
Force-Regulated Calcium Signaling of Lymphoid Cell RPMI 8226 Mediated by Integrin α4β7/MAdCAM-1 in Flow


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
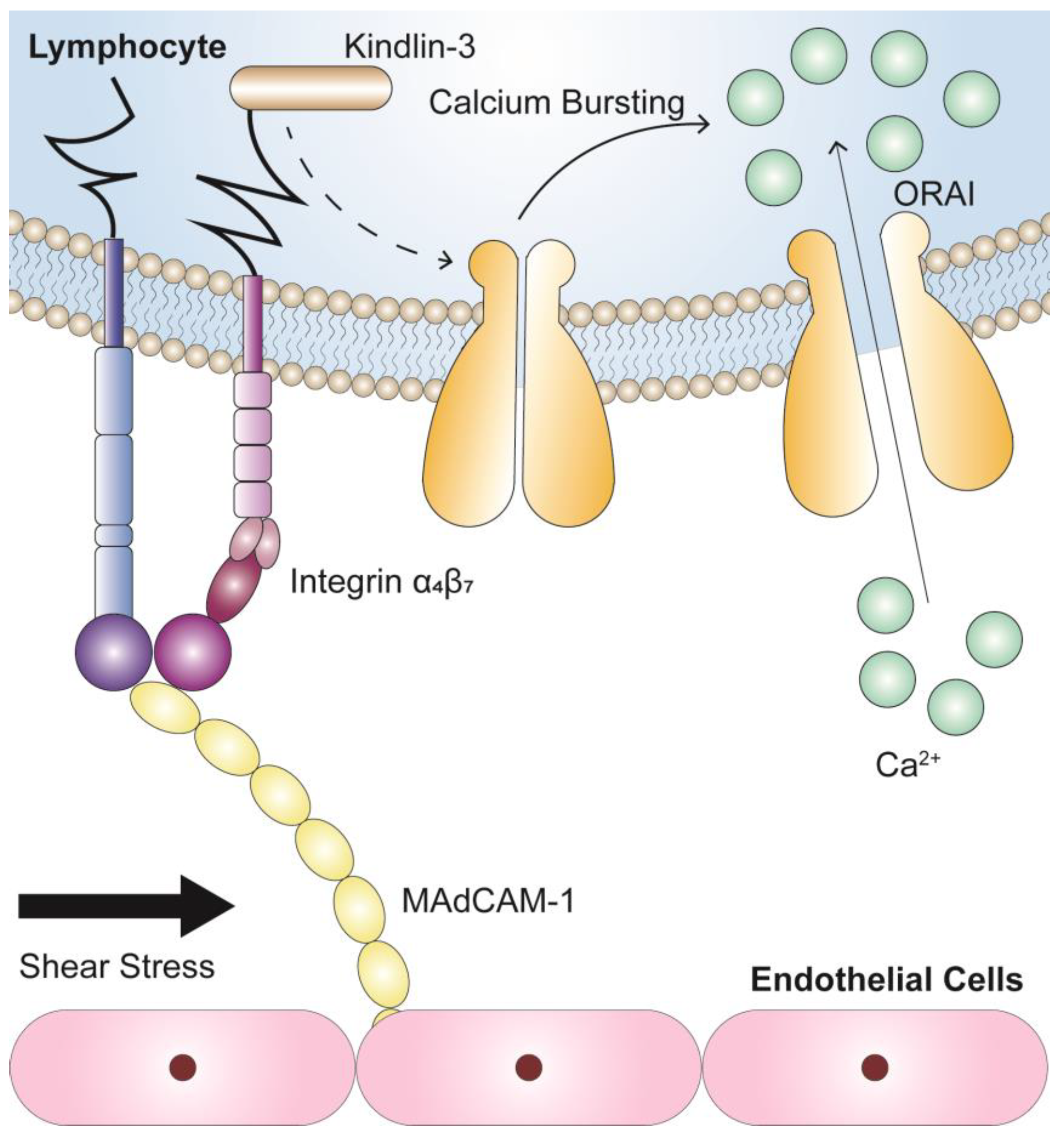{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Proteins and Cells
2.2. Cell Transfection
2.3. Measurement with Flow Cytometry and Western Blotting
2.4. Functionalization of Flow Chamber Substrates
2.5. Loading with Calcium-Sensitive Dye and Treatment with Inhibitors
2.6. Flow Chamber and Calcium Signaling Assays
2.7. Statistical Methods
3. Results
3.1. Calcium Bursting of RPMI 8226 Cells Adhered to MAdCAM-1 under Flow
3.2. Concentration Dependence of Calcium Signaling of RPMI 8226 Cells
3.3. Shear Stresses Quickened and Enhanced Calcium Signaling of RPMI 8226 Cells
3.4. Integrin α4β7 Induced Calcium Influx Rather than Release into the Cytoplasm of RPMI 8226 Cells
3.5. Kindlin-3 Was Involved in Calcium Signaling of RPMI 8226 Cells Triggered by the Integrin α4β7 in Flow
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geremia, A.; Biancheri, P.; Allan, P.; Corazza, G.R.; Di Sabatino, A. Innate and adaptive immunity in inflammatory bowel disease. Autoimmun. Rev. 2014, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- McCully, M.L.; Kouzeli, A.; Moser, B. Peripheral tissue chemokines: Homeostatic control of immune surveillance T cells. Trends Immunol. 2018, 39, 734–747. [Google Scholar] [CrossRef] [PubMed]
- Zundler, S.; Becker, E.; Schulze, L.L.; Neurath, M.F. Immune cell trafficking and retention in inflammatory bowel disease: Mechanistic insights and therapeutic advances. Gut 2019, 68, 1688–1700. [Google Scholar] [CrossRef] [PubMed]
- Lamb, C.A.; O’Byrne, S.; Keir, M.E.; Butcher, E.C. Gut-selective integrin-targeted therapies for inflammatory bowel disease. J. Crohns. Colitis. 2018, 12, S653–S668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargatze, R.F.; Jutila, M.A.; Butcher, E.C. Distinct roles of L-selectin and integrins alpha 4 beta 7 and LFA-1 in lymphocyte homing to Peyer’s patch-HEV in situ: The multistep model confirmed and refined. Immunity 1995, 3, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Shyjan, A.M.; Bertagnolli, M.; Kenney, C.J.; Briskin, M.J. Human mucosal addressin cell adhesion molecule-1 (MAdCAM-1) demonstrates structural and functional similarities to the alpha 4 beta 7-integrin binding domains of murine MAdCAM-1, but extreme divergence of mucin-like sequences. J. Immunol. 1996, 156, 2851–2857. [Google Scholar] [CrossRef]
- Berlin, C.; Berg, E.L.; Briskin, M.J.; Andrew, D.P.; Kilshaw, P.J.; Holzmann, B.; Weissman, I.L.; Hamann, A.; Butcher, E.C. Alpha 4 beta 7 integrin mediates lymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell 1993, 74, 185–195. [Google Scholar] [CrossRef]
- Sjaastad, M.D.; Nelson, W.J. Integrin-mediated calcium signaling and regulation of cell adhesion by intracellular calcium. Bioessays 1997, 19, 47–55. [Google Scholar] [CrossRef]
- Trebak, M.; Kinet, J.P. Calcium signalling in T cells. Nat. Rev. Immunol. 2019, 19, 154–169. [Google Scholar] [CrossRef]
- Feske, S. Calcium signalling in lymphocyte activation and disease. Nat. Rev. Immunol. 2007, 7, 690–702. [Google Scholar] [CrossRef]
- Huang, B.; Ling, Y.; Lin, J.; Fang, Y.; Wu, J. Mechanical regulation of calcium signaling of HL-60 on P-selectin under flow. Biomed. Eng. Online 2016, 15, 153. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Ling, Y.; Lin, J.; Du, X.; Fang, Y.; Wu, J. Force-dependent calcium signaling and its pathway of human neutrophils on P-selectin in flow. Protein Cell 2017, 8, 103–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Liu, J.; Zheng, Y.; Pan, Y.; Zhang, K.; Chen, J. Distinct chemokine signaling regulates integrin ligand specificity to dictate tissue-specific lymphocyte homing. Dev. Cell 2014, 30, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Chen, W.; Evavold, B.D.; Zhu, C. Accumulation of dynamic catch bonds between TCR and agonist peptide-MHC triggers T cell signaling. Cell 2014, 157, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Dixit, N.; Kim, M.H.; Rossaint, J.; Yamayoshi, I.; Zarbock, A.; Simon, S.I. Leukocyte function antigen-1, kindlin-3, and calcium flux orchestrate neutrophil recruitment during inflammation. J. Immunol. 2012, 189, 5954–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morikis, V.A.; Masadeh, E.; Simon, S.I. Tensile force transmitted through LFA-1 bonds mechanoregulate neutrophil inflammatory response. J. Leukoc. Biol. 2020, 108, 1815–1828. [Google Scholar] [CrossRef]
- Abram, C.L.; Lowell, C.A. The ins and outs of leukocyte integrin signaling. Annu. Rev. Immunol. 2009, 27, 339–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognoni, E.; Ruppert, R.; Fassler, R. The kindlin family: Functions, signaling properties and implications for human disease. J. Cell Sci. 2016, 129, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Malinin, N.L.; Plow, E.F.; Byzova, T.V. Kindlins in FERM adhesion. Blood 2010, 115, 4011–4017. [Google Scholar] [CrossRef]
- Moore, G.E.; Kitamura, H. Cell line derived from patient with myeloma. N. Y. State J. Med. 1968, 68, 2054–2060. [Google Scholar] [PubMed]
- Lu, L.; Lin, C.; Yan, Z.; Wang, S.; Zhang, Y.; Wang, S.; Wang, J.; Liu, C.; Chen, J. Kindlin-3 is essential for the resting alpha4beta1 integrin-mediated firm cell adhesion under shear flow conditions. J. Biol. Chem. 2016, 291, 10363–10371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Fang, Y.; Ding, X.; Wu, J. Force-dependent bond dissociation govern rolling of HL-60 cells through E-selectin. Exp. Cell Res. 2012, 318, 1649–1658. [Google Scholar] [CrossRef]
- Sun, X.; Huang, B.; Pan, Y.; Fang, J.; Wang, H.; Ji, Y.; Ling, Y.; Guo, P.; Lin, J.; Li, Q.; et al. Spatiotemporal characteristics of P-selectin-induced β2 integrin activation of human neutrophils under flow. Front. Immunol. 2022, 13, 1023865. [Google Scholar] [CrossRef]
- Kanner, S.B.; Grosmaire, L.S.; Ledbetter, J.A.; Damle, N.K. Beta 2-integrin LFA-1 signaling through phospholipase C-gamma 1 activation. Proc. Natl. Acad. Sci. USA 1993, 90, 7099–7103. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, W.S.; Kulkarni, S.; Giuliano, S.; Goncalves, I.; Dopheide, S.M.; Yap, C.L.; Harper, I.S.; Salem, H.H.; Jackson, S.P. Distinct glycoprotein Ib/V/IX and integrin alpha IIbbeta 3-dependent calcium signals cooperatively regulate platelet adhesion under flow. J. Biol. Chem. 2002, 277, 2965–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimolo, Z.; Wesolowski, G.; Tanaka, H.; Hyman, J.L.; Hoyer, J.R.; Rodan, G.A. Soluble alpha v beta 3-integrin ligands raise [Ca2+]i in rat osteoclasts and mouse-derived osteoclast-like cells. Am. J. Physiol. 1994, 266, C376–C381. [Google Scholar] [CrossRef]
- Dixit, N.; Simon, S.I. Chemokines, selectins and intracellular calcium flux: Temporal and spatial cues for leukocyte arrest. Front. Immunol. 2012, 3, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEver, R.P.; Zhu, C. Rolling cell adhesion. Annu. Rev. Cell Dev. Biol. 2010, 26, 363–396. [Google Scholar] [CrossRef]
- Long, M.; Sato, M.; Lim, C.T.; Wu, J.H.; Adachi, T.; Inoue, Y. Advances in experiments and modeling in micro- and nano-biomechanics: A mini review. Cell Mol. Bioeng. 2011, 4, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Dai, C.; Shen, H.; Gu, M.; Wang, Y.; Liu, J.; Chen, L.; Sun, L. Recent advances on the model, measurement technique, and application of single cell mechanics. Int. J. Mol. Sci. 2020, 21, 6248. [Google Scholar] [CrossRef]
- Yu, X.; Qiu, S.; Sun, D.; Guo, P.; Li, Q. Comparison of intermolecular interactions of irreversible and reversible inhibitors with Bruton’s tyrosine kinase via molecular dynamics simulations. Molecules 2022, 27, 7451. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Liu, Y.; Li, Q. A mechanism for localized dynamics-driven activation in Bruton’s tyrosine kinase. R. Soc. Open Sci. 2021, 8, 210066. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Qiu, S.; Fang, Y.; Wu, J.; Li, Q. Comparison of linear vs. cyclic RGD pentapeptide interactions with integrin alpha(v)beta(3) by molecular dynamics simulations. Biology 2021, 10, 688. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Lin, J.; Cruz, M.A.; Dong, J.F.; Zhu, C. Force-induced cleavage of single VWFA1A2A3 tridomains by ADAMTS-13. Blood 2010, 115, 370–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilkan, Z.; Wright, J.R.; Goodall, A.H.; Gibbins, J.M.; Jones, C.I.; Mahaut-Smith, M.P. Evidence for shear-mediated Ca(2+) entry through mechanosensitive cation channels in human platelets and a megakaryocytic cell line. J. Biol. Chem. 2017, 292, 9204–9217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Chateau, M.; Chen, S.; Salas, A.; Springer, T.A. Kinetic and mechanical basis of rolling through an integrin and novel Ca2+-dependent rolling and Mg2+-dependent firm adhesion modalities for the alpha 4 beta 7-MAdCAM-1 interaction. Biochemistry 2001, 40, 13972–13979. [Google Scholar] [CrossRef]
- Moser, M.; Bauer, M.; Schmid, S.; Ruppert, R.; Schmidt, S.; Sixt, M.; Wang, H.V.; Sperandio, M.; Fassler, R. Kindlin-3 is required for beta2 integrin-mediated leukocyte adhesion to endothelial cells. Nat. Med. 2009, 15, 300–305. [Google Scholar] [CrossRef]
- Lefort, C.T.; Rossaint, J.; Moser, M.; Petrich, B.G.; Zarbock, A.; Monkley, S.J.; Critchley, D.R.; Ginsberg, M.H.; Fassler, R.; Ley, K. Distinct roles for talin-1 and kindlin-3 in LFA-1 extension and affinity regulation. Blood 2012, 119, 4275–4282. [Google Scholar] [CrossRef]
- Xu, Z.; Ni, B.; Cao, Z.; Zielonka, J.; Gao, J.; Chen, F.; Kalyanaraman, B.; White, G.C.; Ma, Y.Q. Kindlin-3 negatively regulates the release of neutrophil extracellular traps. J. Leukoc. Biol. 2018, 104, 597–602. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, D.; Luo, Z.; Kong, Y.; Huang, R.; Li, Q. Force-Regulated Calcium Signaling of Lymphoid Cell RPMI 8226 Mediated by Integrin α4β7/MAdCAM-1 in Flow. Biomolecules 2023, 13, 587. https://doi.org/10.3390/biom13040587
Sun D, Luo Z, Kong Y, Huang R, Li Q. Force-Regulated Calcium Signaling of Lymphoid Cell RPMI 8226 Mediated by Integrin α4β7/MAdCAM-1 in Flow. Biomolecules. 2023; 13(4):587. https://doi.org/10.3390/biom13040587
Chicago/Turabian StyleSun, Dongshan, Zhiqing Luo, Ying Kong, Ruiting Huang, and Quhuan Li. 2023. "Force-Regulated Calcium Signaling of Lymphoid Cell RPMI 8226 Mediated by Integrin α4β7/MAdCAM-1 in Flow" Biomolecules 13, no. 4: 587. https://doi.org/10.3390/biom13040587
APA StyleSun, D., Luo, Z., Kong, Y., Huang, R., & Li, Q. (2023). Force-Regulated Calcium Signaling of Lymphoid Cell RPMI 8226 Mediated by Integrin α4β7/MAdCAM-1 in Flow. Biomolecules, 13(4), 587. https://doi.org/10.3390/biom13040587