty liver disease: A randomized double-blind controlled trial. Trials 2022, 23, 551. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, L.; Jiang, W.; Du, T.; Yuan, G. Non-obese NAFLD had no better cardio-metabolic risk profile than obese NAFLD in type 2 diabetic patients. Cardiovasc. Diabetol. 2022, 21, 210. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes. 2018, 9, 308–325. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Baptista, L.C.; Roberts, L.M.; Jumbo-Lucioni, P.; McMahon, L.L.; Buford, T.W.; Carter, C.S. The Gut Microbiome as a Therapeutic Target for Cognitive Impairment. J. Gerontol. A. Biol. Sci. Med. Sci. 2020, 75, 1242–1250. [Google Scholar] [CrossRef]
- Bonsembiante, L.; Targher, G.; Maffeis, C. Non-alcoholic fatty liver disease in obese children and adolescents: A role for nutrition? Eur. J. Clin. Nutr. 2022, 76, 28–39. [Google Scholar] [CrossRef]
- Mutalub, Y.B.; Abdulwahab, M.; Mohammed, A.; Yahkub, A.M.; Al-Mhanna, S.B.; Yusof, W.; Tang, S.P.; Rasool, A.H.G.; Mokhtar, S.S. Gut Microbiota Modulation as a Novel Therapeutic Strategy in Cardiometabolic Diseases. Foods 2022, 11, 2575. [Google Scholar] [CrossRef]
- Frolova, M.S.; Suvorova, I.A.; Iablokov, S.N.; Petrov, S.N.; Rodionov, D.A. Genomic reconstruction of short-chain fatty acid production by the human gut microbiota. Front. Mol. Biosci. 2022, 9, 949563. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Deng, M.; Lu, A.; Chen, Y.; Chen, Y.; Wu, C.; Tan, Z.; Boini, K.M.; Yang, T.; Zhu, Q.; et al. Sodium butyrate attenuates angiotensin II-induced cardiac hypertrophy by inhibiting COX2/PGE2 pathway via a HDAC5/HDAC6-dependent mechanism. J. Cell. Mol. Med. 2019, 23, 8139–8150. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Yoo, S.A.; Kim, W.U. Gut microbiota in autoimmunity: Potential for clinical applications. Arch. Pharm. Res. 2016, 39, 1565–1576. [Google Scholar] [CrossRef]
- Motta, J.P.; Magne, L.; Descamps, D.; Rolland, C.; Squarzoni-Dale, C.; Rousset, P.; Martin, L.; Cenac, N.; Balloy, V.; Huerre, M.; et al. Modifying the protease, antiprotease pattern by elafin overexpression protects mice from colitis. Gastroenterology 2011, 140, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Houghton, P.J. Targeting mTOR signaling for cancer therapy. Curr. Opin. Pharmacol. 2003, 3, 371–377. [Google Scholar] [CrossRef]
- Janku, F.; Yap, T.A.; Meric-Bernstam, F. Targeting the PI3K pathway in cancer: Are we making headway? Nat. Rev. Clin. Oncol. 2018, 15, 273–291. [Google Scholar] [CrossRef]
- Wu, C.; Jiang, F.; Wei, K.; Jiang, Z. Exercise activates the PI3K-AKT signal pathway by decreasing the expression of 5α-reductase type 1 in PCOS rats. Sci. Rep. 2018, 8, 7982. [Google Scholar] [CrossRef]
- Kitagishi, Y.; Nakanishi, A.; Ogura, Y.; Matsuda, S. Dietary regulation of PI3K/AKT/GSK-3β pathway in Alzheimer’s disease. Alzheimers Res. Ther. 2014, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Guan, P.; Sun, Z.M.; Wang, N.; Zhou, J.; Luo, L.F.; Zhao, Y.S.; Ji, E.S. Resveratrol prevents chronic intermittent hypoxia-induced cardiac hypertrophy by targeting the PI3K/AKT/mTOR pathway. Life Sci. 2019, 233, 116748. [Google Scholar] [CrossRef]
- Liu, M.H.; Yuan, C.; He, J.; Tan, T.P.; Wu, S.J.; Fu, H.Y.; Liu, J.; Yu, S.; Chen, Y.D.; Le, Q.F.; et al. Resveratrol protects PC12 cells from high glucose-induced neurotoxicity via PI3K/Akt/FoxO3a pathway. Cell Mol. Neurobiol. 2015, 35, 513–522. [Google Scholar] [CrossRef]
- Han, Z.; Zhao, D.; Han, M.; Zhang, R.; Hao, Y. Knockdown of miR-372-3p Inhibits the Development of Diabetic Cardiomyopathy by Accelerating Angiogenesis via Activating the PI3K/AKT/mTOR/HIF-1α Signaling Pathway and Suppressing Oxidative Stress. Oxid. Med. Cell Longev. 2022, 2022, 4342755. [Google Scholar] [CrossRef]
- Sangweni, N.F.; Mosa, R.A.; Dludla, P.V.; Kappo, A.P.; Opoku, A.R.; Muller, C.J.F.; Johnson, R. The triterpene, methyl-3β-hydroxylanosta-9,24-dien-21-oate (RA3), attenuates high glucose-induced oxidative damage and apoptosis by improving energy metabolism. Phytomedicine 2021, 85, 153546. [Google Scholar] [CrossRef]
- Wang, S.; Duan, J.; Liao, J.; Wang, Y.; Xiao, X.; Li, L.; Liu, Y.; Gu, H.; Yang, P.; Fu, D.; et al. LncRNA H19 inhibits ER stress induced apoptosis and improves diabetic cardiomyopathy by regulating PI3K/AKT/mTOR axis. Aging 2022, 14, 6809–6828. [Google Scholar] [CrossRef]
- Yang, X.; Yao, L.; Yuan, M.; Zhang, X.; Jakubowska, M.A.; Ferdek, P.E.; Dai, L.; Yang, J.; Jin, T.; Deng, L.; et al. Transcriptomics and Network Pharmacology Reveal the Protective Effect of Chaiqin Chengqi Decoction on Obesity-Related Alcohol-Induced Acute Pancreatitis via Oxidative Stress and PI3K/Akt Signaling Pathway. Front. Pharmacol. 2022, 13, 896523. [Google Scholar] [CrossRef]
- Tamana, S.K.; Tun, H.M.; Konya, T.; Chari, R.S.; Field, C.J.; Guttman, D.S.; Becker, A.B.; Moraes, T.J.; Turvey, S.E.; Subbarao, P.; et al. Bacteroides-dominant gut microbiome of late infancy is associated with enhanced neurodevelopment. Gut Microbes. 2021, 13, 1–17. [Google Scholar] [CrossRef]
- Gong, Y.; Guo, Y.; Jiang, Y.; Xing, Z.; Zhang, H.; Wang, H.; Gong, Y. Intracerebroventricular injection of ghrelin receptor antagonist alleviated NAFLD via improving hypothalamic insulin resistance. Iran J. Basic Med. Sci. 2022, 25, 1117–1122. [Google Scholar]
- Zhang, C.Y.; Tan, X.H.; Yang, H.H.; Jin, L.; Hong, J.R.; Zhou, Y.; Huang, X.T. COX-2/sEH Dual Inhibitor Alleviates Hepatocyte Senescence in NAFLD Mice by Restoring Autophagy through Sirt1/PI3K/AKT/mTOR. Int. J. Mol. Sci. 2022, 23, 8267. [Google Scholar] [CrossRef]
- Feng, S.Y.; Wu, S.J.; Chang, Y.C.; Ng, L.T.; Chang, S.J. Stimulation of GLUT4 Glucose Uptake by Anthocyanin-Rich Extract from Black Rice (Oryza sativa L.) via PI3K/Akt and AMPK/p38 MAPK Signaling in C2C12 Cells. Metabolites 2022, 12, 856. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Anand, S.K.; Singh, N.; Dwarkanath, A.; Dwivedi, U.N.; Kakkar, P. Berbamine induced activation of the SIRT1/LKB1/AMPK signaling axis attenuates the development of hepatic steatosis in high-fat diet-induced NAFLD rats. Food Funct. 2021, 12, 892–909. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Q.; Yang, S.; Zheng, T.; Shao, J.; Guan, W.; Chen, F.; Zhang, S. Energy deprivation-induced AMPK activation inhibits milk synthesis by targeting PrlR and PGC-1α. Cell Commun. Signal. 2022, 20, 25. [Google Scholar] [CrossRef]
- Berthou, F.; Sobolewski, C.; Abegg, D.; Fournier, M.; Maeder, C.; Dolicka, D.; de Sousa, M.C.; Adibekian, A.; Foti, M. Hepatic PTEN Signaling Regulates Systemic Metabolic Homeostasis through Hepatokines-Mediated Liver-to-Peripheral Organs Crosstalk. Int. J. Mol. Sci. 2022, 23, 3959. [Google Scholar] [CrossRef]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef]
- Zhang, J.; Pivovarova-Ramich, O.; Kabisch, S.; Markova, M.; Hornemann, S.; Sucher, S.; Rohn, S.; Machann, J.; Pfeiffer, A.F.H. High Protein Diets Improve Liver Fat and Insulin Sensitivity by Prandial but Not Fasting Glucagon Secretion in Type 2 Diabetes. Front. Nutr. 2022, 9, 808346. [Google Scholar] [CrossRef]
- Sinha-Hikim, I.; Sinha-Hikim, A.P.; Shen, R.; Kim, H.J.; French, S.W.; Vaziri, N.D.; Crum, A.C.; Rajavashisth, T.B.; Norris, K.C. A novel cystine based antioxidant attenuates oxidative stress and hepatic steatosis in diet-induced obese mice. Exp. Mol. Pathol. 2011, 91, 419–428. [Google Scholar] [CrossRef] [Green Version]
- El Sayed, N.S.; Kandil, E.A.; Ghoneum, M.H. Enhancement of Insulin/PI3K/Akt Signaling Pathway and Modulation of Gut Microbiome by Probiotics Fermentation Technology, a Kefir Grain Product, in Sporadic Alzheimer’s Disease Model in Mice. Front. Pharmacol. 2021, 12, 666502. [Google Scholar] [CrossRef]
- Xia, J.; Lv, L.; Liu, B.; Wang, S.; Zhang, S.; Wu, Z.; Yang, L.; Bian, X.; Wang, Q.; Wang, K.; et al. Akkermansia muciniphila Ameliorates Acetaminophen-Induced Liver Injury by Regulating Gut Microbial Composition and Metabolism. Microbiol. Spectr. 2022, 10, e0159621. [Google Scholar] [CrossRef] [PubMed]
- Won, G.; Choi, S.I.; Kang, C.H.; Kim, G.H. Lactiplantibacillus plantarum MG4296 and Lacticaseibacillus paracasei MG5012 Ameliorates Insulin Resistance in Palmitic Acid-Induced HepG2 Cells and High Fat Diet-Induced Mice. Microorganisms 2021, 9, 1139. [Google Scholar] [CrossRef] [PubMed]
- Won, G.; Choi, S.I.; Park, N.; Kim, J.E.; Kang, C.H.; Kim, G.H. In Vitro Antidiabetic, Antioxidant Activity, and Probiotic Activities of Lactiplantibacillus plantarum and Lacticaseibacillus paracasei Strains. Curr. Microbiol. 2021, 78, 3181–3191. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Feng, S.; Yan, J.; Luan, D.; Sun, P.; Shao, P. Antidiabetic potential of polysaccharides from Brasenia schreberi regulating insulin signaling pathway and gut microbiota in type 2 diabetic mice. Curr. Res. Food Sci. 2022, 5, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Zhou, M.; Ju, Z.; Luo, J.; Jin, J.; Shen, L.; Zhou, P.; Huang, R. Potential role of gut microbiota-LCA-INSR axis in high fat-diet-induced non-alcoholic fatty liver dysfunction: From perspective of radiation variation. Curr. Res. Food Sci. 2022, 5, 1685–1700. [Google Scholar] [CrossRef]
- Xu, M.; Fu, L.; Zhang, J.; Wang, T.; Fan, J.; Zhu, B.; Dziugan, P.; Zhang, B.; Zhao, H. Potential of Inactivated Bifidobacterium Strain in Attenuating Benzo(A)Pyrene E xposure-Induced Damage in Colon Epithelial Cells In Vitro. Toxics 2020, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, S.; Fan, H.; Han, M.; Xie, J.; Du, J.; Peng, F. Bifidobacterium lactis combined with Lactobacillus plantarum inhibit glioma growth in mice through modulating PI3K/AKT pathway and gut microbiota. Front. Microbiol. 2022, 13, 986837. [Google Scholar] [CrossRef]
- Saikia, R.; Joseph, J. AMPK: A key regulator of energy stress and calcium-induced autophagy. J. Mol. Med. 2021, 99, 1539–1551. [Google Scholar] [CrossRef]
- Lin, J.Y.; Kuo, W.W.; Baskaran, R.; Kuo, C.H.; Chen, Y.A.; Chen, W.S.; Ho, T.J.; Day, C.H.; Mahalakshmi, B.; Huang, C.Y. Swimming exercise stimulates IGF1/PI3K/Akt and AMPK/SIRT1/PGC1α survival signaling to suppress apoptosis and inflammation in aging hippocampus. Aging 2020, 12, 6852–6864. [Google Scholar] [CrossRef]
- Zhou, S.; Xue, J.; Shan, J.; Hong, Y.; Zhu, W.; Nie, Z.; Zhang, Y.; Ji, N.; Luo, X.; Zhang, T.; et al. Gut-Flora-Dependent Metabolite Trimethylamine-N-Oxide Promotes Atherosclerosis-Associated Inflammation Responses by Indirect ROS Stimulation and Signaling Involving AMPK and SIRT1. Nutrients 2022, 14, 3338. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Li, Y.; Chen, M.; Xue, L.; Wang, J.; Ding, Y.; Zhang, J.; Wu, S.; Ye, Q.; Zhang, S.; et al. Antihypertensive Activity of Milk Fermented by Lactiplantibacillus plantarum SR37-3 and SR61-2 in L-NAME-Induced Hypertensive Rats. Foods 2022, 11, 2332. [Google Scholar] [CrossRef]
- Shi, M.; Yue, Y.; Ma, C.; Dong, L.; Chen, F. Pasteurized Akkermansia muciniphila Ameliorate the LPS-Induced Intestinal Barrier Dysfunction via Modulating AMPK and NF-κB through TLR2 in Caco-2 Cells. Nutrients 2022, 14, 764. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Nikpoor, N.; Tompkins, T.A.; Rho, J.M.; Scantlebury, M.H.; Shearer, J. Probiotics counteract hepatic steatosis caused by ketogenic diet and upregulate AMPK signaling in a model of infantile epilepsy. EBioMedicine. 2022, 76, 103838. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Jing, Y.; Zhou, X.; Zhang, N.; Tai, J.; Cao, Y. Bacillus licheniformis Zhengchangsheng® Inhibits Obesity by Regulating the AMP-Activated Protein Kinase Signaling Pathway. Probiotics Antimicrob. Proteins 2021, 13, 1658–1667. [Google Scholar] [CrossRef] [PubMed]
- Wa, Y.; Yin, B.; He, Y.; Xi, W.; Huang, Y.; Wang, C.; Guo, F.; Gu, R. Effects of Single Probiotic- and Combined Probiotic-Fermented Milk on Lipid Metabolism in Hyperlipidemic Rats. Front. Microbiol. 2019, 10, 1312. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Song, X.; An, Y.; Wu, X.; Zhang, W.; Li, J.; Sun, Y.; Jin, G.; Liu, X.; Guo, Z.; et al. Lactobacillus rhamnosus GG Colonization in Early Life Ameliorates Inflammaging of Offspring by Activating SIRT1/AMPK/PGC-1α Pathway. Oxid. Med. Cell Longev. 2021, 2021, 3328505. [Google Scholar] [CrossRef]
- Schrezenmeir, J.; de Vrese, M. Probiotics, prebiotics, and synbiotics--approaching a definition. Am. J. Clin. Nutr. 2001, 73, 361S–364S. [Google Scholar] [CrossRef] [Green Version]
- Endesfelder, D.; Zu Castell, W.; Ardissone, A.; Davis-Richardson, A.G.; Achenbach, P.; Hagen, M.; Pflueger, M.; Gano, K.A.; Fagen, J.R.; Drew, J.C.; et al. Compromised gut microbiota networks in children with anti-islet cell autoimmunity. Diabetes 2014, 63, 2006–2014. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Wade, P.A. Crosstalk between the microbiome and epigenome: Messages from bugs. J. Biochem. 2018, 163, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Wali, J.A.; Jarzebska, N.; Raubenheimer, D.; Simpson, S.J.; Rodionov, R.N.; O’Sullivan, J.F. Cardio-Metabolic Effects of High-Fat Diets and Their Underlying Mechanisms-A Narrative Review. Nutrients 2020, 12, 1505. [Google Scholar] [CrossRef]
- Krautkramer, K.A.; Kreznar, J.H.; Romano, K.A.; Vivas, E.I.; Barrett-Wilt, G.A.; Rabaglia, M.E.; Keller, M.P.; Attie, A.D.; Rey, F.E.; Denu, J.M. Diet-Microbiota Interactions Mediate Global Epigenetic Programming in Multiple Host Tissues. Mol. Cell. 2016, 64, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Bolduc, J.F.; Hany, L.; Barat, C.; Ouellet, M.; Tremblay, M.J. Epigenetic Metabolite Acetate Inhibits Class I/II Histone Deacetylases, Promotes Histone Acetylation, and Increases HIV-1 Integration in CD4+ T Cells. J. Virol. 2017, 91, e01943-16. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Chen, H.; Li, X.; Li, D.; Sun, Y.; Yang, L.; Ma, Y.; Chan, E.C.Y. Lactobacillus paracasei IMC 502 ameliorate type 2 diabetes by mediating gut microbiota-SCFAs-hormone/inflammation pathway in mice. J. Sci. Food Agric. 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Yang, M.; Zheng, J.; Zong, X.; Yang, X.; Zhang, Y.; Man, C.; Jiang, Y. Preventive Effect and Molecular Mechanism of Lactobacillus rhamnosus JL1 on Food-Borne Obesity in Mice. Nutrients 2021, 13, 3989. [Google Scholar] [CrossRef] [PubMed]
- Galfo, M.; Censi, L.; D’Addezio, L.; Melini, F.; Martone, D. Overweight/obesity and lifestyle factors among Italian adolescents: The ALIADO study. Minerva Pediatr 2022, 74, 251–258. [Google Scholar] [CrossRef]
- Choi, B.S.; Brunelle, L.; Pilon, G.; Cautela, B.G.; Tompkins, T.A.; Drapeau, V.; Marette, A.; Tremblay, A. Lacticaseibacillus rhamnosus HA-114 improves eating behaviors and mood-related factors in adults with overweight during weight loss: A randomized controlled trial. Nutr. Neurosci. 2022, 17, 1–13. [Google Scholar] [CrossRef]
- Zafar, H.; Ain, N.U.; Alshammari, A.; Alghamdi, S.; Raja, H.; Ali, A.; Siddique, A.; Tahir, S.D.; Akbar, S.; Arif, M.; et al. Lacticaseibacillus rhamnosus FM9 and Limosilactobacillus fermentum Y57 Are as Effective as Statins at Improving Blood Lipid Profile in High Cholesterol, High-Fat Diet Model in Male Wistar Rats. Nutrients 2022, 14, 1654. [Google Scholar] [CrossRef]
- Lee, C.S.; Park, M.H.; Kim, B.K.; Kim, S.H. Antiobesity Effect of Novel Probiotic Strains in a Mouse Model of High-Fat Diet-Induced Obesity. Probiotics Antimicrob. Proteins 2021, 13, 1054–1067. [Google Scholar] [CrossRef]
- Van Deuren, T.; Blaak, E.E.; Canfora, E.E. Butyrate to combat obesity and obesity-associated metabolic disorders: Current status and future implications for therapeutic use. Obes. Rev. 2022, 23, e13498. [Google Scholar] [CrossRef]
- Zhu, W.; Peng, K.; Zhao, Y.; Xu, C.; Tao, X.; Liu, Y.; Huang, Y.; Yang, X. Sodium butyrate attenuated diet-induced obesity, insulin resistance and inflammation partly by promoting fat thermogenesis via intro-adipose sympathetic innervation. Front. Pharmacol. 2022, 13, 938760. [Google Scholar] [CrossRef]
- Gart, E.; van Duyvenvoorde, W.; Toet, K.; Caspers, M.P.M.; Verschuren, L.; Nielsen, M.J.; Leeming, D.J.; Souto Lima, E.; Menke, A.; Hanemaaijer, R.; et al. Butyrate Protects against Diet-Induced NASH and Liver Fibrosis and Suppresses Specific Non-Canonical TGF-β Signaling Pathways in Human Hepatic Stellate Cells. Biomedicines 2021, 9, 1954. [Google Scholar] [CrossRef]
- Khan, S.; Jena, G.B. Protective role of sodium butyrate, a HDAC inhibitor on beta-cell proliferation, function and glucose homeostasis through modulation of p38/ERK MAPK and apoptotic pathways: Study in juvenile diabetic rat. Chem. Biol. Interact. 2014, 213, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Zhang, J.; Wu, Q.; Chen, J.; Liu, J.; Wang, L.; Chen, C.; Xu, J.; Zhang, H.; Shi, C.; et al. The role of microbiota in the development of colorectal cancer. Int. J. Cancer. 2019, 145, 2032–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondičová, M.; Irwin, R.E.; Thursby, S.J.; Hilman, L.; Caffrey, A.; Cassidy, T.; McLaughlin, M.; Lees-Murdock, D.J.; Ward, M.; Murphy, M.; et al. Folic acid intervention during pregnancy alters DNA methylation, affecting neural target genes through two distinct mechanisms. Clin. Epigenetics. 2022, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-Humarán, L.G.; Aubry, C.; Motta, J.P.; Deraison, C.; Steidler, L.; Vergnolle, N.; Chatel, J.M.; Langella, P. Engineering lactococci and lactobacilli for human health. Curr. Opin. Microbiol. 2013, 16, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Pontes, D.S.; de Azevedo, M.S.; Chatel, J.M.; Langella, P.; Azevedo, V.; Miyoshi, A. Lactococcus lactis as a live vector: Heterologous protein production and DNA delivery systems. Protein Expr. Purif. 2011, 79, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Siegers, K.; Entian, K.D. Genes involved in immunity to the lantibiotic nisin produced by Lactococcus lactis 6F3. Appl. Environ. Microbiol. 1995, 61, 1082–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levit, R.; Cortes-Perez, N.G.; de Moreno de Leblanc, A.; Loiseau, J.; Aucouturier, A.; Langella, P.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Use of genetically modified lactic acid bacteria and bifidobacteria as live delivery vectors for human and animal health. Gut Microbes. 2022, 14, 2110821. [Google Scholar] [CrossRef]
- Steidler, L.; Hans, W.; Schotte, L.; Neirynck, S.; Obermeier, F.; Falk, W.; Fiers, W.; Remaut, E. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science 2000, 289, 1352–1355. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tian, M.; Li, W.; Hao, F. Preventative delivery of IL-35 by Lactococcus lactis ameliorates DSS-induced colitis in mice. Appl. Microbiol. Biotechnol. 2019, 103, 7931–7941. [Google Scholar] [CrossRef]
- Maddaloni, M.; Kochetkova, I.; Hoffman, C.; Pascual, D.W. Delivery of IL-35 by Lactococcus lactis Ameliorates Collagen-Induced Arthritis in Mice. Front. Immunol. 2018, 9, 2691. [Google Scholar] [CrossRef] [Green Version]
- Robert, S.; Gysemans, C.; Takiishi, T.; Korf, H.; Spagnuolo, I.; Sebastiani, G.; Van Huynegem, K.; Steidler, L.; Caluwaerts, S.; Demetter, P.; et al. Oral delivery of glutamic acid decarboxylase (GAD)-65 and IL10 by Lactococcus lactis reverses diabetes in recent-onset NOD mice. Diabetes 2014, 63, 2876–2887. [Google Scholar] [CrossRef]
- Gomes-Santos, A.C.; de Oliveira, R.P.; Moreira, T.G.; Castro-Junior, A.B.; Horta, B.C.; Lemos, L.; de Almeida, L.A.; Rezende, R.M.; Cara, D.C.; Oliveira, S.C.; et al. Hsp65-Producing Lactococcus lactis Prevents Inflammatory Intestinal Disease in Mice by IL-10- and TLR2-Dependent Pathways. Front. Immunol. 2017, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Liu, J.; Hou, J.; Dong, Y.; Lu, Y.; Jin, L.; Cao, R.; Li, T.; Wu, J. Oral administration of recombinant Lactococcus lactis expressing HSP65 and tandemly repeated P277 reduces the incidence of type I diabetes in non-obese diabetic mice. PLoS ONE 2014, 9, e105701. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Yu, R.; Zuo, F.; Zhang, B.; Ma, H.; Chen, S. Recombinant Lactococcus lactis expressing bioactive exendin-4 to promote insulin secretion and beta-cell proliferation in vitro. Appl. Microbiol. Biotechnol. 2017, 101, 7177–7186. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.F.; Chiang, C.H.; Lin, S.J.; Song, P.P.; Liu, H.C.; Wu, T.J.; Lin, W.W. Recombinant Lactococcus lactis Expressing Ling Zhi 8 Protein Ameliorates Nonalcoholic Fatty Liver and Early Atherogenesis in Cholesterol-Fed Rabbits. Biomed. Res. Int. 2020, 2020, 3495682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.L.; Liang, Y.C.; Tseng, Y.S.; Huang, H.Y.; Chou, S.Y.; Hseu, R.S.; Huang, C.T.; Chiang, B.L. An immunomodulatory protein, Ling Zhi-8, induced activation and maturation of human monocyte-derived dendritic cells by the NF-kappaB and MAPK pathways. J. Leukoc. Biol. 2009, 86, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.J.; Chang, Y.S.; Lin, L.H.; Haung, C.F.; Wu, C.Y.; Ou, K.L. An Immunomodulatory Protein (Ling Zhi-8) from a Ganoderma lucidum Induced Acceleration of Wound Healing in Rat Liver Tissues after Monopolar Electrosurgery. Evid. Based Complement Alternat. Med. 2014, 2014, 916531. [Google Scholar] [CrossRef] [Green Version]
- Watterlot, L.; Rochat, T.; Sokol, H.; Cherbuy, C.; Bouloufa, I.; Lefèvre, F.; Gratadoux, J.J.; Honvo-Hueto, E.; Chilmonczyk, S.; Blugeon, S.; et al. Intragastric administration of a superoxide dismutase-producing recombinant Lactobacillus casei BL23 strain attenuates DSS colitis in mice. Int. J. Food Microbiol. 2010, 144, 35–41. [Google Scholar] [CrossRef]
- Verma, A.; Xu, K.; Du, T.; Zhu, P.; Liang, Z.; Liao, S.; Zhang, J.; Raizada, M.K.; Grant, M.B.; Li, Q. Expression of Human ACE2 in Lactobacillus and Beneficial Effects in Diabetic Retinopathy in Mice. Mol. Ther. Methods Clin. Dev. 2019, 14, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Floyd, J.L.; Dupont, M.; Harbour, A.; Adu Aggyeiwaah, Y.; Asare-Bediako, B.; Chakraborty, D.; Kichler, K.; Rohella, A.; Li Calzi, S.; et al. Maintenance of Enteral ACE2 Prevents Diabetic Retinopathy in Type 1 Diabetes. Circ. Res. 2022. [Google Scholar] [CrossRef]
- Greenhill, C. IBD: Elafin--a potential IBD therapy. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 686. [Google Scholar] [CrossRef]
- Bermúdez-Humarán, L.G.; Motta, J.P.; Aubry, C.; Kharrat, P.; Rous-Martin, L.; Sallenave, J.M.; Deraison, C.; Vergnolle, N.; Langella, P. Serine protease inhibitors protect better than IL-10 and TGF-β anti-inflammatory cytokines against mouse colitis when delivered by recombinant lactococci. Microb. Cell Fact. 2015, 14, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, S.; Van Huynegem, K.; Gysemans, C.; Mathieu, C.; Rottiers, P.; Steidler, L. Trimming of two major type 1 diabetes driving antigens, GAD65 and IA-2, allows for successful expression in Lactococcus lactis. Benef. Microbes. 2015, 6, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Cao, G.; Luo, C.; Tan, D.; Vong, C.T.; Xu, Y.; Wang, S.; Lu, H.; Wang, Y.; Jing, W. Emerging pharmacotherapy for inflammatory bowel diseases. Pharmacol. Res. 2022, 178, 106146. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Fang, D.; Lv, Q.; Wang, Z.; Liu, Y. Targeted Therapeutic Strategies in the Battle Against Pathogenic Bacteria. Front. Pharmacol. 2021, 12, 673239. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Xin, Y.; Zhang, Y.; Gu, X.; Kong, J. A rapid and versatile tool for genomic engineering in Lactococcus lactis. Microb. Cell Fact. 2019, 18, 22. [Google Scholar] [CrossRef] [Green Version]
- van der Els, S.; James, J.K.; Kleerebezem, M.; Bron, P.A. Versatile Cas9-Driven Subpopulation Selection Toolbox for Lactococcus lactis. Appl. Environ. Microbiol. 2018, 84, e02752-17. [Google Scholar] [CrossRef]
- Van Pijkeren, J.P.; Britton, R.A. High efficiency recombineering in lactic acid bacteria. Nucleic. Acids. Res. 2012, 40, e76. [Google Scholar] [CrossRef]
- Singh, B.; Mal, G.; Marotta, F. Designer Probiotics: Paving the Way to Living Therapeutics. Trends Biotechnol. 2017, 35, 679–682. [Google Scholar] [CrossRef]
- Oh, J.H.; van Pijkeren, J.P. CRISPR-Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic. Acids. Res. 2014, 42, e131. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; O’Flaherty, S.; Barrangou, R. CRISPR-based engineering of next-generation lactic acid bacteria. Curr. Opin. Microbiol. 2017, 37, 79–87. [Google Scholar] [CrossRef]
- Leenay, R.T.; Vento, J.M.; Shah, M.; Martino, M.E.; Leulier, F.; Beisel, C.L. Genome Editing with CRISPR-Cas9 in Lactobacillus plantarum Revealed That Editing Outcomes Can Vary Across Strains and Between Methods. Biotechnol. J. 2019, 14, e1700583. [Google Scholar] [CrossRef] [PubMed]
- Altenbuchner, J. Editing of the Bacillus subtilis Genome by the CRISPR-Cas9 System. Appl. Environ. Microbiol. 2016, 82, 5421–5427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Liu, R.; Garst, A.D.; Lee, T.; Nogué, V.S.I.; Beckham, G.T.; Gill, R.T. CRISPR EnAbled Trackable genome Engineering for isopropanol production in Escherichia coli. Metab. Eng. 2017, 41, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pursey, E.; Sünderhauf, D.; Gaze, W.H.; Westra, E.R.; van Houte, S. CRISPR-Cas antimicrobials: Challenges and future prospects. PLoS Pathog. 2018, 14, e1006990. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asai, T.; Yoshikawa, S.; Ikeda, Y.; Taniguchi, K.; Sawamura, H.; Tsuji, A.; Matsuda, S. Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders. Biomolecules 2023, 13, 10. https://doi.org/10.3390/biom13010010
Asai T, Yoshikawa S, Ikeda Y, Taniguchi K, Sawamura H, Tsuji A, Matsuda S. Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders. Biomolecules. 2023; 13(1):10. https://doi.org/10.3390/biom13010010
Chicago/Turabian StyleAsai, Tomoko, Sayuri Yoshikawa, Yuka Ikeda, Kurumi Taniguchi, Haruka Sawamura, Ai Tsuji, and Satoru Matsuda. 2023. "Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders" Biomolecules 13, no. 1: 10. https://doi.org/10.3390/biom13010010
APA StyleAsai, T., Yoshikawa, S., Ikeda, Y., Taniguchi, K., Sawamura, H., Tsuji, A., & Matsuda, S. (2023). Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders. Biomolecules, 13(1), 10. https://doi.org/10.3390/biom13010010