

Maternal Tryptophan Catabolites and Insulin Resistance Parameters in Preeclampsia

 ,
,  and
and 
Abstract
:1. Introduction
2. Subjects and Methods
2.1. Subjects
2.1.1. Patients
2.1.2. Controls
2.1.3. Exclusion Criteria
2.2. Methods
2.2.1. Biomarkers Assays
2.2.2. Statistical Analysis
3. Results
3.1. Comparison of Demographic and Clinical Data between PE and Control Group
3.2. Comparison of the TRYCATs between PE and Control Group
3.3. Comparison of IR Parameters between the PE and Control Group
3.4. Intercorrelation between TRYCATs and Other Parameters
3.5. ROC of the IR Parameters
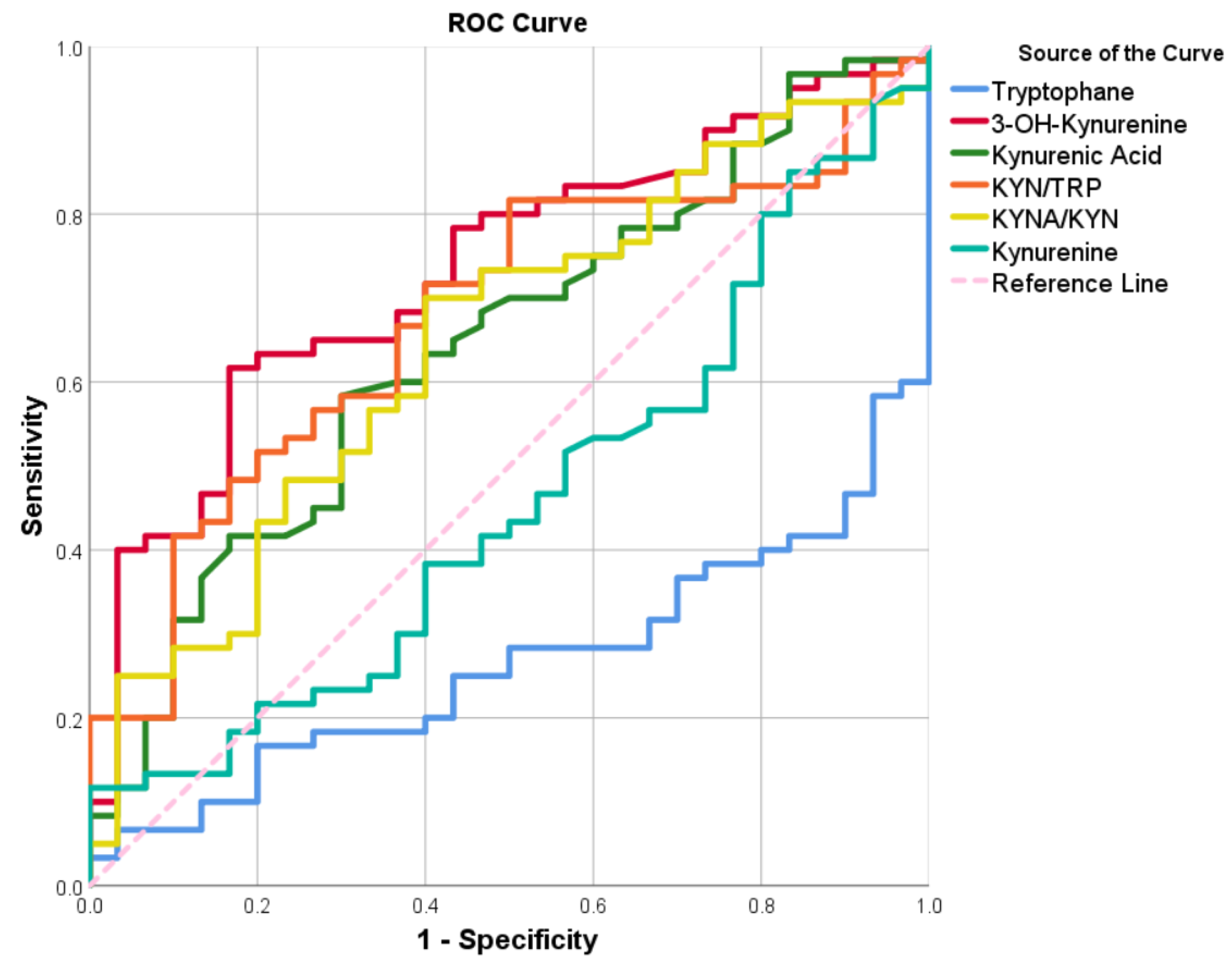
3.6. ROC of the TRYCATs Parameters
4. Discussion
5. Limitations of the Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banu, S.; Akhtary, S.; Pervin, R. Obstetric Outcomes of Pregnant Women with Eclampsia: A study in a Tertiary Care Hospital. Sch. Int. J. Obs. Gynecol. 2022, 5, 147–150. [Google Scholar]
- Tossetta, G.; Fantone, S.; Balercia, G.; Ciavattini, A.; Giannubilo, S.R.; Marzioni, D. Preeclampsia and severe acute respiratory syndrome coronavirus 2 infection: A systematic review. J. Hypertens. 2022, 40, 1629–1638. [Google Scholar] [CrossRef]
- Tanner, M.S.; Davey, M.-A.; Mol, B.W.; Rolnik, D.L. The evolution of the diagnostic criteria of preeclampsia-eclampsia. Am. J. Obstet. Gynecol. 2022, 226, S835–S843. [Google Scholar] [CrossRef]
- Phipps, E.; Prasanna, D.; Brima, W.; Jim, B. Preeclampsia: Updates in Pathogenesis, Definitions, and Guidelines. Clin. J. Am. Soc. Nephrol. 2016, 11, 1102–1113. [Google Scholar] [CrossRef]
- Tsigas, E.Z. The Preeclampsia Foundation: The voice and views of the patient and her family. Am. J. Obstet. Gynecol. 2022, 226, S1254–S1264.e1251. [Google Scholar] [CrossRef] [PubMed]
- Thilaganathan, B.; Kalafat, E. Cardiovascular System in Preeclampsia and Beyond. Hypertension 2019, 73, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Ventura, V.; Li, Y.; Pasupuleti, V.; Roman, Y.M.; Hernandez, A.V.; Pérez-López, F.R. Effects of preeclampsia and eclampsia on maternal metabolic and biochemical outcomes in later life: A systematic review and meta-analysis. Metabolism 2020, 102, 154012. [Google Scholar] [CrossRef]
- Kaaja, R.; Laivuori, H.; Laakso, M.; Tikkanen, M.J.; Ylikorkala, O. Evidence of a state of increased insulin resistance in preeclampsia. Metabolism 1999, 48, 892–896. [Google Scholar] [CrossRef]
- Roberts, J.M.; Gammill, H. Insulin Resistance in Preeclampsia. Hypertension 2006, 47, 341–342. [Google Scholar] [CrossRef]
- Anim-Nyame, N.; Gamble, J.; Sooranna, S.R.; Johnson, M.R.; Steer, P.J. Relationship between insulin resistance and tissue blood flow in preeclampsia. J. Hypertens. 2015, 33, 1057–1063. [Google Scholar] [CrossRef]
- Hauth, J.C.; Clifton, R.G.; Roberts, J.M.; Myatt, L.; Spong, C.Y.; Leveno, K.J.; Varner, M.W.; Wapner, R.J.; Thorp, J.M.; Mercer, B.M.; et al. Maternal insulin resistance and preeclampsia. Am. J. Obstet. Gynecol. 2011, 204, 327.e321–327.e326. [Google Scholar] [CrossRef] [PubMed]
- Garcómez, E.; Bobadilla-Bravo, M.; Díaz-Díaz, E.; Vázquez-Martínez, E.R.; Nava-Salazar, S.; Torres-Ramos, Y.; Garc-Romero, C.S.; Camacho-Arroyo, I.; Cerbón, M. High plasmatic levels of advanced glycation end products are associated with metabolic alterations and insulin resistance in preeclamptic women. Curr. Mol. Med. 2020, 20, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Coppens, V.; Verkerk, R.; Morrens, M. Tracking TRYCAT: A critical appraisal of kynurenine pathway quantifications in blood. Front. Pharmacol. 2022, 13, 825948. [Google Scholar] [CrossRef]
- Schwarcz, R. Kynurenines and Glutamate: Multiple Links and Therapeutic Implications. Adv. Pharmacol. (San Diego Calif.) 2016, 76, 13–37. [Google Scholar] [CrossRef]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging Roles for Serotonin in Regulating Metabolism: New Implications for an Ancient Molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Kanova, M.; Kohout, P. Tryptophan: A Unique Role in the Critically Ill. Int. J. Mol. Sci. 2021, 22, 11714. [Google Scholar] [CrossRef] [PubMed]
- Allegri, G.; Costa, C.V.; Ragazzi, E.; Steinhart, H.; Laresio, L. Developments in Tryptophan and Serotonin Metabolism; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 527. [Google Scholar]
- Kudo, Y.; Boyd, C.A.; Sargent, I.L.; Redman, C.W. Decreased tryptophan catabolism by placental indoleamine 2,3-dioxygenase in preeclampsia. Am. J. Obs. Gynecol. 2003, 188, 719–726. [Google Scholar] [CrossRef]
- Abad, C.; Karahoda, R.; Kastner, P.; Portillo, R.; Horackova, H.; Kucera, R.; Nachtigal, P.; Staud, F. Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit during Gestation. Int. J. Mol. Sci. 2020, 21, 7578. [Google Scholar] [CrossRef]
- Sordillo, P.P.; Sordillo, L.A. Chapter 7—Tryptophan, after inflammatory cytokine stimulation, determines plaque vulnerability and risk of myocardial infarction. In Biophotonics, Tryptophan and Disease; Sordillo, L.A., Sordillo, P.P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 81–91. [Google Scholar]
- Worton, S.A.; Greenwood, S.L.; Wareing, M.; Heazell, A.E.P.; Myers, J. The kynurenine pathway; A new target for treating maternal features of preeclampsia? Placenta 2019, 84, 44–49. [Google Scholar] [CrossRef]
- Oh, C.-M.; Namkung, J.; Go, Y.; Shong, K.E.; Kim, K.; Kim, H.; Park, B.-Y.; Lee, H.W.; Jeon, Y.H.; Song, J. Regulation of systemic energy homeostasis by serotonin in adipose tissues. Nat. Commun. 2015, 6, 6794. [Google Scholar] [CrossRef]
- Gumusoglu, S.; Scroggins, S.; Vignato, J.; Santillan, D.; Santillan, M. The Serotonin-Immune Axis in Preeclampsia. Curr. Hypertens. Rep. 2021, 23, 37. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.; Vidaeff, A.; Pettker, C.M.; Simhan, H. Gestational hypertension and preeclampsia. Obstet. Gynecol. 2019, 133, E1–E25. [Google Scholar]
- Al-Hakeim, H.K.; Abdulzahra, M.S. Correlation Between Glycated Hemoglobin and Homa Indices in Type 2 Diabetes Mellitus: Prediction of Beta-Cell Function from Glycated Hemoglobin/Korelacija Između Glikoliziranog Hemoglobina I Homa Indeksa U Dijabetes Melitusu Tipa 2: Predviđanje Funkcije Beta Ćelija Na Osnovu Glikoliziranog Hemoglobina. J. Med. Biochem. 2015, 34, 191–199. [Google Scholar] [PubMed]
- Sangeeta, S. Metformin and pioglitazone in polycystic ovarian syndrome: A comparative study. J. Obstet. Gynecol. India 2012, 62, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Pernicova, I.; Korbonits, M. Metformin—Mode of action and clinical implications for diabetes and cancer. Nat. Rev. Endocrinol. 2014, 10, 143. [Google Scholar] [CrossRef]
- Broekhuizen, M.; Danser, A.H.J.; Reiss, I.K.M.; Merkus, D. The Function of the Kynurenine Pathway in the Placenta: A Novel Pharmacotherapeutic Target? Int. J. Environ. Res. Public. Health 2021, 18, 11545. [Google Scholar] [CrossRef]
- Keaton, S.A.; Heilman, P.; Bryleva, E.Y.; Madaj, Z.; Krzyzanowski, S.; Grit, J.; Miller, E.S.; Jälmby, M.; Kalapotharakos, G.; Racicot, K.; et al. Altered Tryptophan Catabolism in Placentas from Women with Pre-eclampsia. Int. J. Tryptophan Res. 2019, 12, 1178646919840321. [Google Scholar] [CrossRef]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef]
- Senior, J.; Fahim, I.; Sullivan, F.; Robson, J. Possible role of 5-hydroxytryptamine in toxaemia of pregnancy. Lancet 1963, 282, 553–554. [Google Scholar] [CrossRef]
- Wang, Y.; Walsh, S.W. Hormonal and related mechanisms for preeclampsia of pregnancy. Endocrinologist 1997, 7, 8–244. [Google Scholar] [CrossRef]
- Nilsen, R.M.; Bjørke-Monsen, A.L.; Midttun, O.; Nygård, O.; Pedersen, E.R.; Ulvik, A.; Magnus, P.; Gjessing, H.K.; Vollset, S.E.; Ueland, P.M. Maternal tryptophan and kynurenine pathway metabolites and risk of preeclampsia. Obs. Gynecol. 2012, 119, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.J.; Zhou, C.; Wei, Y.Y.; Li, H.H.; Lei, W.; Boeldt, D.S.; Wang, K.; Zheng, J. Differential Distribution of Tryptophan-Metabolites in Fetal and Maternal Circulations during Normotensive and Preeclamptic Pregnancies. Reprod. Sci. (Thousand Oaks Calif.) 2022, 29, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Favennec, M.; Hennart, B.; Caiazzo, R.; Leloire, A.; Yengo, L.; Verbanck, M.; Arredouani, A.; Marre, M.; Pigeyre, M.; Bessede, A.; et al. The kynurenine pathway is activated in human obesity and shifted toward kynurenine monooxygenase activation. Obesity (Silver Spring) 2015, 23, 2066–2074. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.B. Hypothesis kynurenic and quinolinic acids: The main players of the kynurenine pathway and opponents in inflammatory disease. Med. Hypotheses 2018, 118, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Almulla, A.F.; Maes, M. The tryptophan catabolite or kynurenine pathway’s role in major depression. Curr. Top. Med. Chem. 2022, 22, 1731–1735. [Google Scholar] [CrossRef]
- Goda, K.; Hamane, Y.; Kishimoto, R.; Ogishi, Y.J.T. Radical scavenging properties of tryptophan metabolites: Estimation of their radical reactivity. Adv. Exp. Med. Biol. 1999, 467, 397–402. [Google Scholar]
- Maes, M.; Mihaylova, I.; Ruyter, M.D.; Kubera, M.; Bosmans, E.J.N.L. The immune effects of TRYCATs (tryptophan catabolites along the IDO pathway): Relevance for depression—And other conditions characterized by tryptophan depletion induced by inflammation. Neuro Endocrinol. Lett. 2007, 28, 826–831. [Google Scholar]
- Reyes Ocampo, J.; Lugo Huitrón, R.; González-Esquivel, D.; Ugalde-Muñiz, P.; Jiménez-Anguiano, A.; Pineda, B.; Pedraza-Chaverri, J.; Ríos, C.; Perez de la Cruz, V.J.O.M.; Longevity, C. Kynurenines with neuroactive and redox properties: Relevance to aging and brain diseases. Oxidative Med. Cell. Longev. 2014, 2014, 646909. [Google Scholar] [CrossRef]
- Cai, Z.; Tian, S.; Klein, T.; Tu, L.; Geenen, L.W.; Koudstaal, T.; van den Bosch, A.E.; de Rijke, Y.B.; Reiss, I.K.M.; Boersma, E.; et al. Kynurenine metabolites predict survival in pulmonary arterial hypertension: A role for IL-6/IL-6Rα. Sci. Rep. 2022, 12, 12326. [Google Scholar] [CrossRef]
- Xu, K.; Liu, H.; Bai, M.; Gao, J.; Wu, X.; Yin, Y. Redox Properties of Tryptophan Metabolism and the Concept of Tryptophan Use in Pregnancy. Int. J. Mol. Sci. 2017, 18, 1595. [Google Scholar] [CrossRef]
- Keaton, S.A. The Role of Inflammation and the Kynurenine Pathway in Mood Disorders and Pregnancy; Michigan State University: East Lansing, MI, USA, 2019. [Google Scholar]
- Badawy, A.A.-B.; Namboodiri, A.M.; Moffett, J.R. The end of the road for the tryptophan depletion concept in pregnancy and infection. Clin. Sci. 2016, 130, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A. Plasma free tryptophan revisited: What you need to know and do before measuring it. J. Psychopharmacol. (Oxf. Engl.) 2010, 24, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Broekhuizen, M.; Klein, T.; Hitzerd, E.; de Rijke, Y.B.; Schoenmakers, S.; Sedlmayr, P.; Danser, A.H.J.; Merkus, D.; Reiss, I.K.M. l-Tryptophan–Induced Vasodilation Is Enhanced in Preeclampsia. Hypertension 2020, 76, 184–194. [Google Scholar] [CrossRef] [PubMed]
- van Zundert, S.K.; Broekhuizen, M.; Smit, A.J.; van Rossem, L.; Mirzaian, M.; Willemsen, S.P.; Danser, A.J.; De Rijke, Y.B.; Reiss, I.K.; Merkus, D. The Role of the Kynurenine Pathway in the (Patho) physiology of Maternal Pregnancy and Fetal Outcomes: A Systematic Review. Int. J. Tryptophan Res. 2022, 15, 11786469221135545. [Google Scholar] [CrossRef]
- Kudo, Y.; Boyd, C.; Spyropoulou, I.; Redman, C.; Takikawa, O.; Katsuki, T.; Hara, T.; Ohama, K.; Sargent, I.J.J.o.r.i. Indoleamine 2,3-dioxygenase: Distribution and function in the developing human placenta. J. Reprod. Immunol. 2004, 61, 87–98. [Google Scholar] [CrossRef]
- Popkov, S.A. Kynurenine Metabolic Pathway of Tryptophan Catabolism in Healthy and Pathological Pregnancy. Available online: https://www.priory.com/med/kynurenine.htm (accessed on 2 June 2023).
- Wang, Q.; Liu, D.; Song, P.; Zou, M.H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front. Biosci. 2015, 20, 1116–1143. [Google Scholar] [CrossRef]
- Maes, M.; Leonard, B.; Myint, A.; Kubera, M.; Verkerk, R. The new ‘5-HT’hypothesis of depression: Cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to the onset of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 702–721. [Google Scholar]
- Smith, A.J.; Smith, R.A.; Stone, T.W. 5-Hydroxyanthranilic acid, a tryptophan metabolite, generates oxidative stress and neuronal death via p38 activation in cultured cerebellar granule neurones. Neurotox. Res. 2009, 15, 303–310. [Google Scholar] [CrossRef]
- Holtze, M.; Saetre, P.; Engberg, G.; Schwieler, L.; Werge, T.; Andreassen, O.A.; Hall, H.; Terenius, L.; Agartz, I.; Jönsson, E.G.J.J.o.P.; et al. Kynurenine 3-monooxygenase polymorphisms: Relevance for kynurenic acid synthesis in patients with schizophrenia and healthy controls. J. Psychiatry Neurosci. 2012, 37, 53–57. [Google Scholar] [CrossRef]
- Marić, I.; Contrepois, K.; Moufarrej, M.N.; Stelzer, I.A.; Feyaerts, D.; Han, X.; Tang, A.; Stanley, N.; Wong, R.J.; Traber, G.M.; et al. Early prediction and longitudinal modeling of preeclampsia from multiomics. Patterns 2022, 3, 100655. [Google Scholar] [CrossRef]
- Stone, T.W.; Darlington, L.G. Endogenous kynurenines as targets for drug discovery and development. Nat. Rev. Drug Discov. 2002, 1, 609–620. [Google Scholar] [CrossRef]
- Zavalza-Gómez, A.B. Obesity and oxidative stress: A direct link to preeclampsia? Arch. Gynecol. Obstet. 2011, 283, 415–422. [Google Scholar] [CrossRef]
- Oxenkrug, G.F. Role of kynurenine pathway in insulin resistance: Toward kynurenine hypothesis of insulin resistance and diabetes. In Targeting the Broadly Pathogenic Kynurenine Pathway; Springer: Cham, Switzerland, 2015; pp. 169–178. [Google Scholar]
- Sordillo, P.P.; Sordillo, L.A. Chapter 10—Excess activity of 3-hydroxykynurenine, quinolinic acid, and other toxic tryptophan metabolites in neurogenerative diseases and other protein misfolding diseases. In Biophotonics, Tryptophan and Disease; Sordillo, L.A., Sordillo, P.P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 115–130. [Google Scholar]
- Sedlmayr, P.; Blaschitz, A.; Stocker, R. The role of placental tryptophan catabolism. Front. Immunol. 2014, 5, 230. [Google Scholar] [CrossRef]
- Lakshmi, P.S.; Shanmuga, P.V.; Suganthy, K.; Kalaiselvi, K. A Study of Association of Insulin Resistance with Preeclampsia. J. Evol. Med. Dent. Sci. 2020, 9, 527–532. [Google Scholar]
- Seely, E.W.; Solomon, C.G. Insulin resistance and its potential role in pregnancy-induced hypertension. J. Clin. Endocrinol. Metab. 2003, 88, 2393–2398. [Google Scholar] [CrossRef]
- Kaaja, R. Insulin resistance syndrome in preeclampsia. Semin. Reprod. Endocrinol. 1998, 16, 41–46. [Google Scholar] [CrossRef]
- Tezikov, Y.V.; Lipatov, I.S.; Azamatov, A.R.; Tyutyunnik, V.L.; Kan, N.E.; Zumorina, E.M.; Kuzmina, A.I. Preeclampsia as a separate gestational clinical and pathogenetic form of insulin resistance syndrome. Obstet. Gynecol. 2022, 4, 64–74. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Control (n = 30) | PE (n = 60) | F/χ2 | p |
|---|---|---|---|---|
| Age years | 27.670 ± 5.933 | 29.230 ± 6.649 | 1.190 | 0.278 |
| Residency Urban/Rural | 13/17 | 27/33 | 0.023 | 0.881 |
| Previous abortion No/Yes | 22/8 | 30/30 | 4.464 | 0.035 |
| Nullipara No/Yes | 24/6 | 53/7 | 1.124 | 0.289 |
| Multipara No/Yes | 7/23 | 7/53 | 2.072 | 0.150 |
| Family history No/Yes | 29/1 | 40/20 | 10.857 | 0.017 |
| SBP mmHg | 122.200 ± 2.075 | 146.900 ± 17.445 | 59.389 | <0.001 |
| DBP mmHg | 82.133 ± 2.030 | 89.033 ± 9.671 | 14.864 | <0.001 |
| Gestational age Wks. | 30.370 ± 4.965 | 30.350 ± 3.0360 | 0.001 | 0.984 |
| Gravidity | 3.00 (1.00–4.00) | 3.00 (1.25–4.75) | - | 0.332 |
| Parity | 2.00 (0–3.00) | 2.00 (0.25–3.75) | - | 0.332 |
| Number of children | 2.00 (1.00–3.00) | 3.00 (1.00–4.00) | - | 0.092 |
| Cesarean delivery | 0(0–1) | 1 (0.5–2.75) | - | 0.007 |
| Natural delivery | 1.50 (0.75–3.00) | 1.00 (1.00–3.00) | - | 0.542 |
| Variables | Control (n = 30) | PE (n = 60) | p |
|---|---|---|---|
| TRP µM | 46.697 (43.534–53.376) | 40.484 (36.743–47.486) | <0.001 |
| KYN µM | 1.629 (1.465–1.875) | 1.689 (1.545–1.907) | 0.502 |
| KYNA nM | 41.788 (35.985–45.096) | 42.460 (39.191–51.290) | 0.027 |
| 3-HK nM | 30.464 (28.300–36.082) | 39.814 (31.372–51.656) | <0.001 |
| KYNA/KYN | 25.025 (24.067–26.111) | 24.706 (22.061–28.113) | 0.024 |
| KYN/TRP | 0.034 (0.033–0.040) | 0.042 (0.034–0.049) | 0.006 |
| Variables | Control (n = 30) | PE (n = 60) | p |
|---|---|---|---|
| Glucose mM | 5.241 ± 0.391 | 5.899 ± 0.914 | <0.001 |
| Insulin pM | 52.097 (39.429–60.620) | 65.439 (51.003–82.088) | 0.001 |
| HOMA%B | 80.300 (72.675–100.150) | 81.600 (67.925–102.150) | 0.742 |
| HOMA%S | 99.750 (88.050–132.700) | 82.750 (65.050–100.475) | <0.001 |
| HOMA2IR | 1.003 (0.754–1.136) | 1.239 (0.995–1.537) | <0.001 |
| I/G | 9.663 (7.724–11.308) | 11.177 (8.660–14.450) | 0.022 |
| Parameters | TRP | 3-HK | KYNA | KYN | KYN/TRP | KYNA/KYN |
|---|---|---|---|---|---|---|
| Residency | −0.019 | −0.089 | 0.087 | 0.022 | 0.002 | 0.046 |
| Abortion | 0.170 | 0.069 | −0.072 | 0.005 | −0.129 | −0.026 |
| Nullipara | −0.067 | −0.153 | −0.100 | −0.085 | 0.033 | −0.060 |
| Multipara | 0.024 | 0.173 | 0.100 | 0.044 | −0.019 | 0.099 |
| Family history | −0.171 | 0.177 | −0.060 | −0.124 | 0.060 | 0.051 |
| Age | 0.114 | 0.020 | 0.084 | −0.013 | −0.080 | 0.114 |
| SBP | −0.216 * | 0.325 ** | 0.125 | −0.036 | 0.172 | 0.134 |
| DBP | −0.219 * | 0.182 | 0.093 | −0.003 | 0.155 | 0.093 |
| Gestational age | −0.081 | −0.205 | −0.114 | 0.003 | 0.071 | −0.060 |
| Gravidity | 0.261 * | 0.162 | 0.061 | 0.140 | −0.172 | −0.005 |
| having a child | 0.203 | 0.131 | 0.087 | 0.133 | −0.132 | 0.000 |
| Cesarean delivery | 0.098 | 0.151 | 0.067 | 0.200 | −0.016 | 0.011 |
| Natural delivery | 0.203 | 0.066 | 0.036 | −0.052 | −0.227 * | 0.016 |
| Parity | 0.261 * | 0.162 | 0.061 | 0.140 | −0.172 | −0.005 |
| Age of Onset | 0.320 * | −0.038 | 0.083 | 0.059 | −0.229 | 0.111 |
| Duration of Symptoms | −0.081 | −0.105 | −0.022 | −0.077 | −0.057 | 0.106 |
| Glucose | −0.011 | 0.129 | 0.139 | −0.039 | 0.041 | 0.143 |
| Insulin pM | −0.205 | 0.096 | 0.005 | −0.120 | 0.091 | 0.187 |
| HOMA%B | −0.109 | 0.037 | −0.153 | −0.066 | 0.004 | 0.013 |
| HOMA%S | 0.191 | −0.090 | −0.034 | 0.129 | −0.080 | −0.212 * |
| HOMA2IR | −0.191 | 0.090 | 0.034 | −0.129 | 0.080 | 0.212 * |
| I/G | −0.188 | 0.080 | −0.074 | −0.117 | 0.058 | 0.141 |
| Variables | Cut off | Sensitivity | Specificity | Youdin J Statistic | p | AUC | 95% CI-AUC |
|---|---|---|---|---|---|---|---|
| I/G nM | 10.189 | 65.0 | 64.3 | 0.29 | <0.001 | 0.748 | 0.532–0.764 |
| HOMA2IR | 1.055 | 65.0 | 63.3 | 0.28 | <0.001 | 0.736 | 0.630–0.841 |
| HOMA%S * | 94.25 | 63.3 | 63.3 | 0.27 | <0.001 | 0.736 | 0.630–0.841 |
| Insulin pM | 54.62 | 63.3 | 63.3 | 0.27 | 0.001 | 0.721 | 0.613–0.828 |
| Glucose mM | 5.48 | 61.7 | 60.0 | 0.22 | 0.001 | 0.719 | 0.616–0.822 |
| HOMA%B | 80.6 | 50.0 | 50.0 | 0 | 0.742 | 0.479 | 0.358–0.599 |
| Variable(s) | Cut off | Sensitivity | Specificity | Youdin J Statistic | p | AUC | 95% CI-AUC |
|---|---|---|---|---|---|---|---|
| Tryptophane * µM | 44.896 | 66.7 | 68.3 | 0.35 | <0.001 | 0.748 | 0.165–0.365 |
| 3-HK nM | 38.491 | 65.0 | 66.7 | 0.32 | <0.001 | 0.734 | 0.629–0.839 |
| KYN/TRP | 0.037 | 65.0 | 64.3 | 0.29 | 0.006 | 0.677 | 0.565–0.790 |
| KYNA/KYN | 24.935 | 60.0 | 60.0 | 0.20 | 0.024 | 0.646 | 0.528–0.764 |
| Kynurenic Acid nM | 41.591 | 60.0 | 60.0 | 0.20 | 0.027 | 0.643 | 0.525–0.762 |
| Kynurenine µM | 1.668 | 53.3 | 53.3 | 0.07 | 0.502 | 0.456 | 0.332–0.581 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasim, Z.A.; Al-Hakeim, H.K.; Zolghadri, S.; Stanek, A. Maternal Tryptophan Catabolites and Insulin Resistance Parameters in Preeclampsia. Biomolecules 2023, 13, 1447. https://doi.org/10.3390/biom13101447
Jasim ZA, Al-Hakeim HK, Zolghadri S, Stanek A. Maternal Tryptophan Catabolites and Insulin Resistance Parameters in Preeclampsia. Biomolecules. 2023; 13(10):1447. https://doi.org/10.3390/biom13101447
Chicago/Turabian StyleJasim, Zainab Abdulameer, Hussein Kadhem Al-Hakeim, Samaneh Zolghadri, and Agata Stanek. 2023. "Maternal Tryptophan Catabolites and Insulin Resistance Parameters in Preeclampsia" Biomolecules 13, no. 10: 1447. https://doi.org/10.3390/biom13101447
APA StyleJasim, Z. A., Al-Hakeim, H. K., Zolghadri, S., & Stanek, A. (2023). Maternal Tryptophan Catabolites and Insulin Resistance Parameters in Preeclampsia. Biomolecules, 13(10), 1447. https://doi.org/10.3390/biom13101447