
Oral Immunization with Escherichia coli Nissle 1917 Expressing SARS-CoV-2 Spike Protein Induces Mucosal and Systemic Antibody Responses in Mice
 , , , ,
, , , ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of E. coli Nissle Expressing SARS-CoV-2 Spike Protein on Cell Membrane as Immunogen
2.2. Immunofluorescent Analysis of Spike Protein Expression on the Bacterial Surface
2.3. Western Blot Analysis of Spike Protein Expression
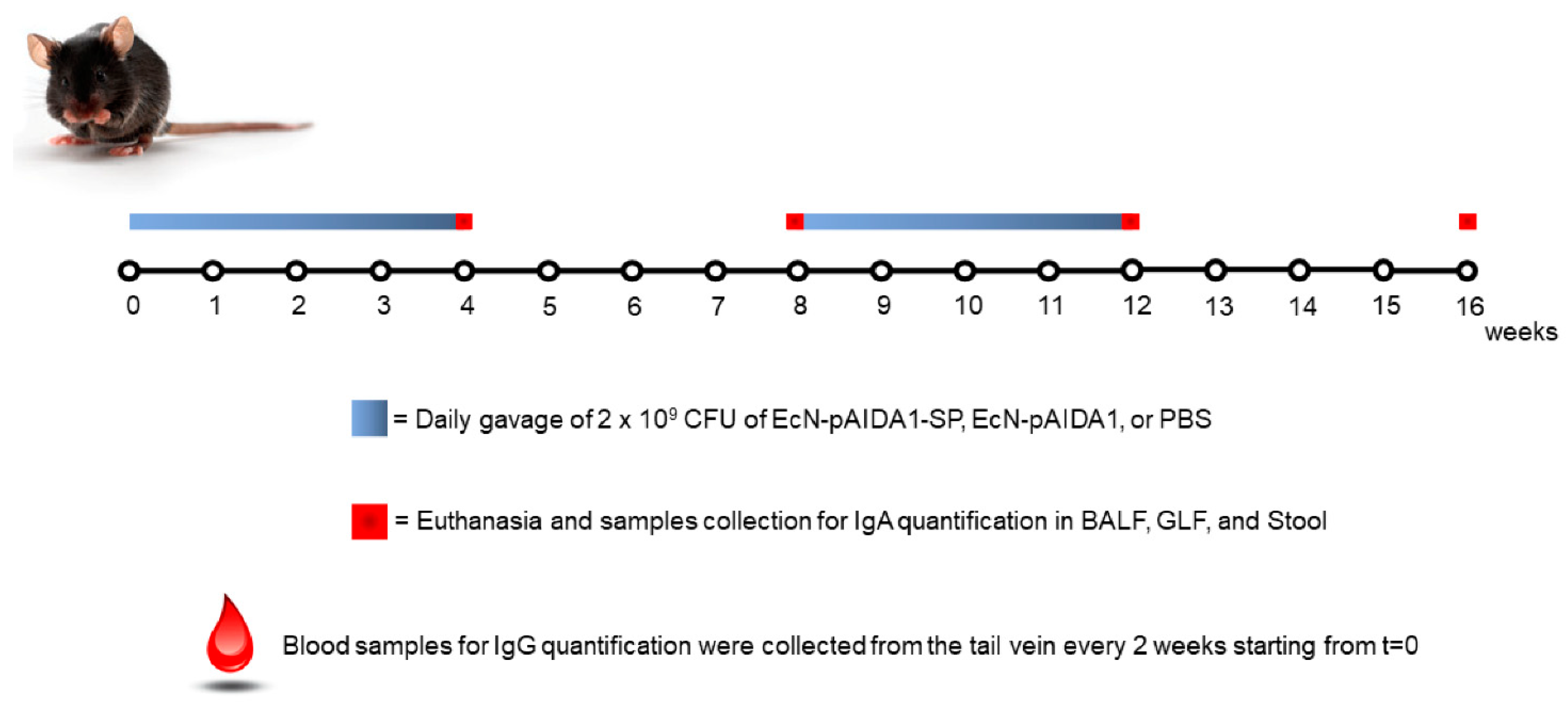
2.4. Animals, Immunization Protocol, and Sample Collection
2.5. Assessment of Safety Profile and Adverse Effects
2.6. ELISA for Specific Anti-SARS-CoV-2 S Antibodies Detection
2.7. Relative Avidity Index for Anti-SARS-CoV-2 SP IgA and IgG
2.8. Immunofluorescence Analysis
2.9. Histopathological Analysis
2.10. Enzyme-Linked Immunosorbent Assay for LPS
2.11. Statistical Analysis
3. Results
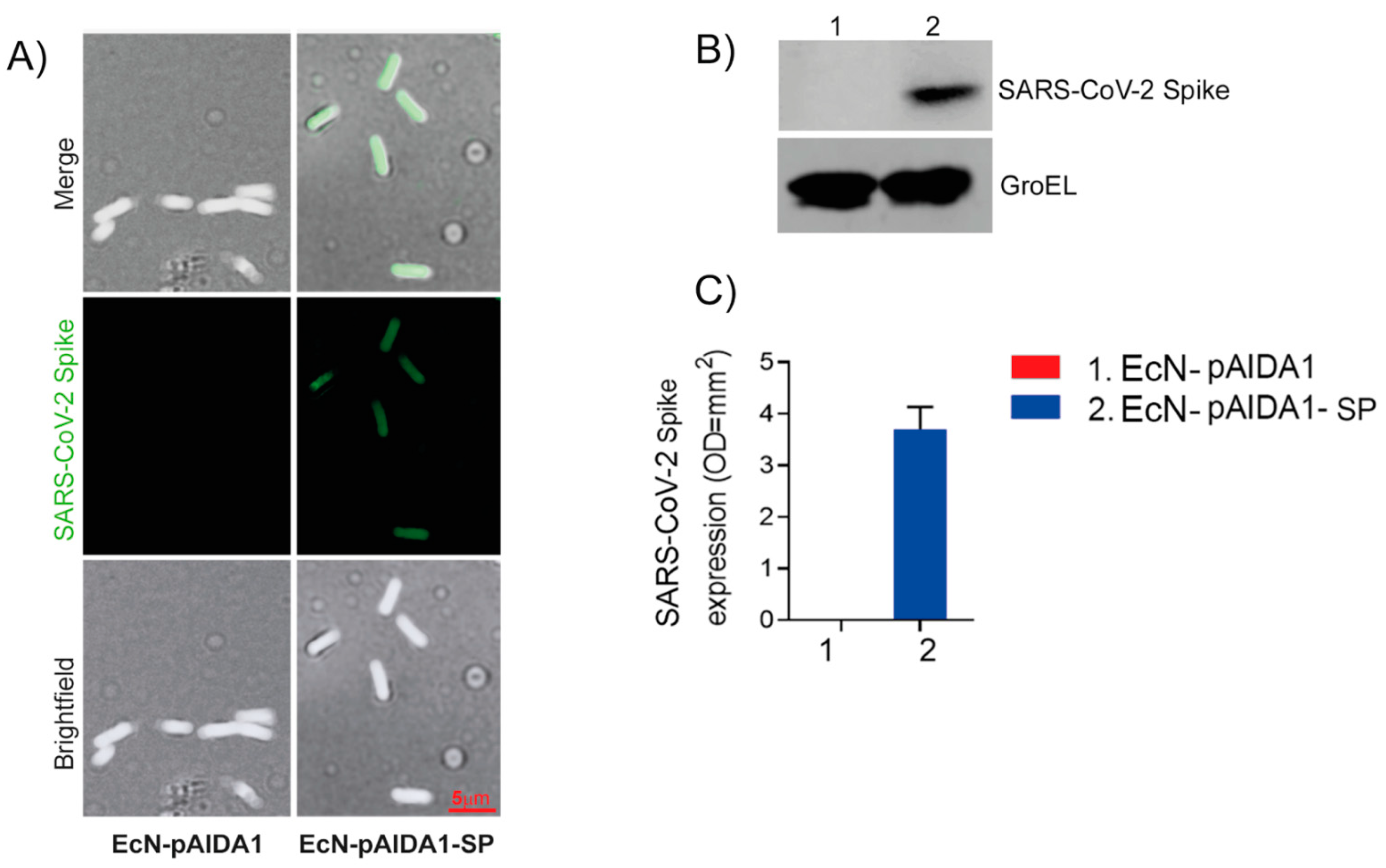
3.1. Expression of SARS-CoV-2 Spike Protein by Engineered E. coli Nissle
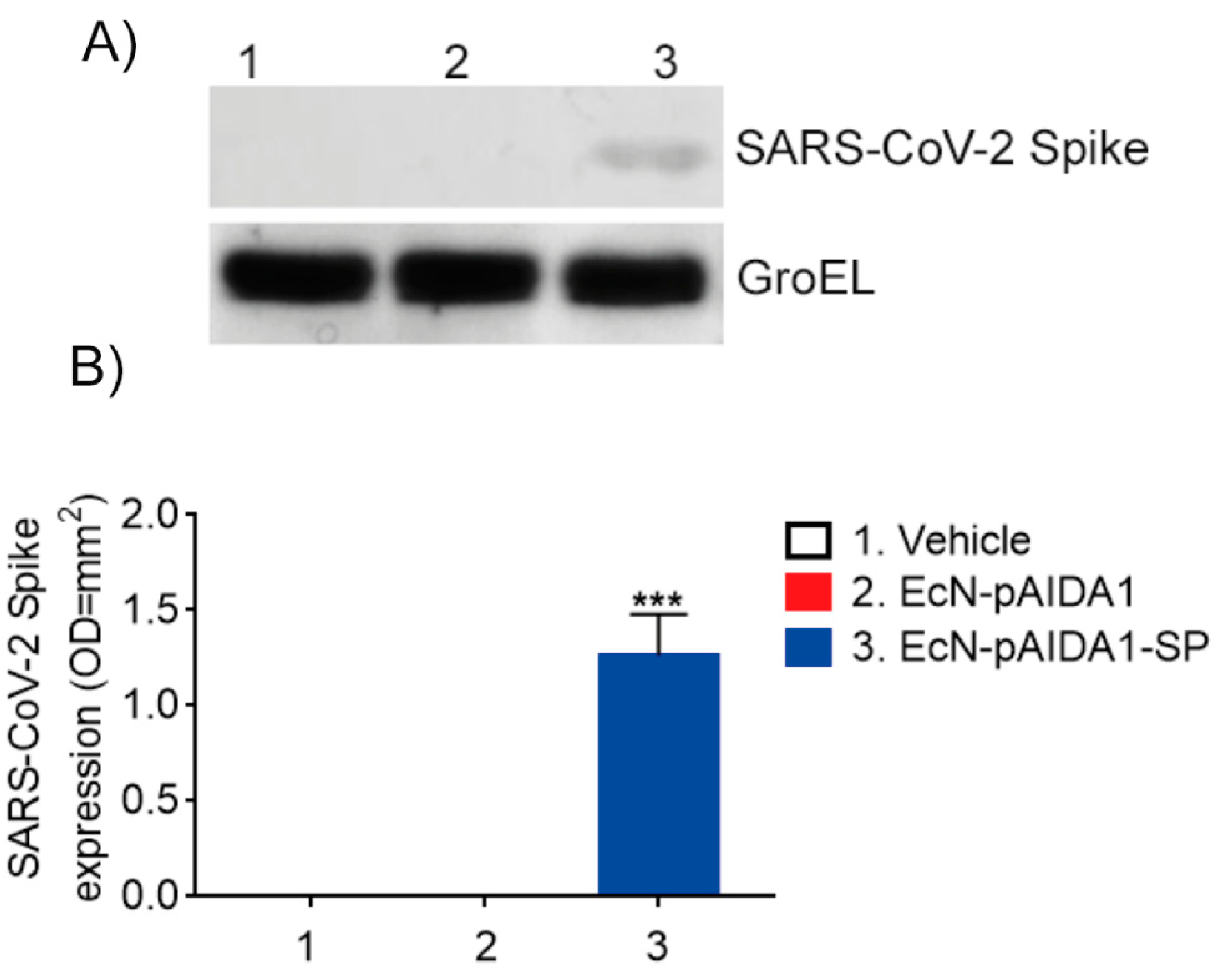
3.2. EcN-pAIDA1-SP Is Capable of Expressing and Delivering the SARS-CoV-2 SP to the Intestinal Site
3.3. Engineered EcN-pAIDA1-SP Elicits a Time-Dependent Increase in Circulating Anti-SARS-CoV-2 SP IgGs
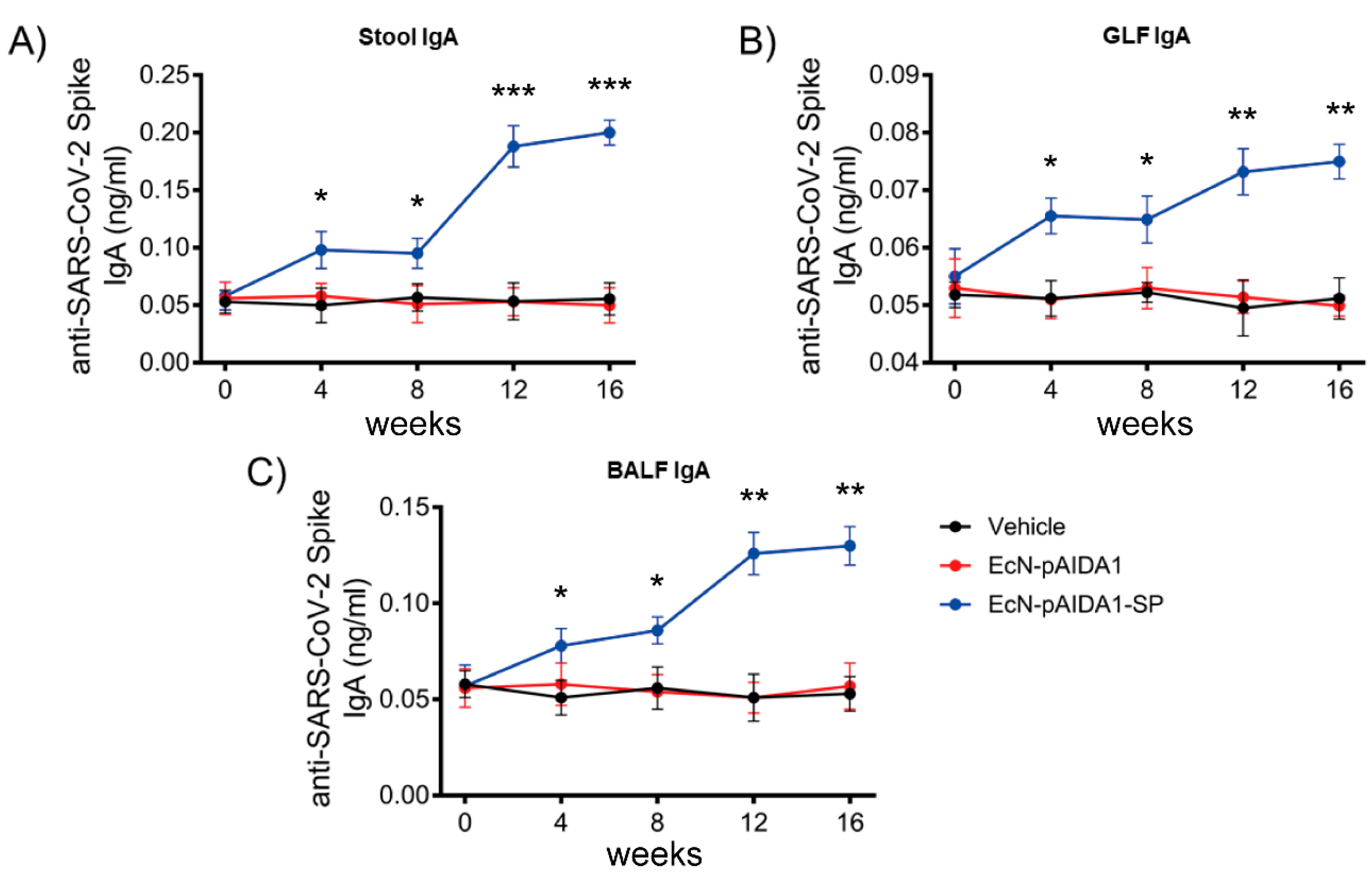
3.4. Engineered EcN-pAIDA1-SP Induces a Specific IgA-Mediated Mucosal Immune Response at Gastrointestinal and Pulmonary Interfaces
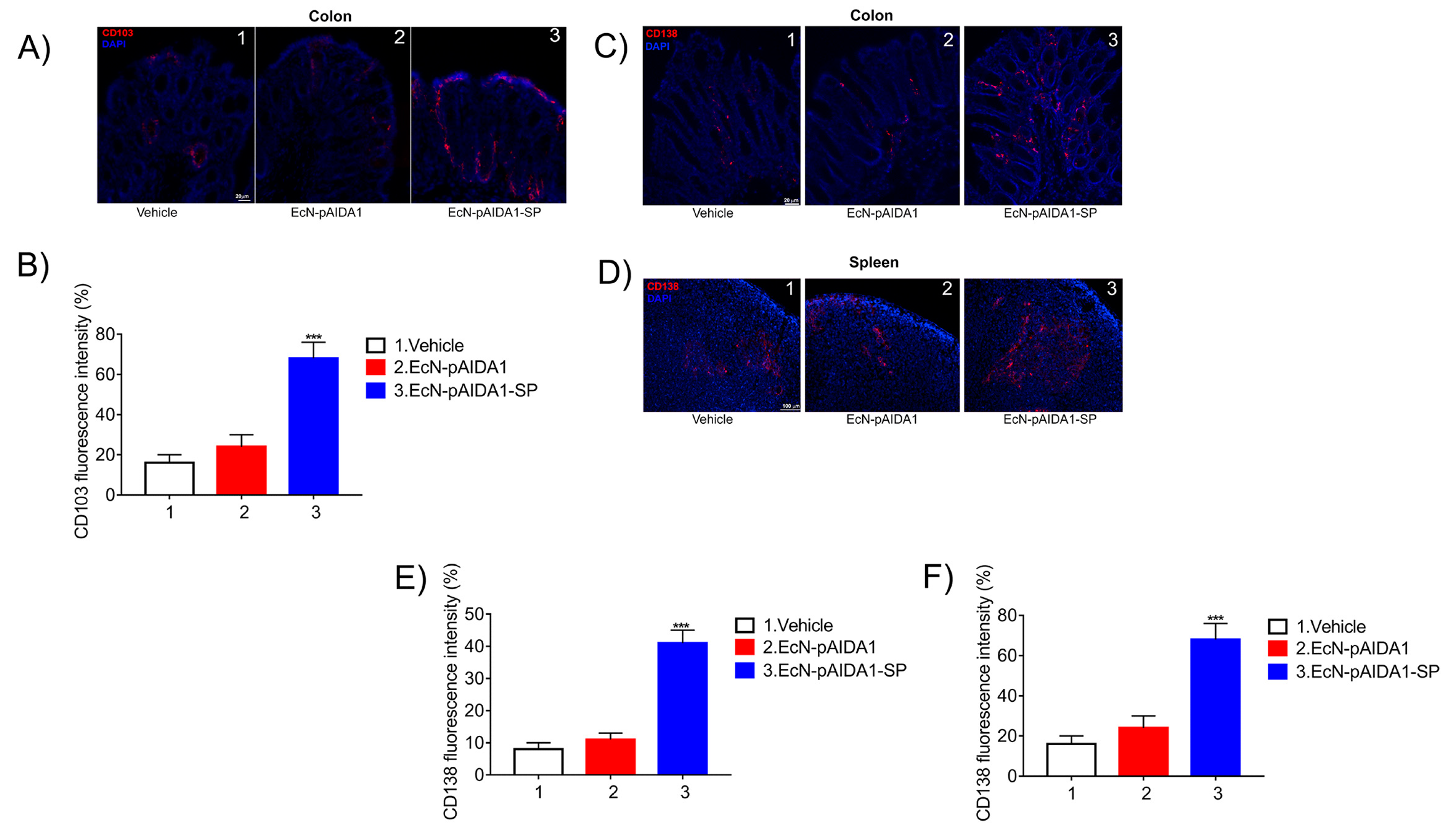
3.5. Engineered EcN-pAIDA1-SP Increases the Dendritic Cell Activity, Increases Plasma Cell Numbers in Both Colon and Spleen
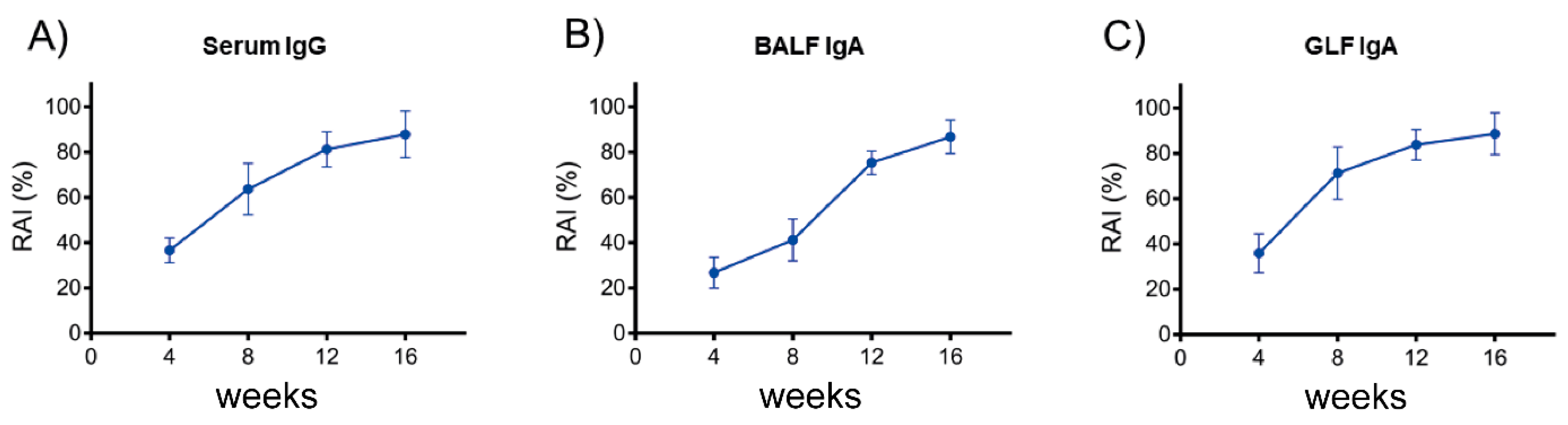
3.6. Anti-SARS-CoV-2 SP IgG and IgA Exhibit a High Relative Avidity Index (RAI) for SP
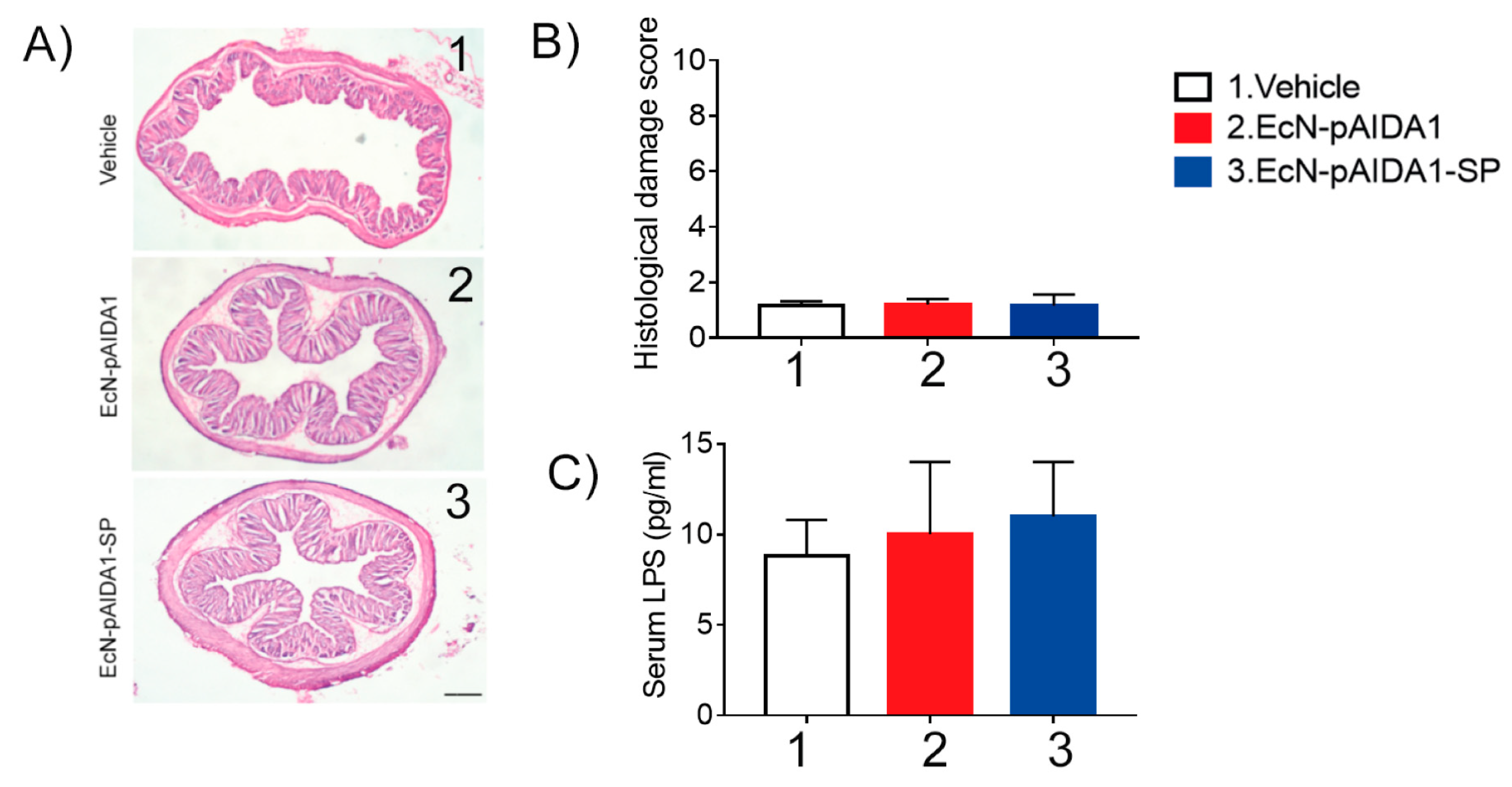
3.7. EcN-pAIDA1-SP Immunization Did Not Induce Side Effects or Affect Mice Survival
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Brüssow, H.; Zuber, S. Can a combination of vaccination and face mask wearing contain the COVID-19 pandemic? Microb. Biotechnol. 2021, 15, 721–737. [Google Scholar] [CrossRef] [PubMed]
- Araf, Y.; Akter, F.; Tang, Y.; Fatemi, R.; Alam Parvez, S.; Zheng, C.; Hossain, G. Omicron variant of SARS-CoV-2: Genomics, transmissibility, and responses to current COVID-19 vaccines. J. Med. Virol. 2022, 94, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Azzi, L.; Dalla Gasperina, D.; Veronesi, G.; Shallak, M.; Ietto, G.; Iovino, D.; Baj, A.; Gianfagna, F.; Maurino, V.; Focosi, D.; et al. Mucosal immune response in BNT162b2 COVID-19 vaccine recipients. eBioMedicine 2021, 75, 103788. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Kalkstein, N.; Mizrahi, B.; Alroy-Preis, S.; Ash, N.; Milo, R.; et al. Protection of BNT162b2 Vaccine Booster against COVID-19 in Israel. N. Engl. J. Med. 2021, 385, 1393–1400. [Google Scholar] [CrossRef]
- Lucero-Prisno, D.E.; Ogunkola, I.O.; Imo, U.F.; Adebisi, Y.A. Who Will Pay for the COVID-19 Vaccines for Africa? Am. J. Trop. Med. Hyg. 2021, 104, 794–796. [Google Scholar] [CrossRef]
- Aggarwal, N.; Breedon, A.M.E.; Davis, C.M.; Hwang, I.Y.; Chang, M.W. Engineering probiotics for therapeutic applications: Recent examples and translational outlook. Curr. Opin. Biotechnol. 2020, 65, 171–179. [Google Scholar] [CrossRef]
- Jiang, B.; Li, Z.; Ou, B.; Duan, Q.; Zhu, G. Targeting ideal oral vaccine vectors based on probiotics: A systematical view. Appl. Microbiol. Biotechnol. 2019, 103, 3941–3953. [Google Scholar] [CrossRef]
- Baker, P.J. Advantages of an Oral Vaccine to Control the COVID-19 Pandemic. Am. J. Med. 2021, 135, 133–134. [Google Scholar] [CrossRef]
- Blount, Z.D.; Michigan State University, United States. The unexhausted potential of E. coli. eLife 2015, 4, e05826. [Google Scholar] [CrossRef]
- Kamionka, M. Engineering of Therapeutic Proteins Production in Escherichia coli. Curr. Pharm. Biotechnol. 2011, 12, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-J.; Lin, H.; Yang, X. Industrial production of recombinant therapeutics in Escherichia coli and its recent advancements. J. Ind. Microbiol. Biotechnol. 2012, 39, 383–399. [Google Scholar] [CrossRef]
- Maeda, D.L.N.F.; Tian, D.; Yu, H.; Dar, N.; Rajasekaran, V.; Meng, S.; Mahsoub, H.M.; Sooryanarain, H.; Wang, B.; Heffron, C.L.; et al. Killed whole-genome reduced-bacteria surface-expressed coronavirus fusion peptide vaccines protect against disease in a porcine model. Proc. Natl. Acad. Sci. USA 2021, 118, e2025622118. [Google Scholar] [CrossRef] [PubMed]
- Samrat, S.K.; Tharappel, A.M.; Li, Z.; Li, H. Prospect of SARS-CoV-2 spike protein: Potential role in vaccine and therapeutic development. Virus Res. 2020, 288, 198141. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer Membrane Vesicles: Current Status and Future Direction of These Novel Vaccine Adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [Green Version]
- Zariri, A.; van der Ley, P. Biosynthetically engineered lipopolysaccharide as vaccine adjuvant. Expert Rev. Vaccines 2015, 14, 861–876. [Google Scholar] [CrossRef]
- Oberhettinger, P.; Schütz, M.; Leo, J.C.; Heinz, N.; Berger, J.; Autenrieth, I.B.; Linke, D. Intimin and Invasin Export Their C-Terminus to the Bacterial Cell Surface Using an Inverse Mechanism Compared to Classical Autotransport. PLoS ONE 2012, 7, e47069. [Google Scholar] [CrossRef]
- Jarmander, J.; Gustavsson, M.; Do, T.-H.; Samuelson, P.; Larsson, G. A dual tag system for facilitated detection of surface expressed proteins in Escherichia coli. Microb. Cell Fact. 2012, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Parasuraman, S.; Raveendran, R.; Kesavan, R. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 2010, 1, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, E.A. Sampling and Preparation of Mouse and Rat Serum. Cold Spring Harb. Protoc. 2017, 2017, prot100271. [Google Scholar] [CrossRef]
- Baz, M.; Samant, M.; Zekki, H.; Tribout-Jover, P.; Plante, M.; Lanteigne, A.-M.; Hamelin, M.-E.; Mallett, C.; Papadopoulou, B.; Boivin, G. Effects of Different Adjuvants in the Context of Intramuscular and Intranasal Routes on Humoral and Cellular Immune Responses Induced by Detergent-Split A/H3N2 Influenza Vaccines in Mice. Clin. Vaccine Immunol. 2012, 19, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Smeekens, J.M.; Johnson-Weaver, B.T.; Hinton, A.L.; Azcarate-Peril, M.A.; Moran, T.P.; Immormino, R.M.; Kesselring, J.R.; Steinbach, E.C.; Orgel, K.A.; Staats, H.F.; et al. Fecal IgA, Antigen Absorption, and Gut Microbiome Composition Are Associated with Food Antigen Sensitization in Genetically Susceptible Mice. Front. Immunol. 2021, 11, 599637. [Google Scholar] [CrossRef]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar]
- Meyer, C.W.; Ootsuka, Y.; Romanovsky, A.A. Body Temperature Measurements for Metabolic Phenotyping in Mice. Front. Physiol. 2017, 8, 520. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.K.S.; Lim, P.; Liu, E.Y.M.; Cheung, J.L.K.; Leung, D.T.M.; Sung, J.J.Y. Antibody Avidity Maturation during Severe Acute Respiratory Syndrome–Associated Coronavirus Infection. J. Infect. Dis. 2005, 192, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.E.; Patel, E.U.; Laeyendecker, O.; Pekosz, A.; Littlefield, K.; Eby, Y.; Fernandez, R.E.; Miller, J.; Kirby, C.S.; Keruly, M.; et al. SARS-CoV-2 Antibody Avidity Responses in COVID-19 Patients and Convalescent Plasma Donors. J. Infect. Dis. 2020, 222, 1974–1984. [Google Scholar] [CrossRef] [PubMed]
- Struck, F.; Schreiner, P.; Staschik, E.; Wochinz-Richter, K.; Schulz, S.; Soutschek, E.; Motz, M.; Bauer, G. Vaccination versus infection with SARS-CoV-2: Establishment of a high avidity IgG response versus incomplete avidity maturation. J. Med. Virol. 2021, 93, 6765–6777. [Google Scholar] [CrossRef]
- Franco-Paredes, C. Transmissibility of SARS-CoV-2 among fully vaccinated individuals. Lancet Infect. Dis. 2022, 22, 16. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community transmission and viral load kinetics of the SARS-CoV-2 delta (B.1.617.2) variant in vaccinated and unvaccinated individuals in the UK: A prospective, longitudinal, cohort study. Lancet Infect. Dis. 2021, 22, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Kazemifard, N.; Dehkohneh, A.; Ghavami, S.B. Probiotics and probiotic-based vaccines: A novel approach for improving vaccine efficacy. Front. Med. 2022, 9, 940454. [Google Scholar] [CrossRef] [PubMed]
- Taghinezhad-S, S.; Mohseni, A.; Bermúdez-Humarán, L.; Casolaro, V.; Cortes-Perez, N.; Keyvani, H.; Simal-Gandara, J. Probiotic-Based Vaccines May Provide Effective Protection against COVID-19 Acute Respiratory Disease. Vaccines 2021, 9, 466. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, T.; Zhou, L.; Sun, J.; Li, Y.; Nie, M.; Xiong, H.; Zhu, Y.; Xue, W.; Wu, Y.; et al. A Bacterially Expressed SARS-CoV-2 Receptor Binding Domain Fused with Cross-Reacting Material 197 A-Domain Elicits High Level of Neutralizing Antibodies in Mice. Front. Microbiol. 2022, 13, 854630. [Google Scholar] [CrossRef]
- Lamichhane, A.; Azegami, T.; Kiyono, H. The mucosal immune system for vaccine development. Vaccine 2014, 32, 6711–6723. [Google Scholar] [CrossRef] [Green Version]
- Fenton, T.M.; Jørgensen, P.; Niss, K.; Rubin, S.; Mörbe, U.; Riis, L.B.; Da Silva, C.; Plumb, A.; Vandamme, J.; Jakobsen, H.L.; et al. Immune Profiling of Human Gut-Associated Lymphoid Tissue Identifies a Role for Isolated Lymphoid Follicles in Priming of Region-Specific Immunity. Immunity 2020, 52, 557–570.e6. [Google Scholar] [CrossRef]
- Fröberg, J.; Diavatopoulos, D.A. Mucosal immunity to severe acute respiratory syndrome coronavirus 2 infection. Curr. Opin. Infect. Dis. 2021, 34, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Corthésy, B. Multi-Faceted Functions of Secretory IgA at Mucosal Surfaces. Front. Immunol. 2013, 4, 185. [Google Scholar] [CrossRef] [Green Version]
- Dobaño, C.; Sanz, H.; Sorgho, H.; Dosoo, D.; Mpina, M.; Ubillos, I.; Aguilar, R.; Ford, T.; Díez-Padrisa, N.; Williams, N.A.; et al. Concentration and avidity of antibodies to different circumsporozoite epitopes correlate with RTS, S/AS01E malaria vaccine efficacy. Nat. Commun. 2019, 10, 2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westendorf, A.M.; Gunzer, F.; Deppenmeier, S.; Tapadar, D.; Hunger, J.K.; Schmidt, M.A.; Buer, J.; Bruder, D. Intestinal immunity of Escherichia coliNISSLE 1917: A safe carrier for therapeutic molecules. FEMS Immunol. Med. Microbiol. 2005, 43, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Dubbert, S.; Klinkert, B.; Schimiczek, M.; Wassenaar, T.M.; von Bünau, R. No genotoxicity is detectable for Escherichia coli strain Nissle 1917 by standard in vitro and in vivo tests. Eur. J. Microbiol. Immunol. 2020, 10, 11–19. [Google Scholar] [CrossRef]
- Smith, H.W. Survival of orally administered E. coli K12 in alimentary tract of man. Nature 1975, 255, 500–502. [Google Scholar] [CrossRef]
- Sturm, A.; Rilling, K.; Baumgart, D.C.; Gargas, K.; Abou-Ghazalé, T.; Raupach, B.; Eckert, J.; Schumann, R.R.; Enders, C.; Sonnenborn, U.; et al. Escherichia coli Nissle 1917 Distinctively Modulates T-Cell Cycling and Expansion via Toll-Like Receptor 2 Signaling. Infect. Immun. 2005, 73, 1452–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebichavsky, I.; Splichal, I.; Rada, V.; Splichalova, A. Modulation of natural immunity in the gut by Escherichia coli strain Nissle 1917. Nutr. Rev. 2010, 68, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Shao, L.; Kandasamy, S.; Fischer, D.D.; Rauf, A.; Langel, S.N.; Chattha, K.S.; Kumar, A.; Huang, H.-C.; Rajashekara, G.; et al. Escherichia coliNissle 1917 protects gnotobiotic pigs against human rotavirus by modulating pDC and NK-cell responses. Eur. J. Immunol. 2016, 46, 2426–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryayev, M.L.; Senkivska, L.I.; Bredeleva, N.K.; Talashova, I.V. Prophylaxis of acute respiratory infections via improving the immune system in late preterm newborns with E. coli strain Nissle 1917: A controlled pilot trial. Pilot Feasibility Stud. 2018, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Vlasova, A.N.; Takanashi, S.; Miyazaki, A.; Rajashekara, G.; Saif, L.J. How the gut microbiome regulates host immune responses to viral vaccines. Curr. Opin. Virol. 2019, 37, 16–25. [Google Scholar] [CrossRef]
- Louis, P.; Galinski, E.A. Survival of Escherichia coli during drying and storage in the presence of compatible solutes. Appl. Microbiol. Biotechnol. 1994, 41, 684–688. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarnelli, G.; Del Re, A.; Pesce, M.; Lu, J.; Esposito, G.; Sanseverino, W.; Corpetti, C.; Basili Franzin, S.; Seguella, L.; Palenca, I.; et al. Oral Immunization with Escherichia coli Nissle 1917 Expressing SARS-CoV-2 Spike Protein Induces Mucosal and Systemic Antibody Responses in Mice. Biomolecules 2023, 13, 569. https://doi.org/10.3390/biom13030569
Sarnelli G, Del Re A, Pesce M, Lu J, Esposito G, Sanseverino W, Corpetti C, Basili Franzin S, Seguella L, Palenca I, et al. Oral Immunization with Escherichia coli Nissle 1917 Expressing SARS-CoV-2 Spike Protein Induces Mucosal and Systemic Antibody Responses in Mice. Biomolecules. 2023; 13(3):569. https://doi.org/10.3390/biom13030569
Chicago/Turabian StyleSarnelli, Giovanni, Alessandro Del Re, Marcella Pesce, Jie Lu, Giovanni Esposito, Walter Sanseverino, Chiara Corpetti, Silvia Basili Franzin, Luisa Seguella, Irene Palenca, and et al. 2023. "Oral Immunization with Escherichia coli Nissle 1917 Expressing SARS-CoV-2 Spike Protein Induces Mucosal and Systemic Antibody Responses in Mice" Biomolecules 13, no. 3: 569. https://doi.org/10.3390/biom13030569
APA StyleSarnelli, G., Del Re, A., Pesce, M., Lu, J., Esposito, G., Sanseverino, W., Corpetti, C., Basili Franzin, S., Seguella, L., Palenca, I., Rurgo, S., De Palma, F. D. E., Zilli, A., & Esposito, G. (2023). Oral Immunization with Escherichia coli Nissle 1917 Expressing SARS-CoV-2 Spike Protein Induces Mucosal and Systemic Antibody Responses in Mice. Biomolecules, 13(3), 569. https://doi.org/10.3390/biom13030569