
Immune Responses Induced at One Hour Post Cataract Surgery Wounding of the Chick Lens




Abstract
:1. Introduction
2. Materials and Methods
3. Results
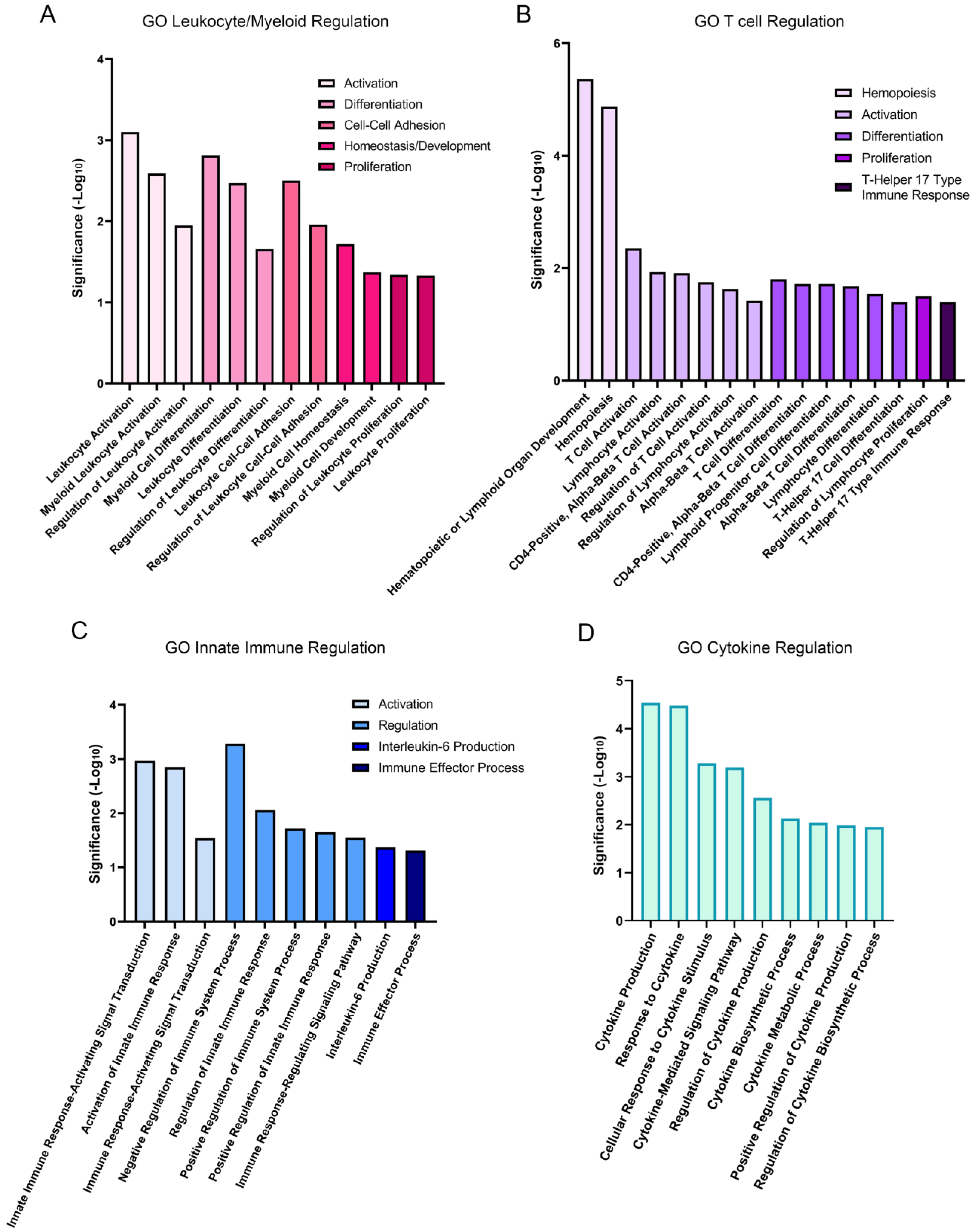
3.1. RNA-Seq Analysis Reveals That Lens Wounding Rapidly Induces an Extensive Immune Response
3.2. Resident Immune Cells of the Lens Are Rapidly Activated in Response to Wounding
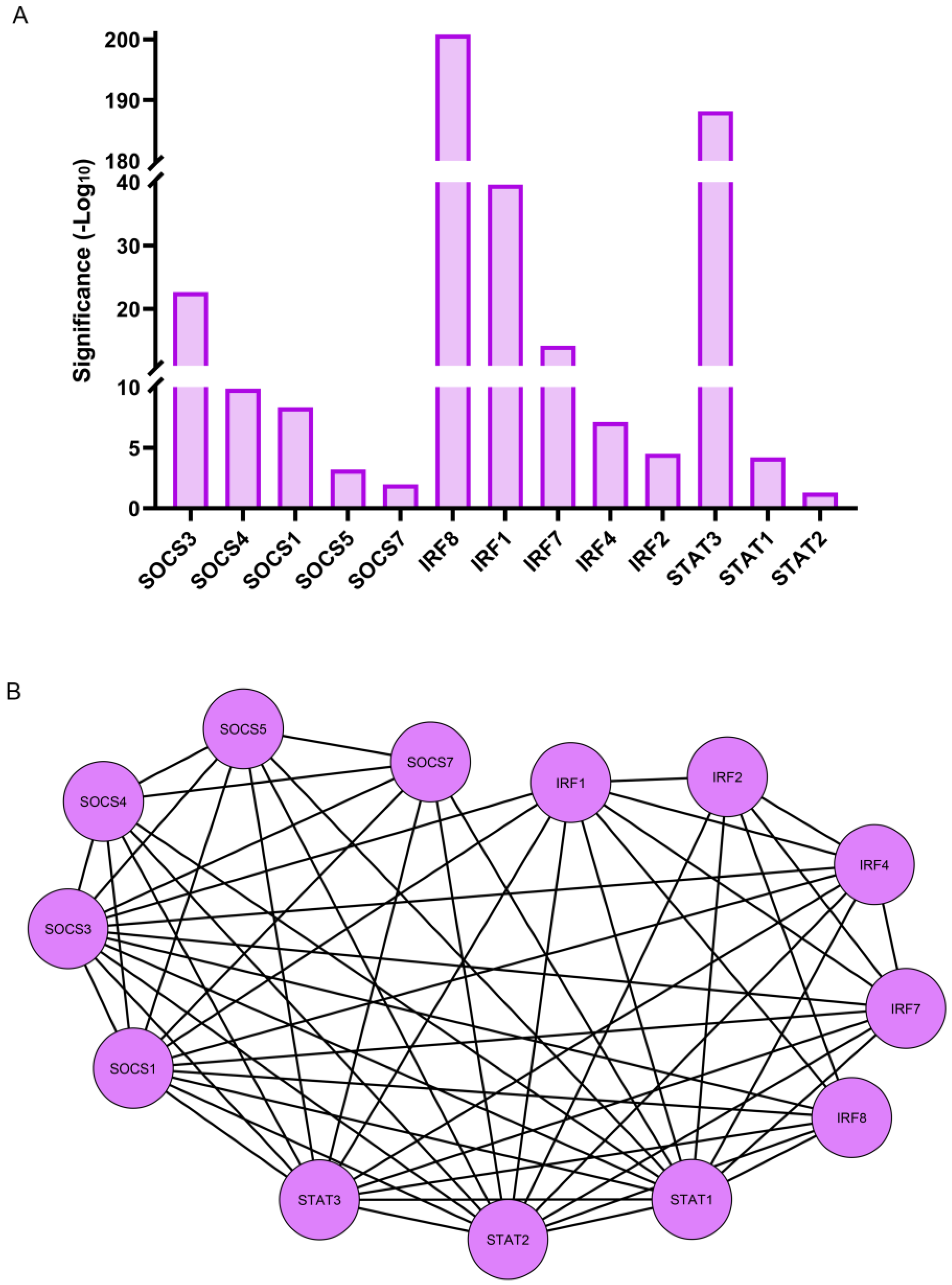
3.3. RNA-Seq Analysis Shows That Transcription Factors Linked to the Immune Response Are Early Responders to Lens Wounding
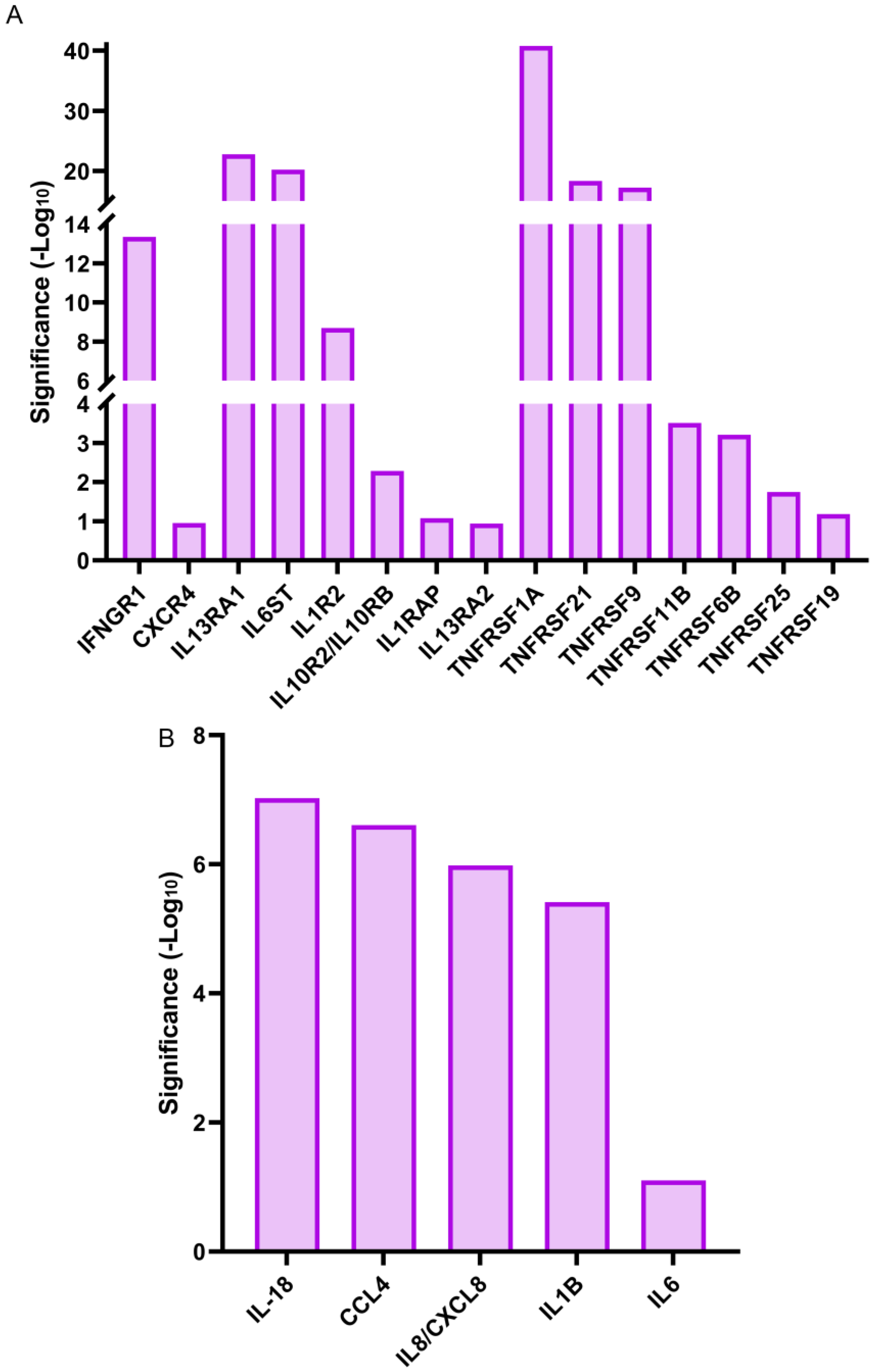
3.4. Gene Expression of a Specific Subset of Cytokines and Cytokine Receptors Were Induced within 1 h of Lens Injury
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tholozan, F.M.; Quinlan, R.A. Lens cells: More than meets the eye. Int. J. Biochem. Cell Biol. 2007, 39, 1754–1759. [Google Scholar] [CrossRef] [PubMed]
- Molzer, C.; Heissigerova, J.; Wilson, H.M.; Kuffova, L.; Forrester, J.V. Immune Privilege: The Microbiome and Uveitis. Front. Immunol. 2020, 11, 608377. [Google Scholar] [CrossRef]
- Taylor, A.W. Ocular immune privilege. Eye 2009, 23, 1885–1889. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.M.; Bowen, C.J.; Menko, A.S. Induction of Immune Surveillance of the Dysmorphogenic Lens. Sci. Rep. 2017, 7, 16235. [Google Scholar] [CrossRef] [PubMed]
- Menko, A.S.; DeDreu, J.; Logan, C.M.; Paulson, H.; Levin, A.V.; Walker, J.L. Resident immune cells of the avascular lens: Mediators of the injury and fibrotic response of the lens. FASEB J. 2021, 35, e21341. [Google Scholar] [CrossRef]
- DeDreu, J.; Pal-Ghosh, S.; Mattapallil, M.J.; Caspi, R.R.; Stepp, M.A.; Menko, A.S. Uveitis-mediated immune cell invasion through the extracellular matrix of the lens capsule. FASEB J. 2022, 36, e21995. [Google Scholar] [CrossRef]
- DeDreu, J.; Bowen, C.J.; Logan, C.M.; Pal-Ghosh, S.; Parlanti, P.; Stepp, M.A.; Menko, A.S. An immune response to the avascular lens following wounding of the cornea involves ciliary zonule fibrils. FASEB J. 2020, 34, 9316–9336. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Quan, Y.; Cheng, H.; Riquelme, M.A.; Li, X.D.; Gu, S.; Jiang, J.X. Macrophage recruitment in immune-privileged lens during capsule repair, necrotic fiber removal, and fibrosis. iScience 2021, 24, 102533. [Google Scholar] [CrossRef]
- Jiang, J.; Shihan, M.H.; Wang, Y.; Duncan, M.K. Lens Epithelial Cells Initiate an Inflammatory Response Following Cataract Surgery. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4986–4997. [Google Scholar] [CrossRef]
- Zhou, Y.; Bennett, T.M.; Shiels, A. Mutation of the TRPM3 cation channel underlies progressive cataract development and lens calcification associated with pro-fibrotic and immune cell responses. FASEB J. 2021, 35, e21288. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Brown, M.A. Innate immunity in the central nervous system. J. Clin. Investig. 2012, 122, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Kivisakk, P.; Kidd, G. Three or more routes for leukocyte migration into the central nervous system. Nat. Rev. Immunol. 2003, 3, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Engelhardt, B. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat. Rev. Immunol. 2012, 12, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Shechter, R.; London, A.; Schwartz, M. Orchestrated leukocyte recruitment to immune-privileged sites: Absolute barriers versus educational gates. Nat. Rev. Immunol. 2013, 13, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Caputa, G.; Castoldi, A.; Pearce, E.J. Metabolic adaptations of tissue-resident immune cells. Nat. Immunol. 2019, 20, 793–801. [Google Scholar] [CrossRef]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-resident macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Tatsumi, N.; Kumamoto, Y. Role of mouse dendritic cell subsets in priming naive CD4 T cells. Curr. Opin. Immunol. 2023, 83, 102352. [Google Scholar] [CrossRef]
- Ginhoux, F.; Guilliams, M. Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity 2016, 44, 439–449. [Google Scholar] [CrossRef]
- Masopust, D.; Soerens, A.G. Tissue-Resident T Cells and Other Resident Leukocytes. Annu. Rev. Immunol. 2019, 37, 521–546. [Google Scholar] [CrossRef]
- Corker, A.; Neff, L.S.; Broughton, P.; Bradshaw, A.D.; DeLeon-Pennell, K.Y. Organized Chaos: Deciphering Immune Cell Heterogeneity’s Role in Inflammation in the Heart. Biomolecules 2021, 12, 11. [Google Scholar] [CrossRef]
- Belz, G.T.; Denman, R.; Seillet, C.; Jacquelot, N. Tissue-resident lymphocytes: Weaponized sentinels at barrier surfaces. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Krueger, P.D.; Kim, T.S.; Sung, S.S.; Braciale, T.J.; Hahn, Y.S. Liver-resident CD103+ dendritic cells prime antiviral CD8+ T cells in situ. J. Immunol. 2015, 194, 3213–3222. [Google Scholar] [CrossRef] [PubMed]
- DeDreu, J.; Le, P.M.; Menko, A.S. The ciliary zonules provide a pathway for immune cells to populate the avascular lens during eye development. Exp. Biol. Med. 2022, 247, 2251–2273. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhao, G.L.; Cheng, S.; Wang, Z.; Yang, X.L. Activation of retinal glial cells contributes to the degeneration of ganglion cells in experimental glaucoma. Prog. Retin. Eye Res. 2023, 93, 101169. [Google Scholar] [CrossRef]
- Yu, F.X.; Lee, P.S.Y.; Yang, L.; Gao, N.; Zhang, Y.; Ljubimov, A.V.; Yang, E.; Zhou, Q.; Xie, L. The impact of sensory neuropathy and inflammation on epithelial wound healing in diabetic corneas. Prog. Retin. Eye Res. 2022, 89, 101039. [Google Scholar] [CrossRef]
- Cao, C.; Yao, Y.; Zeng, R. Lymphocytes: Versatile Participants in Acute Kidney Injury and Progression to Chronic Kidney Disease. Front. Physiol. 2021, 12, 729084. [Google Scholar] [CrossRef]
- Novo, S.G.; Faranda, A.P.; Shihan, M.H.; Wang, Y.; Garg, A.; Duncan, M.K. The Immediate Early Response of Lens Epithelial Cells to Lens Injury. Cells 2022, 11, 3456. [Google Scholar] [CrossRef]
- Rashid, K.; Wolf, A.; Langmann, T. Microglia Activation and Immunomodulatory Therapies for Retinal Degenerations. Front. Cell. Neurosci. 2018, 12, 176. [Google Scholar] [CrossRef]
- Polak, M.E.; Thirdborough, S.M.; Ung, C.Y.; Elliott, T.; Healy, E.; Freeman, T.C.; Ardern-Jones, M.R. Distinct molecular signature of human skin Langerhans cells denotes critical differences in cutaneous dendritic cell immune regulation. J. Investig. Dermatol. 2014, 134, 695–703. [Google Scholar] [CrossRef]
- Miller, J.C.; Brown, B.D.; Shay, T.; Gautier, E.L.; Jojic, V.; Cohain, A.; Pandey, G.; Leboeuf, M.; Elpek, K.G.; Helft, J.; et al. Deciphering the transcriptional network of the dendritic cell lineage. Nat. Immunol. 2012, 13, 888–899. [Google Scholar] [CrossRef]
- Clemen, R.; Arlt, K.; Miebach, L.; von Woedtke, T.; Bekeschus, S. Oxidized Proteins Differentially Affect Maturation and Activation of Human Monocyte-Derived Cells. Cells 2022, 11, 3659. [Google Scholar] [CrossRef] [PubMed]
- Menko, A.S.; Bleaken, B.M.; Libowitz, A.A.; Zhang, L.; Stepp, M.A.; Walker, J.L. A central role for vimentin in regulating repair function during healing of the lens epithelium. Mol. Biol. Cell 2014, 25, 776–790. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.L.; Bleaken, B.M.; Wolff, I.M.; Menko, A.S. Establishment of a Clinically Relevant Ex Vivo Mock Cataract Surgery Model for Investigating Epithelial Wound Repair in a Native Microenvironment. J. Vis. Exp. 2015, 100, e52886. [Google Scholar] [CrossRef]
- Bleaken, B.M.; Menko, A.S.; Walker, J.L. Cells activated for wound repair have the potential to direct collective invasion of an epithelium. Mol. Biol. Cell 2016, 27, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.L.; Bleaken, B.M.; Romisher, A.R.; Alnwibit, A.A.; Menko, A.S. In wound repair vimentin mediates the transition of mesenchymal leader cells to a myofibroblast phenotype. Mol. Biol. Cell 2018, 29, 1555–1570. [Google Scholar] [CrossRef] [PubMed]
- Vial, C.; Zúñiga, L.M.; Cabello-Verrugio, C.; Cañón, P.; Fadic, R.; Brandan, E. Skeletal muscle cells express the profibrotic cytokine connective tissue growth factor (CTGF/CCN2), which induces their dedifferentiation. J. Cell. Physiol. 2008, 215, 410–421. [Google Scholar] [CrossRef]
- Xia, X.; Wang, W.; Yin, K.; Wang, S. Interferon regulatory factor 8 governs myeloid cell development. Cytokine Growth Factor. Rev. 2020, 55, 48–57. [Google Scholar] [CrossRef]
- Cacalano, N.A. Regulation of Natural Killer Cell Function by STAT3. Front. Immunol. 2016, 7, 128. [Google Scholar] [CrossRef]
- Feinberg, M.W.; Shimizu, K.; Lebedeva, M.; Haspel, R.; Takayama, K.; Chen, Z.; Frederick, J.P.; Wang, X.F.; Simon, D.I.; Libby, P.; et al. Essential role for Smad3 in regulating MCP-1 expression and vascular inflammation. Circ. Res. 2004, 94, 601–608. [Google Scholar] [CrossRef]
- Kwon, S. Molecular dissection of Janus kinases as drug targets for inflammatory diseases. Front. Immunol. 2022, 13, 1075192. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.L.; Menko, A.S. Immune cells in lens injury repair and fibrosis. Exp. Eye Res. 2021, 209, 108664. [Google Scholar] [CrossRef] [PubMed]
- Bykovskaia, S.N.; Shurin, G.V.; Graner, S.; Bunker, M.L.; Olson, W.; Thomas, R.; Shurin, M.R.; Marks, S.; Storkus, W.J.; Shogan, J. Differentiation of immunostimulatory stem-cell- and monocyte-derived dendritic cells involves maturation of intracellular compartments responsible for antigen presentation and secretion. Stem Cells 2002, 20, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Jackson, D.G. Hyaluronan and Its Receptors: Key Mediators of Immune Cell Entry and Trafficking in the Lymphatic System. Cells 2021, 10, 2061. [Google Scholar] [CrossRef] [PubMed]
- Sheen, J.H.; Strainic, M.G.; Liu, J.; Zhang, W.; Yi, Z.; Medof, M.E.; Heeger, P.S. TLR-Induced Murine Dendritic Cell (DC) Activation Requires DC-Intrinsic Complement. J. Immunol. 2017, 199, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Dulyaninova, N.G.; House, R.P.; Almo, S.C.; Bresnick, A.R. S100A4 regulates macrophage chemotaxis. Mol. Biol. Cell 2010, 21, 2598–2610. [Google Scholar] [CrossRef]
- Peters, V.A.; Joesting, J.J.; Freund, G.G. IL-1 receptor 2 (IL-1R2) and its role in immune regulation. Brain Behav. Immun. 2013, 32, 1–8. [Google Scholar] [CrossRef]
- Apaydin, D.C.; Zakarauskas-Seth, B.I.; Carnevale, L.; Apaydin, O.; Perrotta, M.; Carnevale, R.; Kotini, M.P.; Kotlar-Goldaper, I.; Belting, H.G.; Carnevale, D.; et al. Interferon-γ drives macrophage reprogramming, cerebrovascular remodelling, and cognitive dysfunction in a zebrafish and a mouse model of ion imbalance and pressure overload. Cardiovasc. Res. 2023, 119, 1234–1249. [Google Scholar] [CrossRef]
- Li, W.; Li, Y.; Jin, X.; Liao, Q.; Chen, Z.; Peng, H.; Zhou, Y. CD38: A Significant Regulator of Macrophage Function. Front. Oncol. 2022, 12, 775649. [Google Scholar] [CrossRef]
- Piedra-Quintero, Z.L.; Wilson, Z.; Nava, P.; Guerau-de-Arellano, M. CD38: An Immunomodulatory Molecule in Inflammation and Autoimmunity. Front. Immunol. 2020, 11, 597959. [Google Scholar] [CrossRef]
- Desai, V.D.; Wang, Y.; Simirskii, V.N.; Duncan, M.K. CD44 expression is developmentally regulated in the mouse lens and increases in the lens epithelium after injury. Differentiation 2010, 79, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Maddala, R.; Gao, J.; Mathias, R.T.; Lewis, T.R.; Arshavsky, V.Y.; Levine, A.; Backer, J.M.; Bresnick, A.R.; Rao, P.V. Absence of S100A4 in the mouse lens induces an aberrant retina-specific differentiation program and cataract. Sci. Rep. 2021, 11, 2203. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, K.; Cheng, A.; Wang, M.; Cui, M.; Huang, J.; Zhu, D.; Chen, S.; Liu, M.; Zhao, X.; et al. SOCS Proteins Participate in the Regulation of Innate Immune Response Caused by Viruses. Front. Immunol. 2020, 11, 558341. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.M.; Heller, N.M. Regulation of Macrophage, Dendritic Cell, and Microglial Phenotype and Function by the SOCS Proteins. Front. Immunol. 2015, 6, 549. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Tanaka, N.; Taki, S. Regulation of the interferon system, immune response and oncogenesis by the transcription factor interferon regulatory factor-1. Eur. Cytokine Netw. 1998, 9, 43–48. [Google Scholar]
- Pandit, H.; Valentin, A.; Angel, M.; Deleage, C.; Bergamaschi, C.; Bear, J.; Sowder, R.; Felber, B.K.; Pavlakis, G.N. Step-dose IL-7 treatment promotes systemic expansion of T cells and alters immune cell landscape in blood and lymph nodes. iScience 2023, 26, 105929. [Google Scholar] [CrossRef]
- Seif, F.; Khoshmirsafa, M.; Aazami, H.; Mohsenzadegan, M.; Sedighi, G.; Bahar, M. The role of JAK-STAT signaling pathway and its regulators in the fate of T helper cells. Cell Commun. Signal 2017, 15, 23. [Google Scholar] [CrossRef]
- Hu, Q.; Bian, Q.; Rong, D.; Wang, L.; Song, J.; Huang, H.S.; Zeng, J.; Mei, J.; Wang, P.Y. JAK/STAT pathway: Extracellular signals, diseases, immunity, and therapeutic regimens. Front. Bioeng. Biotechnol. 2023, 11, 1110765. [Google Scholar] [CrossRef]
- Arend, W.P.; Palmer, G.; Gabay, C. IL-1, IL-18, and IL-33 families of cytokines. Immunol. Rev. 2008, 223, 20–38. [Google Scholar] [CrossRef]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef]
- Somm, E.; Jornayvaz, F.R. Interleukin-18 in metabolism: From mice physiology to human diseases. Front. Endocrinol. 2022, 13, 971745. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ju, X.; Silveira, P.A.; Abadir, E.; Hsu, W.H.; Hart, D.N.J.; Clark, G.J. CD83: Activation Marker for Antigen Presenting Cells and Its Therapeutic Potential. Front. Immunol. 2019, 10, 1312. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.M.; Noon-Song, E.N.; Dabelic, R.; Ahmed, C.M. IFN signaling: How a non-canonical model led to the development of IFN mimetics. Front. Immunol. 2013, 4, 202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, W.; Zhang, J.; Li, T.; Ma, Y.; Wang, Y.; Li, X.; Cui, W.; Du, J. CXCL16 Promotes Ly6Chigh Monocyte Infiltration and Impairs Heart Function after Acute Myocardial Infarction. J. Immunol. 2023, 210, 820–831. [Google Scholar] [CrossRef]
- Lukhele, S.; Rabbo, D.A.; Guo, M.; Shen, J.; Elsaesser, H.J.; Quevedo, R.; Carew, M.; Gadalla, R.; Snell, L.M.; Mahesh, L.; et al. The transcription factor IRF2 drives interferon-mediated CD8(+) T cell exhaustion to restrict anti-tumor immunity. Immunity 2022, 55, 2369–2385.e2310. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Banerjee, S.; Guo, S.; Xie, N.; Liu, G. IFN Regulatory Factor 2 Inhibits Expression of Glycolytic Genes and Lipopolysaccharide-Induced Proinflammatory Responses in Macrophages. J. Immunol. 2018, 200, 3218–3230. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEGs | Significance (-Log10) | DEGs | Significance (-Log10) | ||
|---|---|---|---|---|---|
| 1 | CTGF | * | 26 | SNRK | 187.5951835 |
| 2 | JUN | * | 27 | CBX4 | 185.0514131 |
| 3 | BTG2 | * | 28 | BHLHE40 | 181.4359902 |
| 4 | GADD45B | * | 29 | ANGPTL4 | 176.3661648 |
| 5 | CISH | * | 30 | LONRF3 | 166.8524546 |
| 6 | NFIL3 | * | 31 | NET1 | 166.375484 |
| 7 | MYC | * | 32 | MSX1 | 162.4118963 |
| 8 | CREM | * | 33 | EIF2AK3 | 160.8609588 |
| 9 | F3 | 288.1154735 | 34 | MAFK | 157.9399122 |
| 10 | KLF6 | 286.5836756 | 35 | VCL | 156.3186263 |
| 11 | TGIF1 | 270.8078907 | 36 | EPHA2 | 154.7870455 |
| 12 | FOSL2 | 268.5125469 | 37 | EDN2 | 152.0153907 |
| 13 | PCDH9 | 263.8520327 | 38 | SRF | 147.5372354 |
| 14 | ERRFI1 | 263.0684018 | 39 | PLK3 | 143.6585299 |
| 15 | ARID5A | 241.9775771 | 40 | POLE | 138.7371147 |
| 16 | DUSP8 | 231.57357 | 41 | MAP3K14 | 133.8617124 |
| 17 | MAFF | 215.6329808 | 42 | IER5 | 127.8269684 |
| 18 | ATF3 | 212.2826148 | 43 | BACH1 | 124.3708318 |
| 19 | MAPKKK3L | 212.1738763 | 44 | ZFP36L1 | 123.5899623 |
| 20 | PIM1 | 203.6009763 | 45 | SERTAD2 | 120.4135481 |
| 21 | LONRF1 | 200.8374542 | 46 | ITPKB | 117.3457473 |
| 22 | IRF8 | 200.8362272 | 47 | KLF11 | 116.8070514 |
| 23 | BTG1 | 197.4431125 | 48 | THBS1 | 115.5955296 |
| 24 | PIM3 | 197.4431125 | 49 | PISD | 114.842178 |
| 25 | STAT3 | 188.1798597 | 50 | SGMS2 | 111.7124197 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeDreu, J.; Basta, M.D.; Walker, J.L.; Menko, A.S. Immune Responses Induced at One Hour Post Cataract Surgery Wounding of the Chick Lens. Biomolecules 2023, 13, 1615. https://doi.org/10.3390/biom13111615
DeDreu J, Basta MD, Walker JL, Menko AS. Immune Responses Induced at One Hour Post Cataract Surgery Wounding of the Chick Lens. Biomolecules. 2023; 13(11):1615. https://doi.org/10.3390/biom13111615
Chicago/Turabian StyleDeDreu, JodiRae, Morgan D. Basta, Janice L. Walker, and A. Sue Menko. 2023. "Immune Responses Induced at One Hour Post Cataract Surgery Wounding of the Chick Lens" Biomolecules 13, no. 11: 1615. https://doi.org/10.3390/biom13111615
APA StyleDeDreu, J., Basta, M. D., Walker, J. L., & Menko, A. S. (2023). Immune Responses Induced at One Hour Post Cataract Surgery Wounding of the Chick Lens. Biomolecules, 13(11), 1615. https://doi.org/10.3390/biom13111615