

Analysis of CDR3 Sequences from T-Cell Receptor β in Acute Respiratory Distress Syndrome
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Immune Sequencing, and Retrieval of CDR3 Sequences
2.2. Protein Isoelectric Point
2.3. Immunogenicity Analysis
2.4. Protein Residue Statistics
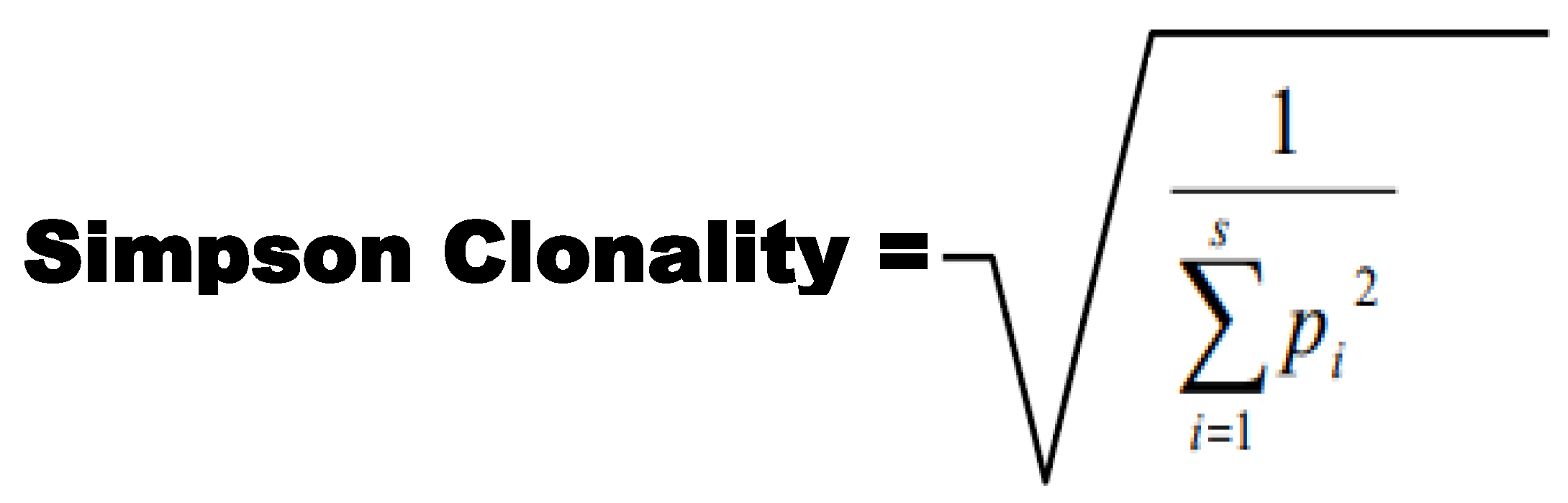
2.5. CDR3 Clonality
2.6. Statistical Analysis
3. Results
3.1. Sequence Analysis
3.1.1. CDR3 Sequences Have Varying Length Distributions
3.1.2. Immunogenicity Ratio of CDR3 Sequences
3.2. Biophysiochemical Properties
3.2.1. CDR3 Residues Have Variations in Isoelectric Point
3.2.2. CDR3 Protein Residue Biochemical Properties
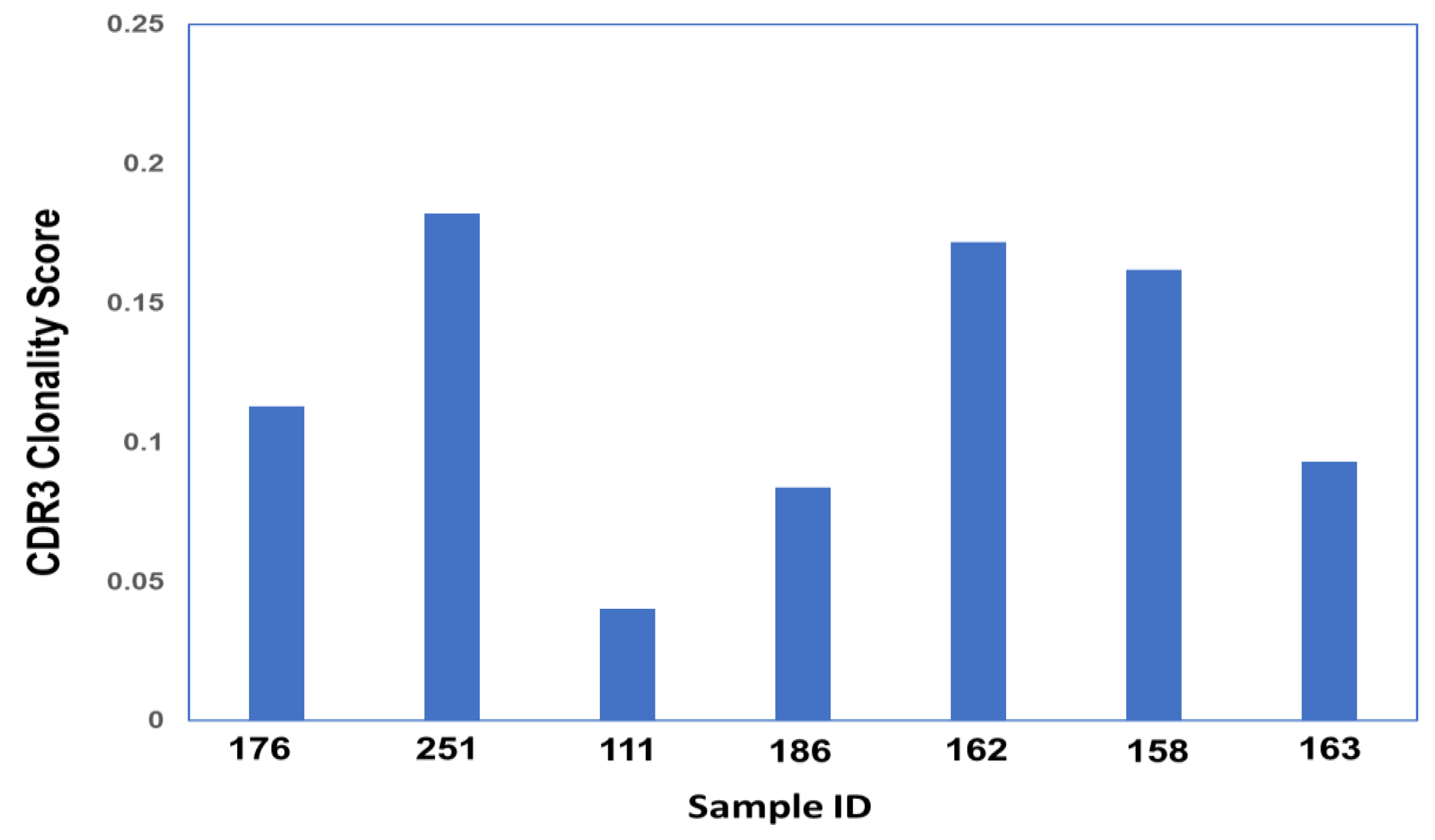
3.3. Clonality
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quintanilla, E.; Diwa, K.; Nguyen, A.; Vu, L.; Toby, I.T. A Data Report on the Curation and Development of a Database of Genes for Acute Respiratory Distress Syndrome. Front. Genet. 2021, 12, 750568. [Google Scholar] [CrossRef] [PubMed]
- Alberti, C.; Brun-Buisson, C.; Goodman, S.V.; Guidici, D.; Granton, J.; Moreno, R.; Smithies, M.; Thomas, O.; Artigas, A.; Le Gall, J.R. Influence of systemic inflammatory response syndrome and sepsis on outcome of critically ill infected patients. Am. J. Respir. Crit. Care Med. 2003, 168, 77–84. [Google Scholar] [CrossRef]
- Katzenstein, A.L.; Bloor, C.M.; Leibow, A.A. Diffuse alveolar damage--the role of oxygen, shock, and related factors. A review. Am. J. Pathol. 1976, 85, 209–228. [Google Scholar]
- Matute-Bello, G.; Frevert, C.W.; Martin, T.R. Animal models of acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, L379–L399. [Google Scholar] [CrossRef] [PubMed]
- Blondonnet, R.; Constantin, J.M.; Sapin, V.; Jabaudon, M. A Pathophysiologic Approach to Biomarkers in Acute Respiratory Distress Syndrome. Dis. Markers 2016, 2016, 3501373. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.M.; Wilson, J.; Ware, L.B. Biomarkers in acute respiratory distress syndrome: From pathobiology to improving patient care. Expert Rev. Respir. Med. 2014, 8, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, H.Y.; Fulmer, J.D.; Kazmierowski, J.A.; Roberts, W.C.; Frank, M.M.; Crystal, R.G. Analysis of cellular and protein content of broncho-alveolar lavage fluid from patients with idiopathic pulmonary fibrosis and chronic hypersensitivity pneumonitis. J. Clin. Investig. 1977, 59, 165–175. [Google Scholar] [CrossRef]
- Rubelt, F.; The AIRR Community; Busse, C.E.; Bukhari, S.A.C.; Bürckert, J.-P.; Mariotti-Ferrandiz, E.; Cowell, L.G.; Watson, C.T.; Marthandan, N.; Faison, W.J.; et al. Adaptive Immune Receptor Repertoire Community recommendations for sharing immune-repertoire sequencing data. Nat. Immunol. 2017, 18, 1274–1278. [Google Scholar] [CrossRef]
- Miqueu, P.; Guillet, M.; Degauque, N.; Doré, J.C.; Soulillou, J.P.; Brouard, S. Statistical analysis of CDR3 length distributions for the assessment of T and B cell repertoire biases. Mol. Immunol. 2007, 44, 1057–1064. [Google Scholar] [CrossRef]
- Tonegawa, S. Somatic generation of antibody diversity. Nature 1983, 302, 575–581. [Google Scholar] [CrossRef]
- Christley, S.; Scarborough, W.; Salinas, E.; Rounds, W.H.; Toby, I.T.; Fonner, J.M.; Levin, M.K.; Kim, M.; Mock, S.A.; Jordan, C.; et al. VDJServer: A Cloud-Based Analysis Portal and Data Commons for Immune Repertoire Sequences and Rearrangements. Front. Immunol. 2018, 9, 976. [Google Scholar] [CrossRef] [PubMed]
- Gorski, J.; Yassai, M.; Zhu, X.; Kissela, B.; Kissela, B.; Keever, C.; Flomenberg, N. Circulating T cell repertoire complexity in normal individuals and bone marrow recipients analyzed by CDR3 size spectratyping. Correlation with immune status. J. Immunol. 1994, 152, 5109–5119. [Google Scholar] [CrossRef] [PubMed]
- Heiden, J.A.V.; Marquez, S.; Marthandan, N.; Bukhari, S.A.C.; Busse, C.; Corrie, B.; Hershberg, U.; Kleinstein, S.H.; Iv, F.A.M.; Ralph, D.K.; et al. AIRR Community Standardized Representations for Annotated Immune Repertoires. Front. Immunol. 2018, 9, 2206. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.L.; Freeman, J.D.; Zeng, T.; Choe, G.; Munro, S.; Moore, R.; Webb, J.R.; Holt, R.A. Exhaustive T-cell repertoire sequencing of human peripheral blood samples reveals signatures of antigen selection and a directly measured repertoire size of at least 1 million clonotypes. Genome Res. 2011, 21, 790–797. [Google Scholar] [CrossRef]
- Ye, J.; Ma, N.; Madden, T.L.; Ostell, J.M. IgBLAST: An immunoglobulin variable domain sequence analysis tool. Nucleic Acids Res. 2013, 41, W34–W40. [Google Scholar] [CrossRef]
- Fink, P.J.; Matis, L.A.; McElligott, D.L.; Bookman, M.; Hedrick, S.M. Correlations between T-cell specificity and the structure of the antigen receptor. Nature 1986, 321, 219–226. [Google Scholar] [CrossRef]
- Barth, R.K.; Kim, B.S.; Lan, N.C.; Hunkapiller, T.; Sobieck, N.; Winoto, A.; Gershenfeld, H.; Okada, C.; Hansburg, D.; Weissman, I.L.; et al. The murine T-cell receptor uses a limited repertoire of expressed V beta gene segments. Nature 1985, 316, 517–523. [Google Scholar] [CrossRef]
- Danska, J.S.; Livingstone, A.M.; Paragas, V.; Ishihara, T.; Fathman, C.G. The presumptive CDR3 regions of both T cell receptor alpha and beta chains determine T cell specificity for myoglobin peptides. J. Exp. Med. 1990, 172, 27–33. [Google Scholar] [CrossRef]
- QIAprep Miniprep Handbook 12/2020. Available online: www.qiagen.com (accessed on 26 January 2022).
- Carlson, C.S.; Emerson, R.O.; Sherwood, A.M.; Desmarais, C.; Chung, M.-W.; Parsons, J.M.; Steen, M.S.; LaMadrid-Herrmannsfeldt, M.A.; Williamson, D.W.; Livingston, R.J.; et al. Using synthetic templates to design an unbiased multiplex PCR assay. Nat. Commun. 2013, 4, 2680. [Google Scholar] [CrossRef]
- Stothard, P.; Hiseni, P.; Wilson, R.C.; Storrø, O.; Johnsen, R.; Øien, T.; Rudi, K. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef]
- Calis, J.J.A.; Maybeno, M.; Greenbaum, J.A.; Weiskopf, D.; De Silva, A.D.; Sette, A.; Keşmir, C.; Peters, B. Properties of MHC class I presented peptides that enhance immunogenicity. PLoS Comput. Biol. 2013, 9, e1003266. [Google Scholar] [CrossRef]
- Wong, G.K.; Millar, D.; Penny, S.; Heather, J.M.; Mistry, P.; Buettner, N.; Bryon, J.; Huissoon, A.P.; Cobbold, M. Accelerated Loss of TCR Repertoire Diversity in Common Variable Immunodeficiency. J. Immunol. 2016, 197, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Hohendorf, T.; Mohan, H.; Bien, C.G.; Breuer, J.; Becker, A.; Görlich, D.; Kuhlmann, T.; Widman, G.; Herich, S.; Elpers, C.; et al. CD8(+) T-cell pathogenicity in Rasmussen encephalitis elucidated by large-scale T-cell receptor sequencing. Nat. Commun. 2016, 7, 11153. [Google Scholar] [CrossRef] [PubMed]
- Weir, G.M.; Hrytsenko, O.; Quinton, T.; Berinstein, N.L.; Stanford, M.M.; Mansour, M. Anti-PD-1 increases the clonality and activity of tumor infiltrating antigen specific T cells induced by a potent immune therapy consisting of vaccine and metronomic cyclophosphamide. J. Immunother. Cancer 2016, 4, 68. [Google Scholar] [CrossRef]
- Nunes-Alves, C.; Booty, M.G.; Carpenter, S.M.; Rothchild, A.C.; Martin, C.; Desjardins, D.; Steblenko, K.; Kløverpris, H.N.; Madansein, R.; Ramsuran, D.; et al. Human and Murine Clonal CD8+ T Cell Expansions Arise during Tuberculosis Because of TCR Selection. PLoS Pathog. 2015, 11, e1004849. [Google Scholar] [CrossRef]
- Suessmuth, Y.; Mukherjee, R.; Watkins, B.; Koura, D.T.; Finstermeier, K.; Desmarais, C.; Stempora, L.; Horan, J.T.; Langston, A.; Qayed, M.; et al. CMV reactivation drives posttransplant T-cell reconstitution and results in defects in the underlying TCRβ repertoire. Blood 2015, 125, 3835–3850. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, D.; Wilkinson, G.A.; Eng, K.H.; Fields, P.; Raber, P.; Moseley, J.L.; Cheetham, K.; Coffey, M.; Nuovo, G.; Kalinski, P.; et al. Pembrolizumab in Combination with the Oncolytic Virus Pelareorep and Chemotherapy in Patients with Advanced Pancreatic Adenocarcinoma: A Phase Ib Study. Clin. Cancer Res. 2020, 26, 71–81. [Google Scholar] [CrossRef]
- Morris, H.; DeWolf, S.; Robins, H.; Sprangers, B.; LoCascio, S.A.; Shonts, B.A.; Kawai, T.; Wong, W.; Yang, S.; Zuber, J.; et al. Tracking donor-reactive T cells: Evidence for clonal deletion in tolerant kidney transplant patients. Sci. Transl. Med. 2015, 7, 272ra10. [Google Scholar] [CrossRef]
- Roh, W.; Chen, P.-L.; Reuben, A.; Spencer, C.N.; Prieto, P.A.; Miller, J.P.; Gopalakrishnan, V.; Wang, F.; Cooper, Z.A.; Reddy, S.M.; et al. Integrated molecular analysis of tumor biopsies on sequential CTLA-4 and PD-1 blockade reveals markers of response and resistance. Sci. Transl. Med. 2017, 9, eaah3560. [Google Scholar] [CrossRef]
- Zhu, W.; Germain, C.; Liu, Z.; Sebastian, Y.; Devi, P.; Knockaert, S.; Brohawn, P.; Lehmann, K.; Damotte, D.; Validire, P.; et al. A high density of tertiary lymphoid structure B cells in lung tumors is associated with increased CD4+ T cell receptor repertoire clonality. Oncoimmunology 2015, 4, e1051922. [Google Scholar] [CrossRef]
- Wu, X.; Deng, Z.; Zhao, Q. Immunotherapy improves disease prognosis by affecting the tumor microenvironment: A bibliometric study. Front. Immunol. 2022, 13, 967076. [Google Scholar] [CrossRef] [PubMed]
- Olschewski, V.; Witte, H.M.; Bernard, V.; Steinestel, K.; Peter, W.; Merz, H.; Rieken, J.; Biersack, H.; von Bubnoff, N.; Feller, A.C.; et al. Systemic Inflammation and Tumour-Infiltrating T-Cell Receptor Repertoire Diversity Are Predictive of Clinical Outcome in High-Grade B-Cell Lymphoma with MYC and BCL2 and/or BCL6 Rearrangements. Cancers 2021, 13, 887. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, I.R.; Watanabe, R.; O’malley, J.T.; Williamson, D.W.; Scott, L.-L.; Elco, C.P.; Teague, J.E.; Gehad, A.; Lowry, E.L.; LeBoeuf, N.R.; et al. TCR sequencing facilitates diagnosis and identifies mature T cells as the cell of origin in CTCL. Sci. Transl. Med. 2015, 7, 308ra158. [Google Scholar] [CrossRef]
- Hershberg, U.; Prak, E.T.L. The analysis of clonal expansions in normal and autoimmune B cell repertoires. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140239. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Sherwood, A.; Fromm, J.R.; Winter, S.S.; Dunsmore, K.P.; Loh, M.L.; Greisman, H.A.; Sabath, D.E.; Wood, B.L.; Robins, H. High-throughput sequencing detects minimal residual disease in acute T lymphoblastic leukemia. Sci. Transl. Med. 2012, 4, 134ra63. [Google Scholar] [CrossRef]
- Carey, A.J.; Hope, J.L.; Mueller, Y.M.; Fike, A.J.; Kumova, O.K.; van Zessen, D.B.H.; Steegers, E.A.P.; van der Burg, M.; Katsikis, P.D. Public Clonotypes and Convergent Recombination Characterize the Naïve CD8+ T-Cell Receptor Repertoire of Extremely Preterm Neonates. Front. Immunol. 2017, 8, 1859. [Google Scholar] [CrossRef]
- Yeh, W.I.; Seay, H.R.; Newby, B.; Posgai, A.L.; Moniz, F.B.; Michels, A.; Mathews, C.E.; Bluestone, J.A.; Brusko, T.M. Avidity and Bystander Suppressive Capacity of Human Regulatory T Cells Expressing De Novo Autoreactive T-Cell Receptors in Type 1 Diabetes. Front. Immunol. 2017, 8, 1313. [Google Scholar] [CrossRef]
- Emerson, R.O.; DeWitt, W.S.; Vignali, M.; Gravley, J.; Hu, J.K.; Osborne, E.J.; Desmarais, C.; Klinger, M.; Carlson, C.S.; Hansen, J.A.; et al. Immunosequencing identifies signatures of cytomegalovirus exposure history and HLA-mediated effects on the T cell repertoire. Nat. Genet. 2017, 49, 659–665. [Google Scholar] [CrossRef]
- Lindau, P.; Mukherjee, R.; Gutschow, M.V.; Vignali, M.; Warren, E.H.; Riddell, S.R.; Makar, K.W.; Turtle, C.J.; Robins, H.S. Cytomegalovirus Exposure in the Elderly Does Not Reduce CD8 T Cell Repertoire Diversity. J. Immunol. 2019, 202, 476–483. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 3 October 2022).
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at EMBL-EBI. Nucleic Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef] [PubMed]
- Toby, I.T.; Levin, M.K.; Salinas, E.A.; Christley, S.; Bhattacharya, S.; Breden, F.; Buntzman, A.; Corrie, B.; Fonner, J.; Gupta, N.T.; et al. VDJML: A file format with tools for capturing the results of inferring immune receptor rearrangements. BMC Bioinform. 2016, 17, 333. [Google Scholar] [CrossRef] [PubMed]
- Huisman, W.; Hageman, L.; Leboux, D.A.T.; Khmelevskaya, A.; Efimov, G.A.; Roex, M.C.J.; Amsen, D.; Falkenburg, J.H.F.; Jedema, I. Public T-Cell Receptors (TCRs) Revisited by Analysis of the Magnitude of Identical and Highly-Similar TCRs in Virus-Specific T-Cell Repertoires of Healthy Individuals. Front. Immunol. 2022, 13, 851868. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsen, K.S.; Høydahl, L.S.; Neumann, R.S.; Bjerregaard-Andersen, K.; Nilssen, N.R.; Sollid, L.M.; Sandlie, I.; Løset, G.Å. Soluble T-cell receptor design influences functional yield in an E. coli chaperone-assisted expression system. PLoS ONE 2018, 13, e0195868. [Google Scholar] [CrossRef]
- Kurosawa, Y.; Tonegawa, S. Organization, structure, and assembly of immunoglobulin heavy chain diversity DNA segments. J. Exp. Med. 1982, 155, 201–218. [Google Scholar] [CrossRef]
- Alamyar, E.; Duroux, P.; Lefranc, M.P.; Giudicelli, V. IMGT(®) tools for the nucleotide analysis of immunoglobulin (IG) and T cell receptor (TR) V-(D)-J repertoires, polymorphisms, and IG mutations: IMGT/V-QUEST and IMGT/HighV-QUEST for NGS. Methods Mol. Biol. 2012, 882, 569–604. [Google Scholar] [CrossRef]
- King, D.J.; Gotch, F.M.; Larsson-Sciard, E.L.; Paediatric European Network for Treatment of AIDS (PENTA). T-cell re-population in HIV-infected children on highly active anti-retroviral therapy (HAART). Clin. Exp. Immunol. 2001, 125, 447–454. [Google Scholar] [CrossRef]
- Gorochov, G.; Neumann, A.U.; Kereveur, A.; Parizot, C.; Li, T.; Katlama, C.; Karmochkine, M.; Raguin, G.; Autran, B.; Debré, P. Perturbation of CD4+ and CD8+ T-cell repertoires during progression to AIDS and regulation of the CD4+ repertoire during antiviral therapy. Nat. Med. 1998, 4, 215–221. [Google Scholar] [CrossRef]
- Hirokawa, M.; Horiuchi, T.; Kitabayashi, A.; Kawabata, Y.; Matsutani, T.; Suzuki, R.; Chihara, J.; Miura, A.B. Delayed recovery of CDR3 complexity of the T-cell receptor-beta chain in recipients of allogeneic bone marrow transplants who had virus-associated interstitial pneumonia: Monitor of T-cell function by CDR3 spectratyping. J. Allergy Clin. Immunol. 2000, 106, S32–S39. [Google Scholar] [CrossRef]
- Nikolich-Žugich, J.; Slifka, M.; Messaoudi, I. The many important facets of T-cell repertoire diversity. Nat. Rev. Immunol. 2004, 4, 123–132. [Google Scholar] [CrossRef]
- Izbicki, G.; Segel, M.J.; Christensen, T.G.; Conner, M.W.; Breuer, R. Time course of bleomycin-induced lung fibrosis. Int. J. Exp. Pathol. 2002, 83, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Kolvenbach, C.M.; Dworschak, G.C.; Frese, S.; Japp, A.S.; Schuster, P.; Wenzlitschke, N.; Yilmaz, Ö.; Lopes, F.M.; Pryalukhin, A.; Schierbaum, L.; et al. Rare Variants in BNC2 Are Implicated in Autosomal-Dominant Congenital Lower Urinary-Tract Obstruction. Am. J. Hum. Genet. 2019, 104, 994–1006. [Google Scholar] [CrossRef]
- Assarsson, E.; Sidney, J.; Oseroff, C.; Pasquetto, V.; Bui, H.H.; Frahm, N.; Brander, C.; Peters, B.; Grey, H.; Sette, A. A quantitative analysis of the variables affecting the repertoire of T cell specificities recognized after vaccinia virus infection. J. Immunol. 2007, 178, 7890–7901. [Google Scholar] [CrossRef] [PubMed]
- Kotturi, M.F.; Botten, J.; Sidney, J.; Bui, H.-H.; Giancola, L.; Maybeno, M.; Babin, J.; Oseroff, C.; Pasquetto, V.; Greenbaum, J.A.; et al. A multivalent and cross-protective vaccine strategy against arenaviruses associated with human disease. PLoS Pathog. 2009, 5, e1000695. [Google Scholar] [CrossRef] [PubMed]
- Ostmeyer, J.; Christley, S.; Toby, I.T.; Cowell, L.G. Biophysicochemical Motifs in T-cell Receptor Sequences Distinguish Repertoires from Tumor-Infiltrating Lymphocyte and Adjacent Healthy Tissue. Cancer Res 2019, 79, 1671–1680. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Positive Scoring Sequences | Total Number of Sequences | Percent % Immunogenicity Observed | Clinical Diagnosis |
|---|---|---|---|---|
| 111 | 515 | 1560 | 33.01% | ARDS |
| 176 | 19 | 56 | 33.93% | ARDS |
| 186 | 501 | 1485 | 33.74% | ARDS |
| 251 | 64 | 202 | 31.68% | ARDS |
| 158 | 7 | 25 | 28.00% | Non-ARDS |
| 162 | 33 | 118 | 27.97% | Non-ARDS |
| 163 | 51 | 169 | 30.18% | Non-ARDS |
| Non-ARDS CDR3 Residues | Times Found | Percentage | ARDS CDR3 Residues | Times Found | Percentage |
|---|---|---|---|---|---|
| Aliphatic G,A,V,L,I | 1270 | 27.46 | Aliphatic G,A,V,L,I | 13,491 | 28.15 |
| Aromatic F,W,Y | 734 | 15.87 | Aromatic F,W,Y | 7858 | 16.4 |
| Sulphur C,M | 334 | 7.22 | Sulphur C,M | 3461 | 7.22 |
| Basic K,R,H | 322 | 6.96 | Basic K,R,H | 2846 | 5.94 |
| Acidic B,D,E,N,Q,Z | 786 | 16.99 | Acidic B,D,E,N,Q,Z | 8228 | 17.17 |
| Aliphatic hydroxyl S,T | 1005 | 21.73 | Aliphatic hydroxyl S,T | 10,611 | 22.14 |
| tRNA synthetase class I Z,E,Q,R,C,M,V,I,L,Y,W | 1668 | 36.06 | tRNA synthetase class I Z,E,Q,R,C,M,V,I,L,Y,W | 17,360 | 36.22 |
| tRNA synthetase class II B,G,A,P,S,T,H,D,N,K,F | 2957 | 63.94 | tRNA synthetase class II B,G,A,P,S,T,H,D,N,K,F | 30,566 | 63.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hey, S.; Whyte, D.; Hoang, M.-C.; Le, N.; Natvig, J.; Wingfield, C.; Onyeama, C.; Howrylak, J.; Toby, I.T. Analysis of CDR3 Sequences from T-Cell Receptor β in Acute Respiratory Distress Syndrome. Biomolecules 2023, 13, 825. https://doi.org/10.3390/biom13050825
Hey S, Whyte D, Hoang M-C, Le N, Natvig J, Wingfield C, Onyeama C, Howrylak J, Toby IT. Analysis of CDR3 Sequences from T-Cell Receptor β in Acute Respiratory Distress Syndrome. Biomolecules. 2023; 13(5):825. https://doi.org/10.3390/biom13050825
Chicago/Turabian StyleHey, Sara, Dayjah Whyte, Minh-Chau Hoang, Nick Le, Joseph Natvig, Claire Wingfield, Charles Onyeama, Judie Howrylak, and Inimary T. Toby. 2023. "Analysis of CDR3 Sequences from T-Cell Receptor β in Acute Respiratory Distress Syndrome" Biomolecules 13, no. 5: 825. https://doi.org/10.3390/biom13050825
APA StyleHey, S., Whyte, D., Hoang, M. -C., Le, N., Natvig, J., Wingfield, C., Onyeama, C., Howrylak, J., & Toby, I. T. (2023). Analysis of CDR3 Sequences from T-Cell Receptor β in Acute Respiratory Distress Syndrome. Biomolecules, 13(5), 825. https://doi.org/10.3390/biom13050825