
Neonatal Maternal Separation Induces Sexual Dimorphism in Brain Development: The Influence on Amino Acid Levels and Cognitive Disorders

 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Drugs
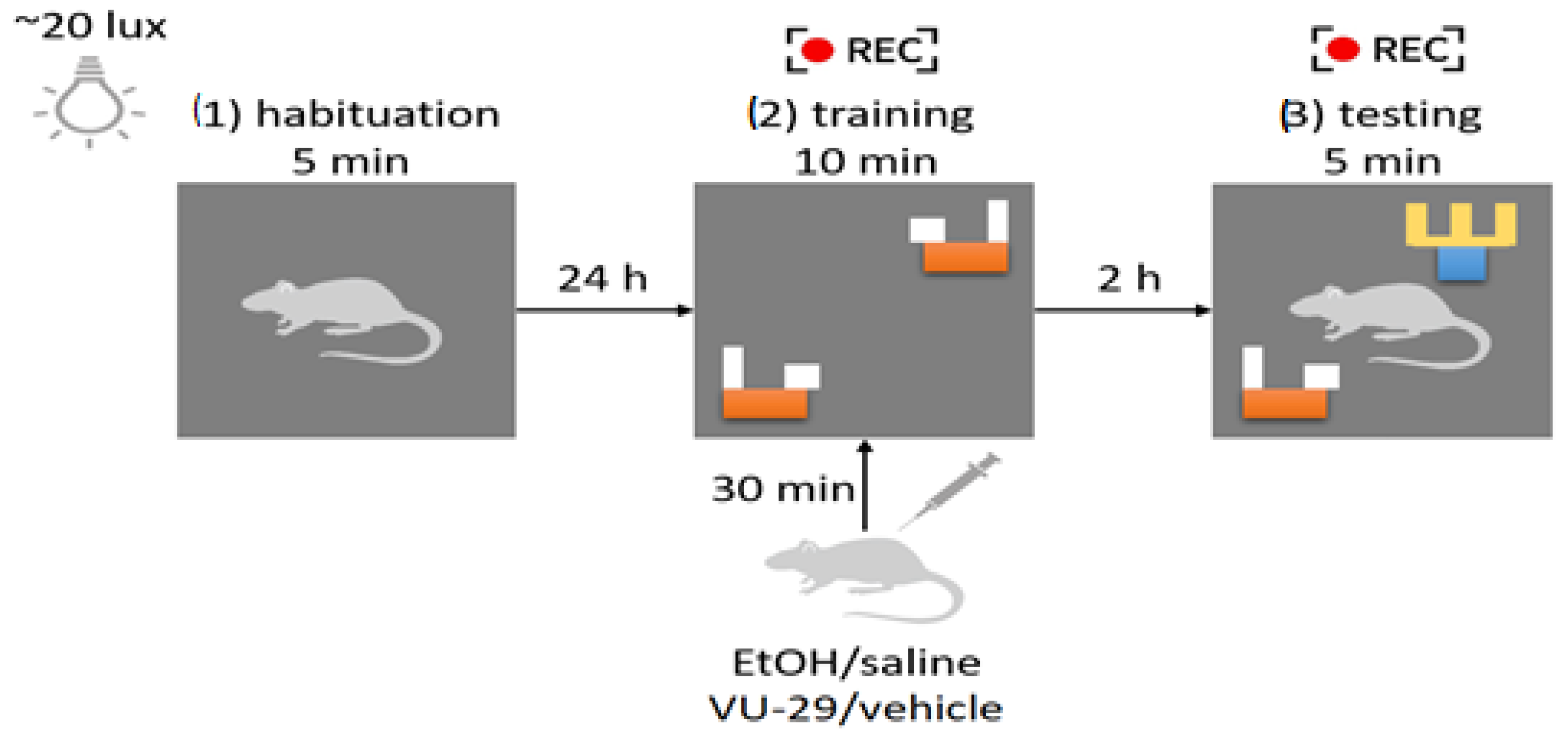
2.3. Maternal Separation Procedure
2.4. Spectral Analysis and Quantification of Neurochemicals in the Hippocampus In Vivo
2.5. Magnetic Resonance Spectroscopy
2.6. Sequence Used for Volumetry Analysis
2.7. Volumetry Analysis
2.8. Novel Object Recognition (NOR)
2.9. Statistical Analysis
3. Results
3.1. The Effect of MS on the Volume of Adult Rat Brain Structures Important for Recognition Memory
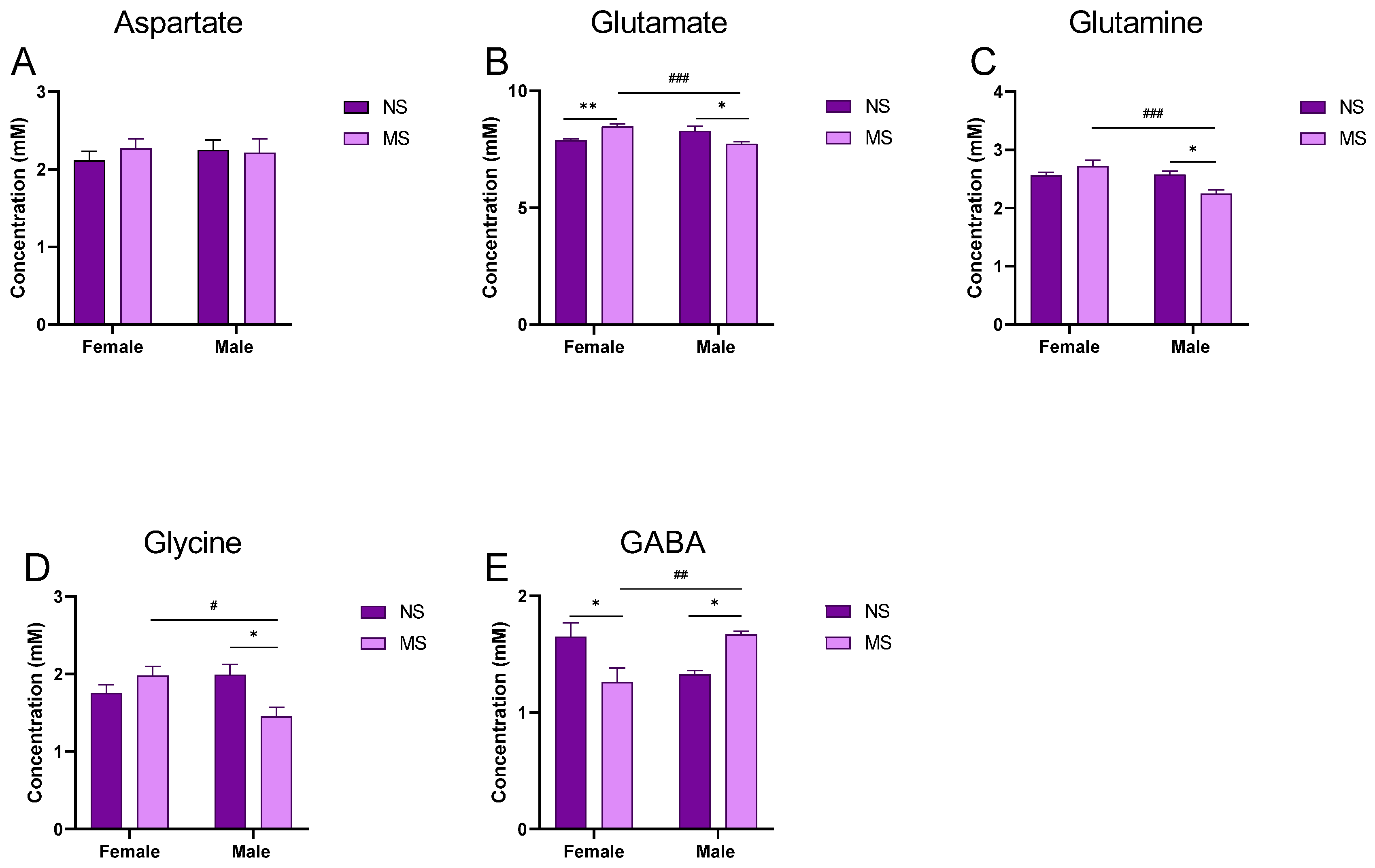
3.2. Impact of MS on the Level of Amino Acids in the Hippocampus of Adult Rats
3.3. Impact of Ethanol on Memory in the NOR Test in Adult Rats Exposed to Repeated MS; Impact of VU-29 on Ethanol-Impaired Memory in MS Rats
3.4. Locomotor Activity in NOR Task
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Di Segni, M.; Andolina, D.; Ventura, R. Long-term effects of early environment on the brain: Lesson from rodent models. Semin. Cell Dev. Biol. 2018, 77, 81–92. [Google Scholar] [CrossRef]
- Huot, R.L.; Plotsky, P.M.; Lenox, R.H.; McNamara, R.K. Neonatal maternal separation reduces hippocampal mossy fiber density in adult Long Evans rats. Brain Res. 2002, 950, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Abraham, A.; Kindler, J.; Helmeke, C.; Braun, K. Exposure to neonatal separation stress alters exploratory behavior and corticotropin releasing factor expression in neurons in the amygdala and hippocampus. Dev. Neurobiol. 2007, 67, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.A.; Jacobvitz, D.; Sroufe, L.A. A developmental investigation of inattentiveness and hyperactivity. Child. Dev. 1995, 66, 37–54. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Effects of adverse experiences for brain structure and function. Biol. Psychiatry 2000, 48, 721–731. [Google Scholar] [CrossRef]
- Oitzl, M.S.; Workel, J.O.; Fluttert, M.; Frösch, F.; De Kloet, E.R. Maternal deprivation affects behaviour from youth to senescence: Amplification of individual differences in spatial learning and memory in senescent Brown Norway rats. Eur. J. Neurosci. 2000, 12, 3771–3780. [Google Scholar] [CrossRef]
- Barnes, L.L.; Wilson, R.S.; Everson-Rose, S.A.; Hayward, M.D.; Evans, D.A.; Mendes de Leon, C.F. Effects of early-life adversity on cognitive decline in older African Americans and whites. Neurology 2012, 79, 2321–2327. [Google Scholar] [CrossRef]
- Pechtel, P.; Pizzagalli, D.A. Effects of early life stress on cognitive and affective function: An integrated review of human literature. Psychopharmacology 2011, 214, 55–70. [Google Scholar] [CrossRef]
- Donley, G.A.R.; Lönnroos, E.; Tuomainen, T.P.; Kauhanen, J. Association of childhood stress with late-life dementia and Alzheimer’s disease: The KIHD study. Eur. J. Public Health 2018, 28, 1069–1073. [Google Scholar] [CrossRef]
- Hoeijmakers, L.; Lesuis, S.L.; Krugers, H.; Lucassen, P.J.; Korosi, A. A preclinical perspective on the enhanced vulnerability to Alzheimer’s disease after early-life stress. Neurobiol. Stress 2018, 8, 172–185. [Google Scholar] [CrossRef]
- Paterniti, S.; Sterner, I.; Caldwell, C.; Bisserbe, J.C. Childhood neglect predicts the course of major depression in a tertiary care sample: A follow-up study. BMC Psychiatry 2017, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Wigger, A.; Neumann, I.D. Periodic maternal deprivation induces gender-dependent alterations in behavioral and neuroendocrine responses to emotional stress in adult rats. Physiol. Behav. 1999, 66, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Kalinichev, M.; Easterling, K.W.; Plotsky, P.M.; Holtzman, S.G. Long-lasting changes in stress-induced corticosterone response and anxiety-like behaviors as a consequence of neonatal maternal separation in Long-Evans rats. Pharmacol. Biochem. Behav. 2002, 73, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Kolb, B. Maternal separation altered behavior and neuronal spine density without influencing amphetamine sensitization. Behav. Brain Res. 2011, 223, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.C.; Ogawa, S. Long-lasting consequences of neonatal maternal separation on social behaviors in ovariectomized female mice. PLoS ONE 2012, 7, e33028. [Google Scholar] [CrossRef]
- Cruz, F.C.; Quadros, I.M.; Planeta Cda, S.; Miczek, K.A. Maternal separation stress in male mice: Long-term increases in alcohol intake. Psychopharmacology 2008, 201, 459–468. [Google Scholar] [CrossRef]
- Chocyk, A.; Dudys, D.; Przyborowska, A.; Maćkowiak, M.; Wędzony, K. Impact of maternal separation on neural cell adhesion molecules expression in dopaminergic brain regions of juvenile, adolescent and adult rats. Pharmacol. Rep. 2010, 62, 1218–1224. [Google Scholar] [CrossRef]
- Levine, S. Maternal and environmental influences on the adrenocortical response to stress in weanling rats. Science 1967, 156, 258–260. [Google Scholar] [CrossRef]
- Grochecki, P.; Smaga, I.; Surowka, P.; Marszalek-Grabska, M.; Kalaba, P.; Dragacevic, V.; Kotlinska, P.; Filip, M.; Lubec, G.; Kotlinska, J.H. Novel dopamine transporter inhibitor, CE-123, ameliorates spatial memory deficits induced by maternal separation in adolescent rats: Impact of sex. Int. J. Mol. Sci. 2022, 23, 10718. [Google Scholar] [CrossRef]
- Banqueri, M.; Gutiérrez-Menéndez, A.; Méndez, M.; Conejo, N.M.; Arias, J.L. Early life stress due to repeated maternal separation alters the working memory acquisition brain functional network. Stress 2021, 24, 87–95. [Google Scholar] [CrossRef]
- Yang, S.; Li, J.; Han, L.; Zhu, G. Early maternal separation promotes apoptosis in dentate gyrus and alters neurological behaviors in adolescent rats. Int. J. Clin. Exp. Pathol. 2017, 10, 10812–10820. [Google Scholar] [PubMed]
- Talani, G.; Biggio, F.; Gorule, A.A.; Licheri, V.; Saolini, E.; Colombo, D.; Sarigu, G.; Petrella, M.; Vedele, F.; Biggio, G.; et al. Sex-dependent changes of hippocampal synaptic plasticity and cognitive performance in C57BL/6J mice exposed to neonatal repeated maternal separation. Neuropharmacology 2023, 222, 109301. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J. Neural Transm. 2014, 121, 799–817. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.J.; Goudet, C. International union of basic and clinical pharmacology. CXI. pharmacology, signaling, and physiology of metabotropic glutamate receptors. Pharmacol. Rev. 2021, 73, 521–569. [Google Scholar] [CrossRef] [PubMed]
- Komuro, H.; Rakic, P. Modulation of neuronal migration by NMDA receptors. Science 1993, 260, 95–97. [Google Scholar] [CrossRef]
- LoTurco, J.J.; Blanton, M.G.; Kriegstein, A.R. Initial expression and endogenous activation of NMDA channels in early neocortical development. J. Neurosci. 1991, 11, 792–799. [Google Scholar] [CrossRef]
- Newell, D.W.; Barth, A.; Ricciardi, T.N.; Malouf, A.T. Glycine causes increased excitability and neurotoxicity by activation of NMDA receptors in the hippocampus. Exp. Neurol. 1997, 145, 235–244. [Google Scholar] [CrossRef]
- Sibson, N.R.; Mason, G.F.; Shen, J.; Cline, G.W.; Herskovits, A.Z.; Wall, J.E.; Behar, K.L.; Rothman, D.L.; Shulman, R.G. In vivo (13)C NMR measurement of neurotransmitter glutamate cycling, anaplerosis and TCA cycle flux in rat brain during. J. Neurochem. 2001, 76, 975–989. [Google Scholar] [CrossRef]
- Hertz, L. The Glutamate-glutamine (GABA) cycle: Importance of late postnatal development and potential reciprocal interactions between biosynthesis and degradation. Front. Endocrinol. 2013, 4, 59. [Google Scholar] [CrossRef]
- Watford, M. Glutamine and glutamate: Nonessential or essential amino acids? Anim. Nutr. 2015, 1, 119–122. [Google Scholar] [CrossRef]
- Rossignol, E. Genetics and function of neocortical GABAergic interneurons in neurodevelopmental disorders. Neural Plast. 2011, 2011, 649325. [Google Scholar] [CrossRef] [PubMed]
- Luscher, B.; Fuchs, T.; Kilpatrick, C.L. GABAA receptor trafficking-mediated plasticity of inhibitory synapses. Neuron 2011, 70, 385–409. [Google Scholar] [CrossRef] [PubMed]
- Tatti, R.; Haley, M.S.; Swanson, O.K.; Tselha, T.; Maffei, A. Neurophysiology and regulation of the balance between excitation and inhibition in neocortical circuits. Biol. Psychiatry 2017, 81, 821–831. [Google Scholar] [CrossRef]
- Barker, G.R.; Bird, F.; Alexander, V.; Warburton, E.C. Recognition memory for objects, place, and temporal order: A disconnection analysis of the role of the medial prefrontal cortex and perirhinal cortex. J. Neurosci. 2007, 27, 2948–2957. [Google Scholar] [CrossRef]
- Winters, B.D.; Saksida, L.M.; Bussey, T.J. Object recognition memory: Neurobiological mechanisms of encoding, consolidation and retrieval. Neurosci. Biobehav. Rev. 2008, 32, 1055–1070. [Google Scholar] [CrossRef] [PubMed]
- Barker, G.R.; Warburton, E.C. When is the hippocampus involved in recognition memory? J. Neurosci. 2011, 31, 10721–10731. [Google Scholar] [CrossRef]
- Hammond, R.S.; Tull, L.E.; Stackman, R.W. On the delay-dependent involvement of the hippocampus in object recognition memory. Neurobiol. Learn. Mem. 2004, 82, 26–34. [Google Scholar] [CrossRef]
- Clark, R.E.; Zola, S.M.; Squire, L.R. Impaired recognition memory in rats after damage to the hippocampus. J. Neurosci. 2000, 20, 8853–8860. [Google Scholar] [CrossRef]
- Broadbent, N.J.; Gaskin, S.; Squire, L.R.; Clark, R.E. Object recognition memory and the rodent hippocampus. Learn. Mem. 2009, 17, 5–11. [Google Scholar] [CrossRef]
- Featherstone, R.E.; Gifford, R.L.; Crown, L.M.; Amirfathi, F.; Alaniz, J.P.; Yi, J.; Tran, A.; Adomian, D.; Schwenk, A.; Melnychenko, O.; et al. Early life social instability stress causes lasting cognitive decrement and elevated hippocampal stress-related gene expression. Exp. Neurol. 2022, 354, 114099. [Google Scholar] [CrossRef]
- Brunson, K.L.; Kramár, E.; Lin, B.; Chen, Y.; Colgin, L.L.; Yanagihara, T.K.; Lynch, G.; Baram, T.Z. Mechanisms of late-onset cognitive decline after early-life stress. J. Neurosci. 2005, 25, 9328–9338. [Google Scholar] [CrossRef] [PubMed]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- de Azeredo, L.A.; Wearick-Silva, L.E.; Viola, T.W.; Tractenberg, S.G.; Centeno-Silva, A.; Orso, R.; Schröder, N.; Bredy, T.W.; Grassi-Oliveira, R. Maternal separation induces hippocampal changes in cadherin-1 (CDH-1) mRNA and recognition memory impairment in adolescent mice. Neurobiol. Learn. Mem. 2017, 141, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Makena, N.; Bugarith, K.; Russell, V.A. Maternal separation enhances object location memory and prevents exercise-induced MAPK/ERK signalling in adult Sprague-Dawley rats. Metab. Brain Dis. 2012, 27, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Reincke, S.A.; Hanganu-Opatz, I.L. Early-life stress impairs recognition memory and perturbs the functional maturation of prefrontal-hippocampal-perirhinal networks. Sci. Rep. 2017, 7, 42042. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shao, F.; Wang, W. Maternal separation produces alterations of forebrain brain-derived neurotrophic factor expression in differently aged rats. Front. Mol. Neurosci. 2015, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Ding, Y.; Rong, Y.; Geng, Y.; Lai, L.; Qi, D.; Tang, Y.; Yang, L.; Li, J.; Zhou, T.; et al. Early life stress increases brain glutamate and induces neurobehavioral manifestations in rats. ACS Chem. Neurosci. 2020, 11, 4169–4178. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef]
- Nagy, J. Alcohol related changes in regulation of NMDA receptor functions. Curr. Neuropharmacol. 2008, 6, 39–54. [Google Scholar] [CrossRef]
- Filarowska-Jurko, J.; Komsta, L.; Smaga, I.; Surowka, P.; Marszalek-Grabska, M.; Grochecki, P.; Nizio, D.; Filip, M.; Kotlinska, J.H. Maternal separation alters ethanol drinking and reversal learning processes in adolescent rats: The impact of sex and glycine transporter type 1 (GlyT1) inhibitor. Int. J. Mol. Sci. 2022, 23, 5350. [Google Scholar] [CrossRef]
- Matta, J.A.; Ashby, M.C.; Sanz-Clemente, A.; Roche, K.W.; Isaac, J.T. mGluR5 and NMDA receptors drive the experience- and activity-dependent NMDA receptor NR2B to NR2A subunit switch. Neuron 2011, 70, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Liao, P.F.; Chan, M.H. mGluR5 positive modulators both potentiate activation and restore inhibition in NMDA receptors by PKC dependent pathway. J. Biomed. Sci. 2011, 18, 19. [Google Scholar] [CrossRef]
- Marszalek-Grabska, M.; Gibula-Bruzda, E.; Bodzon-Kulakowska, A.; Suder, P.; Gawel, K.; Filarowska, J.; Listos, J.; Danysz, W.; Kotlinska, J.H. Effects of the positive allosteric modulator of metabotropic glutamate receptor 5, VU-29, on impairment of novel object recognition induced by acute ethanol and ethanol withdrawal in rats. Neurotox. Res. 2018, 33, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Lopatynska-Mazurek, M.; Pankowska, A.; Gibula-Tarlowska, E.; Pietura, R.; Kotlinska, J.H. Rapamycin improves recognition memory and normalizes amino-acids and amines levels in the hippocampal dentate gyrus in adult rats exposed to ethanol during the neonatal period. Biomolecules 2021, 11, 362. [Google Scholar] [CrossRef] [PubMed]
- Bevins, R.A.; Besheer, J. Object recognition in rats and mice: A one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nat. Protoc. 2006, 1, 1306–1311. [Google Scholar] [CrossRef]
- Lueptow, L.M. Novel object recognition test for the investigation of learning and memory in mice. J. Vis. Exp. 2017, 126, e55718. [Google Scholar]
- González-Pardo, H.; Arias, J.L.; Vallejo, G.; Conejo, N.M. Influence of environmental enrichment on the volume of brain regions sensitive to early life stress by maternal separation in rats. Psicothema 2019, 31, 46–52. [Google Scholar]
- Hui, J.J.; Zhang, Z.J.; Liu, S.S.; Xi, G.J.; Zhang, X.R.; Teng, G.J.; Chan, K.C.; Wu, E.X.; Nie, B.B.; Shan, B.C.; et al. Hippocampal neurochemistry is involved in the behavioural effects of neonatal maternal separation and their reversal by post-weaning environmental enrichment: A magnetic resonance study. Behav. Brain Res. 2011, 217, 122–127. [Google Scholar] [CrossRef]
- Herpfer, I.; Hezel, H.; Reichardt, W.; Clark, K.; Geiger, J.; Gross, C.M.; Heyer, A.; Neagu, V.; Bhatia, H.; Atas, H.C.; et al. Early life stress differentially modulates distinct forms of brain plasticity in young and adult mice. PLoS ONE 2012, 7, e46004. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kenney, J.W.; Okano, E.; Kadoma, K.; Frankland, P.W.; Kida, S. Functional connectivity of multiple brain regions required for the consolidation of social recognition memory. J. Neurosci. 2017, 37, 4103–4116. [Google Scholar] [CrossRef]
- Bremner, J.D.; Vythilingam, M.; Vermetten, E.; Southwick, S.M.; McGlashan, T.; Nazeer, A.; Khan, S.; Vaccarino, L.V.; Soufer, R.; Garg, P.K.; et al. MRI and PET study of deficits in hippocampal structure and function in women with childhood sexual abuse and posttraumatic stress disorder. Am. J. Psychiatry 2003, 160, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Potter, G.G.; McQuoid, D.R.; Boyd, B.; Turner, R.; MacFall, J.R.; Taylor, W.D. Effects of early life stress on depression, cognitive performance and brain morphology. Psychol. Med. 2017, 47, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Teicher, M.H.; Samson, J.A. Annual Research Review: Enduring neurobiological effects of childhood abuse and neglect. J. Child. Psychol. Psychiatry 2016, 57, 241–266. [Google Scholar] [CrossRef]
- He, J.; Zhao, C.; Liu, W.; Huang, J.; Liang, S.; Chen, L.; Tao, J. Neurochemical changes in the hippocampus and prefrontal cortex associated with electroacupuncture for learning and memory impairment. Int. J. Mol. Med. 2018, 41, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Su, B.J.; Shen, X.N.; Bi, Y.L.; Tan, C.C.; Li, J.Q.; Cao, X.P.; Dong, Q.; Tan, L.; Alzheimer’s Disease Neuroimaging Initiative; et al. Plasma sex hormone-binding globulin predicts neurodegeneration and clinical progression in prodromal Alzheimer’s disease. Aging 2020, 12, 14528–14541. [Google Scholar] [CrossRef] [PubMed]
- Mirescu, C.; Peters, J.D.; Gould, E. Early life experience alters response of adult neurogenesis to stress. Nat. Neurosci. 2004, 7, 841–846. [Google Scholar] [CrossRef]
- Hulshof, H.J.; Novati, A.; Sgoifo, A.; Luiten, P.G.; den Boer, J.A.; Meerlo, P. Maternal separation decreases adult hippocampal cell proliferation and impairs cognitive performance but has little effect on stress sensitivity and anxiety in adult Wistar rats. Behav. Brain Res. 2011, 216, 552–560. [Google Scholar] [CrossRef]
- Suri, D.; Veenit, V.; Sarkar, A.; Thiagarajan, D.; Kumar, A.; Nestler, E.J.; Galande, S.; Vaidya, V.A. Early stress evokes age-dependent biphasic changes in hippocampal neurogenesis, BDNF expression, and cognition. Biol. Psychiatry 2013, 73, 658–666. [Google Scholar] [CrossRef]
- Feng, M.; Sheng, G.; Li, Z.; Wang, J.; Ren, K.; Jin, X.; Jiang, K. Postnatal maternal separation enhances tonic GABA current of cortical layer 5 pyramidal neurons in juvenile rats and promotes genesis of GABAergic neurons in neocortical molecular layer and subventricular zone in adult rats. Behav. Brain Res. 2014, 260, 74–82. [Google Scholar] [CrossRef]
- Naninck, E.F.; Hoeijmakers, L.; Kakava-Georgiadou, N.; Meesters, A.; Lazic, S.E.; Lucassen, P.J.; Korosi, A. Chronic early life stress alters developmental and adult neurogenesis and impairs cognitive function in mice. Hippocampus 2015, 25, 309–328. [Google Scholar] [CrossRef]
- Lajud, N.; Torner, L. Early life stress and hippocampal neurogenesis in the neonate: Sexual dimorphism, long term consequences and possible mediators. Front. Mol. Neurosci. 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Roque, A.; Ochoa-Zarzosa, A.; Torner, L. Maternal separation activates microglial cells and induces an inflammatory response in the hippocampus of male rat pups, independently of hypothalamic and peripheral cytokine levels. Brain Behav. Immun. 2016, 55, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S. Neuroprotective function of high glycolytic activity in astrocytes: Common roles in stroke and neurodegenerative diseases. Int. J. Mol. Sci. 2021, 22, 6568. [Google Scholar] [CrossRef] [PubMed]
- Adrover, E.; Pallarés, M.E.; Baier, C.J.; Monteleone, M.C.; Giuliani, F.A.; Waagepetersen, H.S.; Brocco, M.A.; Cabrera, R.; Sonnewald, U.; Schousboe, A.; et al. Glutamate neurotransmission is affected in prenatally stressed offspring. Neurochem. Int. 2015, 88, 73–87. [Google Scholar] [CrossRef]
- Schulz, K.M.; Pearson, J.N.; Neeley, E.W.; Berger, R.; Leonard, S.; Adams, C.E.; Stevens, K.E. Maternal stress during pregnancy causes sex-specific alterations in offspring memory performance, social interactions, indices of anxiety, and body mass. Physiol. Behav. 2011, 104, 340–347. [Google Scholar] [CrossRef]
- Son, G.H.; Geum, D.; Chung, S.; Kim, E.J.; Jo, J.H.; Kim, C.M.; Lee, K.H.; Kim, H.; Choi, S.; Kim, H.T.; et al. Maternal stress produces learning deficits associated with impairment of NMDA receptor-mediated synaptic plasticity. J. Neurosci. 2006, 26, 3309–3318. [Google Scholar] [CrossRef]
- Yaka, R.; Salomon, S.; Matzner, H.; Weinstock, M. Effect of varied gestational stress on acquisition of spatial memory, hippocampal LTP and synaptic proteins in juvenile male rats. Behav. Brain Res. 2007, 179, 126–132. [Google Scholar] [CrossRef]
- Weinstock, M. Effects of maternal stress on development and behaviour in rat offspring. Stress 2001, 4, 157–167. [Google Scholar] [CrossRef]
- Weinstock, M. The long-term behavioural consequences of prenatal stress. Neurosci. Biobehav. Rev. 2008, 32, 1073–1086. [Google Scholar] [CrossRef]
- Bowman, R.E.; MacLusky, N.J.; Sarmiento, Y.; Frankfurt, M.; Gordon, M.; Luine, V.N. Sexually dimorphic effects of prenatal stress on cognition, hormonal responses, and central neurotransmitters. Endocrinology 2004, 145, 3778–3787. [Google Scholar] [CrossRef]
- Woolley, C.S.; Weiland, N.G.; McEwen, B.S.; Schwartzkroin, P.A. Estradiol increases the sensitivity of hippocampal CA1 pyramidal cells to NMDA receptor-mediated synaptic input: Correlation with dendritic spine density. J. Neurosci. 1997, 17, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, H.; Zhang, J.; Huang, Q.; Shen, Z.; Wu, R. Evaluation of neuron-glia integrity by in vivo proton magnetic resonance spectroscopy: Implications for psychiatric disorders. Neurosci. Biobehav. Rev. 2016, 71, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Moffett, J.R.; Ross, B.; Arun, P.; Madhavarao, C.N.; Namboodiri, A.M. N-Acetylaspartate in the CNS: From neurodiagnostics to neurobiology. Prog. Neurobiol. 2007, 81, 89–131. [Google Scholar] [CrossRef] [PubMed]
- de Rosa, V.; Secondo, A.; Pannaccione, A.; Ciccone, R.; Formisano, L.; Guida, N.; Crispino, R.; Fico, A.; Polishchuk, R.; D’Aniello, A.; et al. D-Aspartate treatment attenuates myelin damage and stimulates myelin repair. EMBO Mol. Med. 2019, 11, e9278. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cheng, Z.; Tang, H.; Jiao, H.; Sun, X.; Cui, Q.; Luo, F.; Pan, H.; Ma, C.; Li, B. Neonatal maternal separation impairs prefrontal cortical myelination and cognitive functions in rats through activation of Wnt signaling. Cereb. Cortex 2017, 27, 2871–2884. [Google Scholar] [CrossRef]
- Barker, G.R.; Warburton, E.C. Object-in-place associative recognition memory depends on glutamate receptor neurotransmission within two defined hippocampal-cortical circuits: A critical role for AMPA and NMDA receptors in the hippocampus, perirhinal, and prefrontal cortices. Cereb. Cortex 2015, 25, 472–481. [Google Scholar] [CrossRef]
- Cohen, S.J.; Munchow, A.H.; Rios, L.M.; Zhang, G.; Asgeirsdóttir, H.N.; Stackman, R.W., Jr. The rodent hippocampus is essential for nonspatial object memory. Curr. Biol. 2013, 23, 1685–1690. [Google Scholar] [CrossRef]
- Otto, T.; Eichenbaum, H. Neuronal activity in the hippocampus during delayed non-match to sample performance in rats: Evidence for hippocampal processing in recognition memory. Hippocampus 1992, 2, 323–334. [Google Scholar] [CrossRef]
- Rutishauser, U.; Schuman, E.M.; Mamelak, A.N. Activity of human hippocampal and amygdala neurons during retrieval of declarative memories. Proc. Natl. Acad. Sci. USA 2008, 105, 329–334. [Google Scholar] [CrossRef]
- Hedges, D.W.; Woon, F.L. Early-life stress and cognitive outcome. Psychopharmacology 2011, 214, 121–130. [Google Scholar] [CrossRef]
- Levine, S. Developmental determinants of sensitivity and resistance to stress. Psychoneuroendocrinology 2005, 30, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Gutman, D.A.; Nemeroff, C.B. Neurobiology of early life stress: Rodent studies. Semin. Clin. Neuropsychiatry 2002, 7, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Warburton, E.C.; Barker, G.R.; Brown, M.W. Investigations into the involvement of NMDA mechanisms in recognition memory. Neuropharmacology 2013, 74, 41–47. [Google Scholar] [CrossRef]
- Bashir, Z.I.; Collingridge, G.L. Synaptic plasticity: Long-term potentiation in the hippocampus. Curr. Opin. Neurobiol. 1992, 2, 328–335. [Google Scholar] [CrossRef]
- Malenka, R.C.; Nicoll, R.A. NMDA-receptor-dependent synaptic plasticity: Multiple forms and mechanisms. Trends Neurosci. 1993, 16, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, N.; Aigner, T.G. The glycine/NMDA receptor antagonist HA-966 impairs visual recognition memory in rhesus monkeys. Brain Res. 1996, 731, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zorumski, C.F.; Mennerick, S.; Izumi, Y. Acute and chronic effects of ethanol on learning-related synaptic plasticity. Alcohol 2014, 48, 1–17. [Google Scholar] [CrossRef]
- Yang, Y.; Ge, W.; Chen, Y.; Zhang, Z.; Shen, W.; Wu, C.; Poo, M.; Duan, S. Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine. Proc. Natl. Acad. Sci. USA 2003, 100, 15194–15199. [Google Scholar] [CrossRef]
- Zhang, Z.; Gong, N.; Wang, W.; Xu, L.; Xu, T.L. Bell-shaped D-serine actions on hippocampal long-term depression and spatial memory retrieval. Cereb. Cortex 2008, 18, 2391–2401. [Google Scholar] [CrossRef]
- Xu, T.L.; Gong, N. Glycine and glycine receptor signaling in hippocampal neurons: Diversity, function and regulation. Prog. Neurobiol. 2010, 91, 349–361. [Google Scholar] [CrossRef]
- Zhang, L.H.; Gong, N.; Fei, D.; Xu, L.; Xu, T.L. Glycine uptake regulates hippocampal network activity via glycine receptor-mediated tonic inhibition. Neuropsychopharmacology 2008, 33, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.W.; Ramsey, A.K.; Walker, J.M.; Serfozo, P.; Olive, M.F.; Schachtman, T.R.; Simonyi, A. Functional interaction of mGlu5 and NMDA receptors in aversive learning in rats. Neurobiol. Learn. Mem. 2011, 95, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Marszalek-Grabska, M.; Gawel, K.; Matosiuk, D.; Gibula-Tarlowska, E.; Listos, J.; Kotlinska, J.H. Effects of the positive allosteric modulator of metabotropic glutamate receptor 5, VU-29, on maintenance association between environmental cues and rewarding properties of ethanol in rats. Biomolecules 2020, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Ayala, J.E.; Chen, Y.; Banko, J.L.; Sheffler, D.J.; Williams, R.; Telk, A.N.; Watson, N.L.; Xiang, Z.; Zhang, Y.; Jones, P.J.; et al. mGluR5 positive allosteric modulators facilitate both hippocampal LTP and LTD and enhance spatial learning. Neuropsychopharmacology 2009, 34, 2057–2071. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Total Distance Traveled (cm) | |
|---|---|---|
| Training (Mean ± SEM) | Testing (Mean ± SEM) | |
| Control NS | 1390 ± 133.5 | 1662 ± 121.0 |
| EtOH NS | 1228 ± 144.1 | 1533 ± 109.1 |
| Control MS | 1652 ± 74.54 | 1760 ± 75.69 |
| EtOH MS | 1410 ± 168.1 | 1786 ± 140.1 |
| VU MS | 1240 ± 55.86 | 1871 ± 190.6 |
| VU + EtOH MS | 1522 ± 165.6 | 1912 + 213.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotlinska, J.H.; Grochecki, P.; Michalak, A.; Pankowska, A.; Kochalska, K.; Suder, P.; Ner-Kluza, J.; Matosiuk, D.; Marszalek-Grabska, M. Neonatal Maternal Separation Induces Sexual Dimorphism in Brain Development: The Influence on Amino Acid Levels and Cognitive Disorders. Biomolecules 2023, 13, 1449. https://doi.org/10.3390/biom13101449
Kotlinska JH, Grochecki P, Michalak A, Pankowska A, Kochalska K, Suder P, Ner-Kluza J, Matosiuk D, Marszalek-Grabska M. Neonatal Maternal Separation Induces Sexual Dimorphism in Brain Development: The Influence on Amino Acid Levels and Cognitive Disorders. Biomolecules. 2023; 13(10):1449. https://doi.org/10.3390/biom13101449
Chicago/Turabian StyleKotlinska, Jolanta H., Pawel Grochecki, Agnieszka Michalak, Anna Pankowska, Katarzyna Kochalska, Piotr Suder, Joanna Ner-Kluza, Dariusz Matosiuk, and Marta Marszalek-Grabska. 2023. "Neonatal Maternal Separation Induces Sexual Dimorphism in Brain Development: The Influence on Amino Acid Levels and Cognitive Disorders" Biomolecules 13, no. 10: 1449. https://doi.org/10.3390/biom13101449
APA StyleKotlinska, J. H., Grochecki, P., Michalak, A., Pankowska, A., Kochalska, K., Suder, P., Ner-Kluza, J., Matosiuk, D., & Marszalek-Grabska, M. (2023). Neonatal Maternal Separation Induces Sexual Dimorphism in Brain Development: The Influence on Amino Acid Levels and Cognitive Disorders. Biomolecules, 13(10), 1449. https://doi.org/10.3390/biom13101449