
The Effects of Unilateral Labyrinthectomy on Monoamine Neurotransmitters in the Medial Vestibular Nucleus of Rats
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Designs
2.3. Unilateral Labyrinthectomy (UL)
2.4. Immunofluorescence
2.5. Sample Preparation and Ultra-High-Performance Liquid Chromatography
2.6. Behavioral Assessment
2.7. Statistical Analysis
3. Results
3.1. Qualitative Evaluation of the Acute Vestibular Syndrome
3.2. The Excitation Marked by c-Fos Induction Is Enhanced in the Ipsilesional MVN after Unilateral Labyrinthectomy
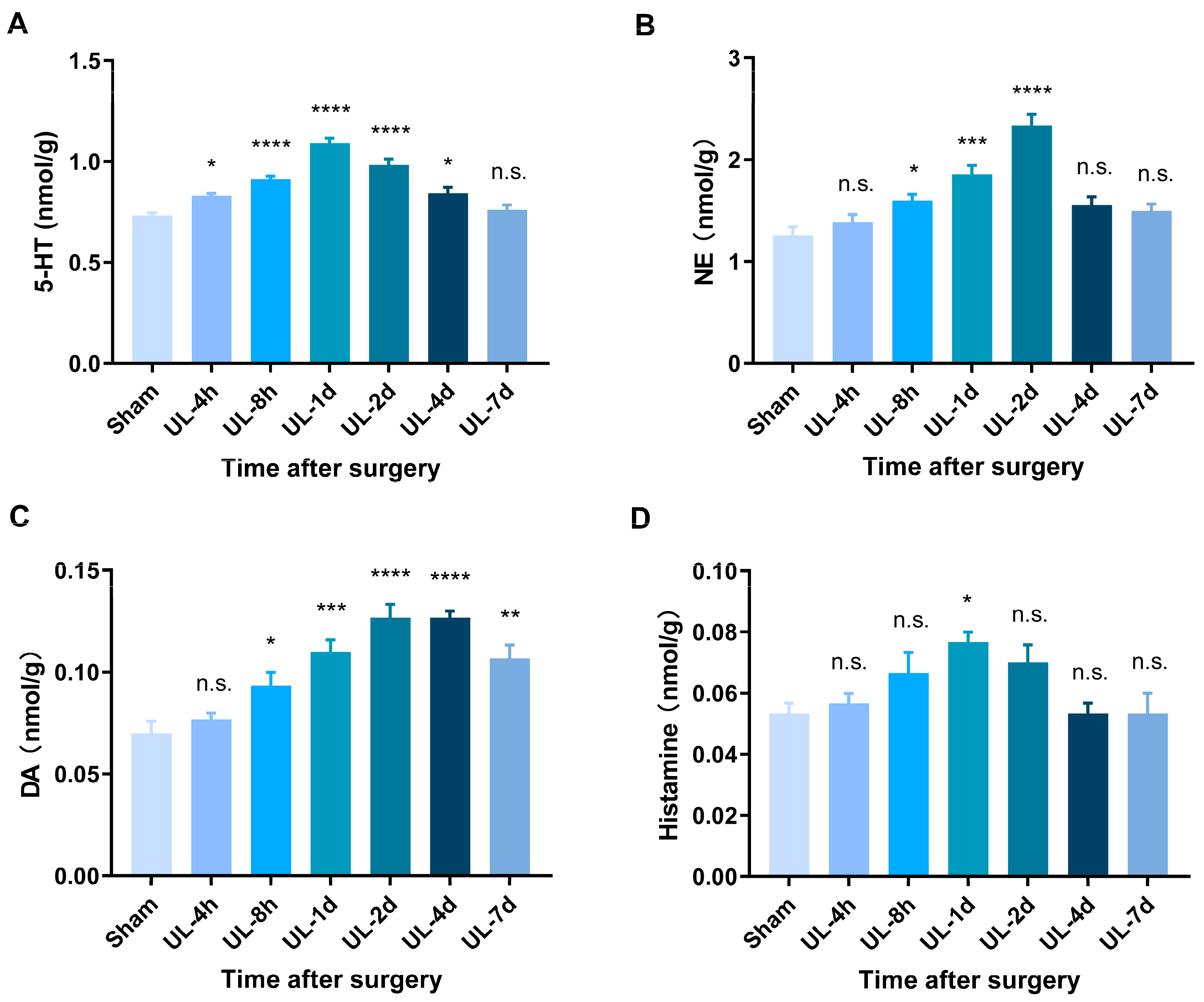
3.3. HPLC Analysis Reveals Elevated Levels of Monoamine Neurotransmitters during Vestibular Compensation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fetter, M. Acute unilateral loss of vestibular function. Handb. Clin. Neurol. 2016, 137, 219–229. [Google Scholar] [PubMed]
- Tighilet, B.; Chabbert, C. Adult neurogenesis promotes balance recovery after vestibular loss. Prog. Neurobiol. 2019, 174, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Strupp, M.; Mandalà, M.; López-Escámez, J.A. Peripheral vestibular disorders: An update. Curr. Opin. Neurol. 2019, 32, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Kammerlind, A.-S.C.; Ledin, T.E.; Ödkvist, L.M.; Skargren, E.I. Recovery after acute unilateral vestibular loss and predictors for remaining symptoms. Am. J. Otolaryngol. 2011, 32, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Lacour, M.; Dutheil, S.; Tighilet, B.; Lopez, C.; Borel, L. Tell me your vestibular deficit, and i’ll tell you how you’ll compensate. Ann. N. Y. Acad. Sci. 2009, 1164, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Dutia, M.B. Mechanisms of vestibular compensation: Recent advances. Curr. Opin. Otolaryngol. Head Neck Surg. 2010, 18, 420–424. [Google Scholar] [CrossRef]
- Grosch, M.; Lindner, M.; Bartenstein, P.; Brandt, T.; Dieterich, M.; Ziegler, S.; Zwergal, A. Dynamic whole-brain metabolic connectivity during vestibular compensation in the rat. Neuroimage 2021, 226, 117588. [Google Scholar] [CrossRef]
- Ris, L.; Capron, B.; de Waele, C.; Vidal, P.P.; Godaux, E. Dissociations between behavioural recovery and restoration of vestibular activity in the unilabyrinthectomized guinea-pig. J. Physiol. 1997, 500 Pt 2, 509–522. [Google Scholar] [CrossRef]
- Han, L.; Wu, K.L.-K.; Kwan, P.-Y.; Chua, O.W.-H.; Shum, D.K.-Y.; Chan, Y.-S. 5-HT1A receptor-mediated attenuation of synaptic transmission in rat medial vestibular nucleus impacts on vestibular-related motor function. J. Physiol. 2021, 599, 253–267. [Google Scholar] [CrossRef]
- Zhang, K.; Li, Q.; Xu, J.; Liu, J.; Ke, J.; Kang, W.; Li, T.; Ma, F. Unilateral horizontal semicircular canal occlusion induces serotonin increase in medial vestibular nuclei: A study using microdialysis in vivo coupled with HPLC-ECD. Analyst 2015, 140, 3846–3851. [Google Scholar] [CrossRef]
- Onodera, K.; Itoh, C.; Sato, M.; Watanabe, T. Motor behavioural function for histamine-dopamine interaction in brain. Inflamm. Res. 1998, 47 (Suppl. S1), S30–S31. [Google Scholar] [CrossRef] [PubMed]
- Ruhé, H.G.; Mason, N.S.; Schene, A.H. Mood is indirectly related to serotonin, norepinephrine and dopamine levels in humans: A meta-analysis of monoamine depletion studies. Mol. Psychiatry 2007, 12, 331–359. [Google Scholar] [CrossRef] [PubMed]
- Bacqué-Cazenave, J.; Bharatiya, R.; Barrière, G.; Delbecque, J.-P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdère, P. Serotonin in Animal Cognition and Behavior. Int. J. Mol. Sci. 2020, 21, 1649. [Google Scholar] [CrossRef] [PubMed]
- Balaban, C.D. Neurotransmitters in the vestibular system. Handb. Clin. Neurol. 2016, 137, 41–55. [Google Scholar] [PubMed]
- Eberhard, K.E.; Kirkeby, S.; Hansen, L.J.; Cayé-Thomasen, P. Neurotransmitter and neurotransmitter receptor expression in the saccule of the human vestibular system. Prog. Neurobiol. 2022, 212, 102238. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, J.; Zhou, L.; Tian, E.; Chen, J.; Kong, W.; Lu, Y.; Zhang, S. Differential Modulation of Cerebellar Flocculus Unipolar Brush Cells during Vestibular Compensation. Biomedicines 2023, 11, 1298. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, W.; Zhang, S.; Liu, B.; Liang, P.; Zhou, Y.; Zhou, T.; Zhang, K.; Leng, Y.; Kong, W. BDNF signaling in the rat cerebello-vestibular pathway during vestibular compensation: BDNF signaling in vestibular compensation. FEBS J. 2015, 282, 3579–3591. [Google Scholar] [CrossRef]
- Chen, Z.-P.; Zhang, X.-Y.; Peng, S.-Y.; Yang, Z.-Q.; Wang, Y.-B.; Zhang, Y.-X.; Chen, X.; Wang, J.-J.; Zhu, J.-N. Histamine H1 Receptor Contributes to Vestibular Compensation. J. Neurosci. 2019, 39, 420–433. [Google Scholar] [CrossRef]
- Cransac, H.; Peyrin, L.; Farhat, F.; Cottet-Emard, J.M.; Pequignot, J.M.; Reber, A. Effect of hemilabyrinthectomy on monoamine metabolism in the medial vestibular nucleus, locus coeruleus, and other brainstem nuclei of albino and pigmented rats. J. Vestib. Res. 1996, 6, 243–253. [Google Scholar] [CrossRef]
- Khalmuratova, R.; Hah, Y.S.; Ahn, S.K.; Jeon, S.Y.; Kim, J.P.; Balaban, C.D. Immunohistochemical and biomolecular identification of 5-HT7 receptor in rat vestibular nuclei. J. Vestib. Res. 2010, 20, 401–406. [Google Scholar] [CrossRef]
- Pietkiewicz, P.; Pepaś, R.; Sułkowski, W.J.; Zielińska-Bliźniewska, H.; Olszewski, J. Electronystagmography versus videonystagmography in diagnosis of vertigo. Int. J. Occup. Med. Environ. Health 2012, 25, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Günther, L.; Beck, R.; Xiong, G.; Potschka, H.; Jahn, K.; Bartenstein, P.; Brandt, T.; Dutia, M.; Dieterich, M.; Strupp, M.; et al. N-acetyl-L-leucine accelerates vestibular compensation after unilateral labyrinthectomy by action in the cerebellum and thalamus. PLoS ONE 2015, 10, e0120891. [Google Scholar] [CrossRef] [PubMed]
- Esenaliev, R.O.; Petrov, I.Y.; Petrov, Y.; Guptarak, J.; Boone, D.R.; Mocciaro, E.; Weisz, H.; Parsley, M.A.; Sell, S.L.; Hellmich, H.; et al. Nano-Pulsed Laser Therapy Is Neuroprotective in a Rat Model of Blast-Induced Neurotrauma. J. Neurotrauma 2018, 35, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Herbin, M.; Simonis, C.; Revéret, L.; Hackert, R.; Libourel, P.-A.; Eugène, D.; Diaz, J.; de Waele, C.; Vidal, P.-P. Dopamine Modulates Motor Control in a Specific Plane Related to Support. PLoS ONE 2016, 11, e0155058. [Google Scholar] [CrossRef] [PubMed]
- Devèze, A.; Montava, M.; Lopez, C.; Lacour, M.; Magnan, J.; Borel, L. Vestibular compensation following vestibular neurotomy. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2015, 132, 197–203. [Google Scholar] [CrossRef]
- Jamali, M.; Mitchell, D.E.; Dale, A.; Carriot, J.; Sadeghi, S.G.; Cullen, K.E. Neuronal detection thresholds during vestibular compensation: Contributions of response variability and sensory substitution. J. Physiol. 2014, 592, 1565–1580. [Google Scholar] [CrossRef]
- Kitahara, T.; Kaneko, T.; Horii, A.; Fukushima, M.; Kizawa-Okumura, K.; Takeda, N.; Kubo, T. Fos-enkephalin signaling in the rat medial vestibular nucleus facilitates vestibular compensation. J. Neurosci. Res. 2006, 83, 1573–1583. [Google Scholar] [CrossRef]
- D’Ascanio, P.; Arrighi, P.; Pompeiano, O. Fos-protein expression in noradrenergic locus coeruleus neurons after unilateral labyrinthectomy in the rat. Arch. Ital. Biol. 1998, 136, 81–102. [Google Scholar]
- Tighilet, B.; Bordiga, P.; Cassel, R.; Chabbert, C. Peripheral vestibular plasticity vs central compensation: Evidence and questions. J. Neurol. 2019, 266 (Suppl. S1), 27–32. [Google Scholar] [CrossRef]
- Zhai, F.; Shi, F.; Wang, J.; Dai, C.-F.; Fan, C. Preliminary study on the mechanism underlying the interaction of balance dysfunction and anxiety disorder. Neuroreport 2019, 30, 53–59. [Google Scholar] [CrossRef]
- Hoshino, K.; Pompeiano, O.; Magherini, P.C.; Mergner, T. The oscillatory system responsible for the oculomotor activity during the bursts of REM. Arch. Ital. Biol. 1976, 114, 278–309. [Google Scholar]
- Ternaux, J.P.; Gambarelli, F. Modulation of the vestibulo-ocular reflex by serotonin in the rat. Pflugers Arch. 1987, 409, 507–511. [Google Scholar] [CrossRef] [PubMed]
- D’Ascanio, P.; Arrighi, P.; Fascetti, F.; Pompeiano, O. Effects of unilateral labyrinthectomy on the norepinephrine content in forebrain and cerebellar structures of albino rats. Arch. Ital. Biol. 2000, 138, 241–270. [Google Scholar] [PubMed]
- Pompeiano, O. Noradrenergic influences on the cerebellar cortex: Effects on vestibular reflexes under basic and adaptive conditions. Otolaryngol. Head Neck Surg. 1998, 119, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Drago, F.; Nardo, L.; Rampello, L.; Raffaele, R. Vestibular compensation in aged rats with unilateral labyrinthectomy treated with dopaminergic drugs. Pharmacol. Res. 1996, 33, 135–140. [Google Scholar] [CrossRef]
- Tighilet, B.; Lacour, M. Histamine immunoreactivity changes in vestibular-lesioned and histaminergic-treated cats. Eur. J. Pharmacol. 1997, 330, 65–77. [Google Scholar] [CrossRef]
- Tighilet, B.; Mourre, C.; Trottier, S.; Lacour, M. Histaminergic ligands improve vestibular compensation in the cat: Behavioural, neurochemical and molecular evidence. Eur. J. Pharmacol. 2007, 568, 149–163. [Google Scholar] [CrossRef]
- Tighilet, B.; Trottier, S.; Mourre, C.; Lacour, M. Changes in the histaminergic system during vestibular compensation in the cat. J. Physiol. 2006, 573 Pt 3, 723–739. [Google Scholar] [CrossRef]
- Lacour, M.; Tighilet, B. Vestibular compensation in the cat: The role of the histaminergic system. Acta Oto-Laryngol. 2000, 544, 15–18. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | 5-HT | NE | DA | Histamine |
|---|---|---|---|---|
| Sham | 0.733 ± 0.012 | 1.257 ± 0.081 | 0.070 ± 0.006 | 0.053 ± 0.003 |
| UL-4 h | 0.830 ± 0.012 | 1.387 ± 0.074 | 0.077 ± 0.003 | 0.057 ± 0.003 |
| UL-8 h | 0.913 ± 0.015 | 1.600 ± 0.061 | 0.093 ± 0.007 | 0.067 ± 0.007 |
| UL-1 d | 1.090 ± 0.025 | 1.857 ± 0.088 | 0.110 ± 0.006 | 0.077 ± 0.003 |
| UL-2 d | 0.983 ± 0.030 | 2.333 ± 0.114 | 0.127 ± 0.007 | 0.070 ± 0.006 |
| UL-4 d | 0.843 ± 0.028 | 1.553 ± 0.082 | 0.127 ± 0.003 | 0.053 ± 0.003 |
| UL-7 d | 0.760 ± 0.025 | 1.497 ± 0.068 | 0.107 ± 0.007 | 0.053 ± 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Tian, E.; Zhang, Y.; Guo, Z.; Chen, J.; Kong, W.; Lu, Y.; Zhang, S. The Effects of Unilateral Labyrinthectomy on Monoamine Neurotransmitters in the Medial Vestibular Nucleus of Rats. Biomolecules 2023, 13, 1637. https://doi.org/10.3390/biom13111637
Wang J, Tian E, Zhang Y, Guo Z, Chen J, Kong W, Lu Y, Zhang S. The Effects of Unilateral Labyrinthectomy on Monoamine Neurotransmitters in the Medial Vestibular Nucleus of Rats. Biomolecules. 2023; 13(11):1637. https://doi.org/10.3390/biom13111637
Chicago/Turabian StyleWang, Jun, E Tian, Yuejin Zhang, Zhaoqi Guo, Jingyu Chen, Weijia Kong, Yisheng Lu, and Sulin Zhang. 2023. "The Effects of Unilateral Labyrinthectomy on Monoamine Neurotransmitters in the Medial Vestibular Nucleus of Rats" Biomolecules 13, no. 11: 1637. https://doi.org/10.3390/biom13111637
APA StyleWang, J., Tian, E., Zhang, Y., Guo, Z., Chen, J., Kong, W., Lu, Y., & Zhang, S. (2023). The Effects of Unilateral Labyrinthectomy on Monoamine Neurotransmitters in the Medial Vestibular Nucleus of Rats. Biomolecules, 13(11), 1637. https://doi.org/10.3390/biom13111637