
9-O Acetylated Gangliosides in Health and Disease



Abstract
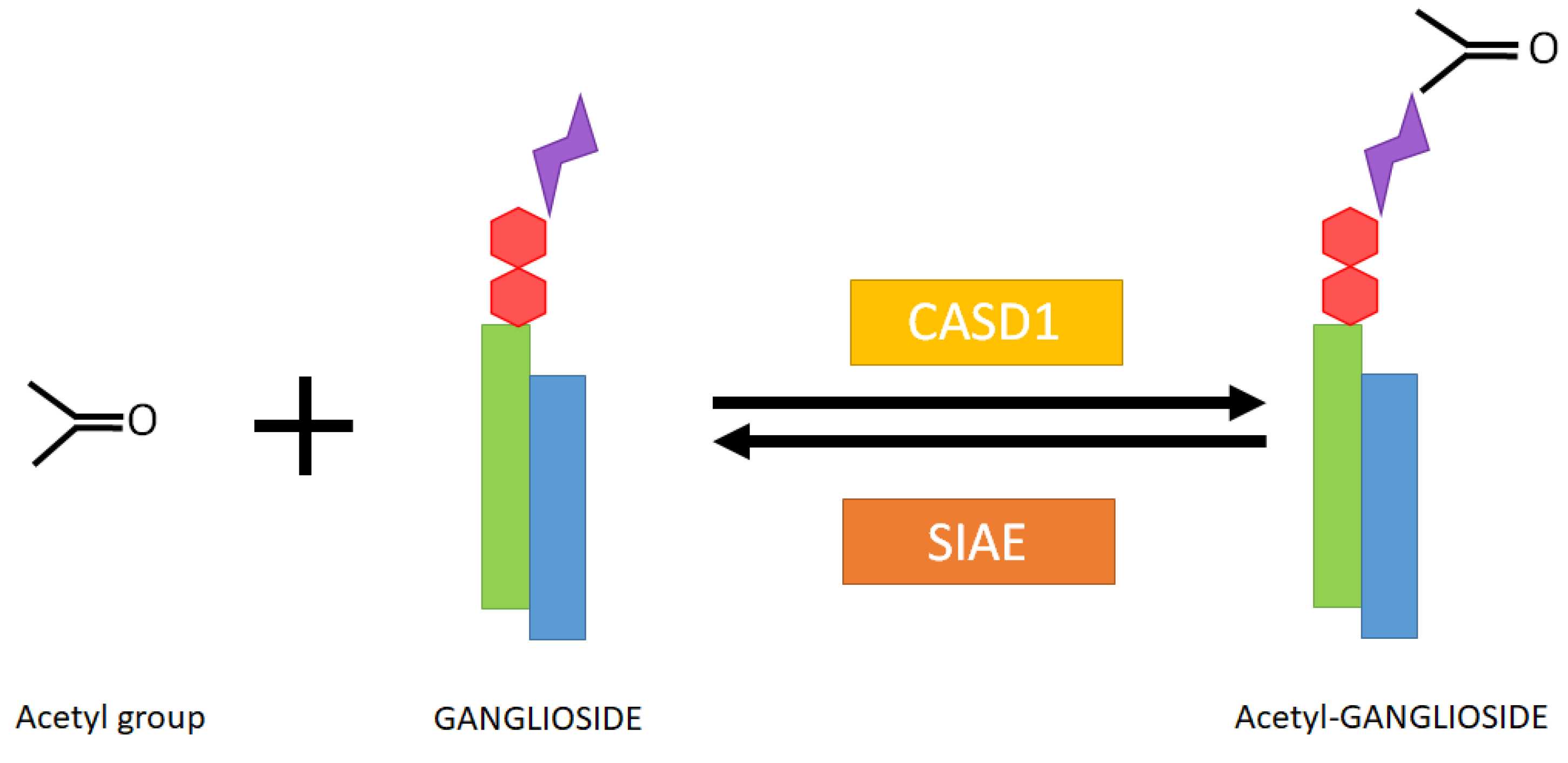
:1. Discovery and Chemistry
1.1. Types of Acetylation and First Findings in Cells
1.2. Chemical Structure and Interactions
1.3. Enzyme Regulation
1.4. Methodological Points
2. 9-O Acetylation of Gangliosides in Pathophysiology
2.1. In Cell Physiology
2.1.1. Embryogenesis
2.1.2. Postnatal Nervous System
2.1.3. Immune System
2.1.4. Hematopoiesis
2.1.5. Kidney
2.2. In Cell Pathology—Diseases
2.2.1. Cancer
2.2.2. Infection
2.2.3. Autoimmune Diseases
2.2.4. Toxicology
3. Concluding Remarks: From Controversy to Future Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 7-O-ac: | 7-O-acetylated |
| 9-N-ac: | 9-N-acetylated |
| 9-O-ac: | 9-O-acetylated |
| 9-O-acLD1: | disialosyl-lacto-N-neotetraosylceramide (LD1) |
| CASD1: | CAS1 domain containing |
| CDw60 = CD60: | 9-O-acetylated GD3 antigen |
| CD60a: | GD3 (non-acetylated) antigen |
| CD60b: | 9-O-acGD3 antigen |
| CD60c: | 7-O-acGD3 antigen |
| CHE-FcD = hemagglutinin Esterase of Influenzavirus C fused to the carboxyl end with human IgG1 Fc region treated with diisopropylfluorophosphate to eradicate its esterase activity. | |
| CM: | confocal microscopy |
| FABMS: | fast atom bombardment mass spectrometry |
| IEM: | immunoelectron Microscopy. |
| IF: | Immunofluorescence |
| IHC: | Immunohistochemistry |
| IP: | Immunoprecipitation |
| N-ac: | N-acetylation |
| NMR: | nuclear magnetic resonance |
| PHA: | Phytohemagglutinin |
| SIAE: | sialate O-acetylesterase |
| SiAOAT: | sialate O-acetyltransferase |
| TLC: | thin-layer chromatography |
References
- Buscher, H.P.; Casals-Stenzel, J.; Schaufer, R. New Sialic Acids. Identification of N-Glycoloyl-O-Acetylneuraminic Acids and N-Acetyl-O-Glycoloylneuraminic Acids by Improved Methods for Detection of N-Acyl and O-Acyl Groups and by Gas-Liquid Chromatography. Eur. J. Biochem. 1974, 50, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Oehler, C.; Kopitz, J.; Cantz, M. Substrate Specificity and Inhibitor Studies of a Membrane-Bound Ganglioside Sialidase Isolated from Human Brain Tissue. Biol. Chem. 2002, 383, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Corfield, A.P.; Ferreira do Amaral, C.; Wember, M.; Schauer, R. The Metabolism of O-Acyl-N-Acylneuraminic Acids. Biosynthesis of O-Acylated Sialic Acids in Bovine and Equine Submandibular Glands. Eur. J. Biochem. 1976, 68, 597–610. [Google Scholar] [CrossRef]
- Baumann, A.-M.T.; Bakkers, M.J.G.; Buettner, F.F.R.; Hartmann, M.; Grove, M.; Langereis, M.A.; de Groot, R.J.; Mühlenhoff, M. 9-O-Acetylation of Sialic Acids Is Catalysed by CASD1 via a Covalent Acetyl-Enzyme Intermediate. Nat. Commun. 2015, 6, 7673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrler, G.; Klenk, H.D. The Surface Receptor Is a Major Determinant of the Cell Tropism of Influenza C Virus. Virology 1987, 159, 102–108. [Google Scholar] [CrossRef]
- Teufel, M.; Roggentin, P.; Schauer, R. Properties of Sialidase Isolated from Actinomyces Viscosus DSM 43798. Biol. Chem. Hoppe Seyler 1989, 370, 435–443. [Google Scholar] [CrossRef]
- Schauer, R.; Reuter, G.; Stoll, S.; Posadas del Rio, F.; Herrler, G.; Klenk, H.D. Isolation and Characterization of Sialate 9(4)-O-Acetylesterase from Influenza C Virus. Biol. Chem. Hoppe Seyler 1988, 369, 1121–1130. [Google Scholar] [CrossRef]
- Hasegawa, A.; Ogawa, H.; Ishida, H.; Kiso, M. Synthesis of an S-(Alpha-Sialosyl)-(2----9)-O-(Alpha-Sialosyl)-(2----3’)-Beta-Lactos Ylceramide. Carbohydr. Res. 1992, 224, 175–184. [Google Scholar] [CrossRef]
- Hunter, C.D.; Khanna, N.; Richards, M.R.; Rezaei Darestani, R.; Zou, C.; Klassen, J.S.; Cairo, C.W. Human Neuraminidase Isoenzymes Show Variable Activities for 9-O-Acetyl-Sialoside Substrates. ACS Chem. Biol. 2018, 13, 922–932. [Google Scholar] [CrossRef]
- Haverkamp, J.; Veh, R.W.; Sander, M.; Schauer, R.; Kamerling, J.P.; Vliegenthart, J.G. Demonstration of 9-O-Acetyl-N-Acetylneuraminic Acid in Brain Gangliosides from Various Vertebrates Including Man. Hoppe Seylers Z Physiol. Chem. 1977, 358, 1609–1612. [Google Scholar] [CrossRef] [Green Version]
- Ghidoni, R.; Sonnino, S.; Tettamanti, G.; Baumann, N.; Reuter, G.; Schauer, R. Isolation and Characterization of a Trisialoganglioside from Mouse Brain, Containing 9-O-Acetyl-N-Acetylneuraminic Acid. J. Biol. Chem. 1980, 255, 6990–6995. [Google Scholar] [CrossRef]
- Chigorno, V.; Sonnino, S.; Ghidoni, R.; Tettamanti, G. Isolation and Characterization of a Tetrasialoganglioside from Mouse Brain, Containing 9-O-Acetyl,N-Acetylneuraminic Acid. Neurochem. Int. 1982, 4, 531–539. [Google Scholar] [CrossRef]
- Hirabayashi, Y.; Li, Y.T.; Li, S.C. Occurrence of a New Hematoside in the Kidney of Guinea Pig. FEBS Lett. 1983, 161, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Bonafede, D.M.; Macala, L.J.; Constantine-Paton, M.; Yu, R.K. Isolation and Characterization of Ganglioside 9-O-Acetyl-GD3 from Bovine Buttermilk. Lipids 1989, 24, 680–684. [Google Scholar] [CrossRef]
- Ren, S.; Scarsdale, J.N.; Ariga, T.; Zhang, Y.; Klein, R.A.; Hartmann, R.; Kushi, Y.; Egge, H.; Yu, R.K. O-Acetylated Gangliosides in Bovine Buttermilk. Characterization of 7-O-Acetyl, 9-O-Acetyl, and 7,9-Di-O-Acetyl GD3. J. Biol. Chem. 1992, 267, 12632–12638. [Google Scholar] [CrossRef]
- Waki, H.; Masuzawa, A.; Kon, K.; Ando, S. A New O-Acetylated Trisialoganglioside, 9-O-Acetyl GT2, in Cod Brain. J. Biochem. 1993, 114, 459–462. [Google Scholar] [CrossRef]
- Waki, H.; Murata, A.; Kon, K.; Maruyama, K.; Kimura, S.; Ogura, H.; Ando, S. Isolation and Characterization of a Trisialyllactosylceramide, GT3, Containing an O-Acetylated Sialic Acid in Cod Fish Brain. J. Biochem. 1993, 113, 502–507. [Google Scholar] [CrossRef]
- Gowda, D.C.; Reuter, G.; Shukla, A.K.; Schauer, R. Identification of a Disialoganglioside (GD1a) Containing Terminal N-Acetyl-9-O-Acetylneuraminic Acid in Rat Erythrocytes. Hoppe Seylers Z Physiol. Chem. 1984, 365, 1247–1253. [Google Scholar] [CrossRef]
- Yachida, Y.; Tsuchihashi, K.; Gasa, S. Characterization of Novel Mono-O-Acetylated GM3s Containing 9-O-Acetyl Sialic Acid and 6-O-Acetyl Galactose in Equine Erythrocytes. Glycoconj. J. 1996, 13, 225–233. [Google Scholar] [CrossRef]
- Inagaki, M.; Shiizaki, M.; Hiwatashi, T.; Miyamoto, T.; Higuchi, R. Constituents of Crinoidea. 5. Isolation and Structure of a New Glycosyl Inositolphosphoceramide-Type Ganglioside from the Feather Star Comanthina Schlegeli. Chem. Pharm. Bull. 2007, 55, 1649–1651. [Google Scholar] [CrossRef] [Green Version]
- Svennerholm, L. Gangliosides of Human Thyroid Gland. Biochim. Biophys. Acta 1985, 835, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Herlyn, M.; Thurin, J.; Balaban, G.; Bennicelli, J.L.; Herlyn, D.; Elder, D.E.; Bondi, E.; Guerry, D.; Nowell, P.; Clark, W.H. Characteristics of Cultured Human Melanocytes Isolated from Different Stages of Tumor Progression. Cancer Res. 1985, 45, 5670–5676. [Google Scholar] [PubMed]
- Thurin, J.; Herlyn, M.; Hindsgaul, O.; Strömberg, N.; Karlsson, K.A.; Elder, D.; Steplewski, Z.; Koprowski, H. Proton NMR and Fast-Atom Bombardment Mass Spectrometry Analysis of the Melanoma-Associated Ganglioside 9-O-Acetyl-GD3. J. Biol. Chem. 1985, 260, 14556–14563. [Google Scholar] [CrossRef] [PubMed]
- Ostrander, G.K.; Bozlee, M.; Fukuda, M.; Dell, A.; Thomas-Oates, J.E.; Levery, S.B.; Eaton, H.L.; Hakomori, S.; Holmes, E.H. Isolation and Characterization of the Major Glycosphingolipids from the Liver of the Rainbow Trout (Oncorhynchus mykiss): Identification of an Abundant Source of 9-O-Acetyl GD3. Arch. Biochem. Biophys. 1991, 284, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kitajima, K.; Inoue, S.; Muto, Y.; Kasama, T.; Handa, S.; Inoue, Y. Structure of Novel Gangliosides, Deaminated Neuraminic Acid (KDN)-Containing Glycosphingolipids, Isolated from Rainbow Trout Ovarian Fluid. Biochemistry 1993, 32, 9221–9229. [Google Scholar] [CrossRef]
- Siebert, H.C.; von der Lieth, C.W.; Dong, X.; Reuter, G.; Schauer, R.; Gabius, H.J.; Vliegenthart, J.F. Molecular Dynamics-Derived Conformation and Intramolecular Interaction Analysis of the N-Acetyl-9-O-Acetylneuraminic Acid-Containing Ganglioside GD1a and NMR-Based Analysis of Its Binding to a Human Polyclonal Immunoglobulin G Fraction with Selectivity for O-Acetylated Sialic Acids. Glycobiology 1996, 6, 561–572. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Battistel, M.D.; Reeves, H.; Oh, L.; Yu, H.; Chen, X.; Wang, L.-P.; Freedberg, D.I. A Combined NMR, MD and DFT Conformational Analysis of 9-O-Acetyl Sialic Acid-Containing GM3 Ganglioside Glycan and Its 9-N-Acetyl Mimic. Glycobiology 2020, 30, 787–801. [Google Scholar] [CrossRef]
- Zhu, J.; Li, Y.T.; Li, S.C.; Cole, R.B. Structural Characterization of Gangliosides Isolated from Mullet Milt Using Electrospray Ionization-Tandem Mass Spectrometry. Glycobiology 1999, 9, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.X.; Chammas, R.; Varki, A. Linkage-Specific Action of Endogenous Sialic Acid O-Acetyltransferase in Chinese Hamster Ovary Cells. J. Biol. Chem. 1996, 271, 15130–15138. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.X.; Chammas, R.; Varki, A. Induction of Sialic Acid 9-O-Acetylation by Diverse Gene Products: Implications for the Expression Cloning of Sialic Acid O-Acetyltransferases. Glycobiology 1998, 8, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Corfield, A.P.; Sander-Wewer, M.; Veh, R.W.; Wember, M.; Schauer, R. The Action of Sialidases on Substrates Containing O-Acetylsialic Acids. Biol. Chem. Hoppe Seyler 1986, 367, 433–439. [Google Scholar] [CrossRef]
- Ritter, G.; Boosfeld, E.; Markstein, E.; Yu, R.K.; Ren, S.L.; Stallcup, W.B.; Oettgen, H.F.; Old, L.J.; Livingston, P.O. Biochemical and Serological Characteristics of Natural 9-O-Acetyl GD3 from Human Melanoma and Bovine Buttermilk and Chemically O-Acetylated GD3. Cancer Res. 1990, 50, 1403–1410. [Google Scholar]
- Manzi, A.E.; Sjoberg, E.R.; Diaz, S.; Varki, A. Biosynthesis and Turnover of O-Acetyl and N-Acetyl Groups in the Gangliosides of Human Melanoma Cells. J. Biol. Chem. 1990, 265, 13091–13103. [Google Scholar] [CrossRef]
- Sjoberg, E.R.; Varki, A. Kinetic and Spatial Interrelationships between Ganglioside Glycosyltransferases and O-Acetyltransferase(s) in Human Melanoma Cells. J. Biol. Chem. 1993, 268, 10185–10196. [Google Scholar] [CrossRef]
- Bora, R.S.; Kanamori, A.; Hirabayashi, Y. Cloning and Characterization of a Putative Mouse Acetyl-CoA Transporter CDNA. Gene 1999, 238, 455–462. [Google Scholar] [CrossRef]
- Satake, H.; Chen, H.Y.; Varki, A. Genes Modulated by Expression of GD3 Synthase in Chinese Hamster Ovary Cells. Evidence That the Tis21 Gene Is Involved in the Induction of GD3 9-O-Acetylation. J. Biol. Chem. 2003, 278, 7942–7948. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Challa, A.K.; Varki, A. 9-O-Acetylation of Exogenously Added Ganglioside GD3. The GD3 Molecule Induces Its Own O-Acetylation Machinery. J. Biol. Chem. 2006, 281, 7825–7833. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, K.; Aixinjueluo, W.; Kasama, T.; Ohkawa, Y.; Yoshihara, M.; Ohmi, Y.; Tajima, O.; Suzumura, A.; Kittaka, D.; Furukawa, K. Disruption of GM2/GD2 Synthase Gene Resulted in Overt Expression of 9-O-Acetyl GD3 Irrespective of Tis21. J. Neurochem. 2008, 105, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Kniep, B.; Kniep, E.; Ozkucur, N.; Barz, S.; Bachmann, M.; Malisan, F.; Testi, R.; Rieber, E.P. 9-O-Acetyl GD3 Protects Tumor Cells from Apoptosis. Int. J. Cancer 2006, 119, 67–73. [Google Scholar] [CrossRef]
- Mandal, C.; Srinivasan, G.V.; Chowdhury, S.; Chandra, S.; Mandal, C.; Schauer, R.; Mandal, C. High Level of Sialate-O-Acetyltransferase Activity in Lymphoblasts of Childhood Acute Lymphoblastic Leukaemia (ALL): Enzyme Characterization and Correlation with Disease Status. Glycoconj. J. 2009, 26, 57–73. [Google Scholar] [CrossRef]
- Ehara, T.; Kameyama, A.; Yamada, Y.; Ishida, H.; Kiso, M.; Hasegawa, A. Total Synthesis of VIM-2 Ganglioside Isolated from Human Chronic Myelogenous Leukemia Cells. Carbohydr. Res. 1996, 281, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Coers, W.; Reivinen, J.; Miettinen, A.; Huitema, S.; Vos, J.T.; Salant, D.J.; Weening, J.J. Characterization of a Rat Glomerular Visceral Epithelial Cell Line. Exp. Nephrol. 1996, 4, 184–192. [Google Scholar] [PubMed]
- Skov, L.; Chan, L.S.; Fox, D.A.; Larsen, J.K.; Voorhees, J.J.; Cooper, K.D.; Baadsgaard, O. Lesional Psoriatic T Cells Contain the Capacity to Induce a T Cell Activation Molecule CDw60 on Normal Keratinocytes. Am. J. Pathol. 1997, 150, 675–683. [Google Scholar] [PubMed]
- Aguilar, R.P.; Genta, S.; Sánchez, S. Renal Gangliosides Are Involved in Lead Intoxication. J. Appl. Toxicol. 2008, 28, 122–131. [Google Scholar] [CrossRef]
- Wipfler, D.; Srinivasan, G.V.; Sadick, H.; Kniep, B.; Arming, S.; Willhauck-Fleckenstein, M.; Vlasak, R.; Schauer, R.; Schwartz-Albiez, R. Differentially Regulated Expression of 9-O-Acetyl GD3 (CD60b) and 7-O-Acetyl-GD3 (CD60c) during Differentiation and Maturation of Human T and B Lymphocytes. Glycobiology 2011, 21, 1161–1172. [Google Scholar] [CrossRef] [Green Version]
- Chammas, R.; Sonnenburg, J.L.; Watson, N.E.; Tai, T.; Farquhar, M.G.; Varki, N.M.; Varki, A. De-N-Acetyl-Gangliosides in Humans: Unusual Subcellular Distribution of a Novel Tumor Antigen. Cancer Res. 1999, 59, 1337–1346. [Google Scholar]
- Schwarz, A.; Futerman, A.H. Determination of the Localization of Gangliosides Using Anti-Ganglioside Antibodies: Comparison of Fixation Methods. J. Histochem. Cytochem. 1997, 45, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Harms, G.; Reuter, G.; Corfield, A.P.; Schauer, R. Binding Specificity of Influenza C-Virus to Variably O-Acetylated Glycoconjugates and Its Use for Histochemical Detection of N-Acetyl-9-O-Acetylneuraminic Acid in Mammalian Tissues. Glycoconj. J. 1996, 13, 621–630. [Google Scholar] [CrossRef]
- Hubl, U.; Ishida, H.; Kiso, M.; Hasegawa, A.; Schauer, R. Studies on the Specificity and Sensitivity of the Influenza C Virus Binding Assay for 9-O-Acetylated Sialic Acids and Its Application to Human Melanomas. J. Biochem. 2000, 127, 1021–1031. [Google Scholar] [CrossRef]
- Krishna, M.; Varki, A. 9-O-Acetylation of Sialomucins: A Novel Marker of Murine CD4 T Cells That Is Regulated during Maturation and Activation. J. Exp. Med. 1997, 185, 1997–2013. [Google Scholar] [CrossRef] [Green Version]
- Gout, E.; Garlatti, V.; Smith, D.F.; Lacroix, M.; Dumestre-Pérard, C.; Lunardi, T.; Martin, L.; Cesbron, J.-Y.; Arlaud, G.J.; Gaboriaud, C.; et al. Carbohydrate Recognition Properties of Human Ficolins: Glycan Array Screening Reveals the Sialic Acid Binding Specificity of M-Ficolin. J. Biol. Chem. 2010, 285, 6612–6622. [Google Scholar] [CrossRef] [Green Version]
- Mandal, C.; Mandal, C.; Chandra, S.; Schauer, R.; Mandal, C. Regulation of O-Acetylation of Sialic Acids by Sialate-O-Acetyltransferase and Sialate-O-Acetylesterase Activities in Childhood Acute Lymphoblastic Leukemia. Glycobiology 2012, 22, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Draper, J.S.; Pigott, C.; Thomson, J.A.; Andrews, P.W. Surface Antigens of Human Embryonic Stem Cells: Changes upon Differentiation in Culture. J. Anat. 2002, 200, 249–258. [Google Scholar] [CrossRef]
- Azevedo-Pereira, R.L.; Morrot, A.; Machado, G.S.; Paredes, B.D.; de Carvalho Rodrigues, D.; de Carvalho, A.C.C.; Mendez-Otero, R. Expression of Ganglioside 9-O Acetyl GD3 in Undifferentiated Embryonic Stem Cells. Cell Biol. Int. 2015, 39, 121–127. [Google Scholar] [CrossRef]
- Fenderson, B.A.; Andrews, P.W.; Nudelman, E.; Clausen, H.; Hakomori, S. Glycolipid Core Structure Switching from Globo- to Lacto- and Ganglio-Series during Retinoic Acid-Induced Differentiation of TERA-2-Derived Human Embryonal Carcinoma Cells. Dev. Biol. 1987, 122, 21–34. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Taga, T.; Nakamura, K.; Ariga, T.; Yu, R.K. Characterization of Glycoconjugate Antigens in Mouse Embryonic Neural Precursor Cells. J. Neurochem. 2005, 95, 1311–1320. [Google Scholar] [CrossRef]
- Blum, A.S.; Barnstable, C.J. O-Acetylation of a Cell-Surface Carbohydrate Creates Discrete Molecular Patterns during Neural Development. Proc. Natl. Acad. Sci. USA 1987, 84, 8716–8720. [Google Scholar] [CrossRef] [Green Version]
- Sparrow, J.R.; Barnstable, C.J. A Gradient Molecule in Developing Rat Retina: Expression of 9-O-Acetyl GD3 in Relation to Cell Type, Developmental Age, and GD3 Ganglioside. J. Neurosci. Res. 1988, 21, 398–409. [Google Scholar] [CrossRef]
- Mendez-Otero, R.; Santiago, M.F. Functional Role of a Specific Ganglioside in Neuronal Migration and Neurite Outgrowth. Braz. J. Med. Biol. Res. 2003, 36, 1003–1013. [Google Scholar] [CrossRef] [Green Version]
- Drazba, J.; Pierce, M.; Lemmon, V. Studies of the Developing Chick Retina Using Monoclonal Antibody 8A2 That Recognizes a Novel Set of Gangliosides. Dev. Biol. 1991, 145, 154–163. [Google Scholar] [CrossRef]
- Hirabayashi, Y.; Hirota, M.; Suzuki, Y.; Matsumoto, M.; Obata, K.; Ando, S. Developmentally Expressed O-Acetyl Ganglioside GT3 in Fetal Rat Cerebral Cortex. Neurosci. Lett. 1989, 106, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ji, L.; Kurono, S.; Fujita, S.C.; Furuya, S.; Hirabayashi, Y. Developmentally Regulated O-Acetylated Sialoglycans in the Central Nervous System Revealed by a New Monoclonal Antibody 493D4 Recognizing a Wide Range of O-Acetylated Glycoconjugates. Glycoconj. J. 1997, 14, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Otero, R.; Constantine-Paton, M. Granule Cell Induction of 9-O-Acetyl Gangliosides on Cerebellar Glia in Microcultures. Dev. Biol. 1990, 138, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Otero, R.; Ramon-Cueto, A. Expression of 9-O-Acetylated Gangliosides during Development of the Rat Olfactory System. Neuroreport 1994, 5, 1755–1759. [Google Scholar] [CrossRef]
- Mello, L.E.; Mendez-Otero, R. Expression of 9-O-Acetylated Gangliosides in the Rat Hippocampus. Neurosci. Lett. 1996, 213, 17–20. [Google Scholar] [CrossRef]
- Mendez-Otero, R.; Cavalcante, L.A. Expression of 9-O-Acetylated Gangliosides Is Correlated with Tangential Cell Migration in the Rat Brain. Neurosci. Lett. 1996, 204, 97–100. [Google Scholar] [CrossRef]
- Dubois, C.; Manuguerra, J.C.; Hauttecoeur, B.; Maze, J. Monoclonal Antibody A2B5, Which Detects Cell Surface Antigens, Binds to Ganglioside GT3 (II3 (NeuAc)3LacCer) and to Its 9-O-Acetylated Derivative. J. Biol. Chem. 1990, 265, 2797–2803. [Google Scholar] [CrossRef]
- Mendez-Otero, R.; Friedman, J.E. Role of Acetylated Gangliosides on Neurite Extension. Eur. J. Cell Biol. 1996, 71, 192–198. [Google Scholar]
- Araujo, H.; Menezes, M.; Mendez-Otero, R. Blockage of 9-O-Acetyl Gangliosides Induces Microtubule Depolymerization in Growth Cones and Neurites. Eur. J. Cell Biol. 1997, 72, 202–213. [Google Scholar]
- Negreiros, E.M.A.; Leão, A.C.M.; Santiago, M.F.; Mendez-Otero, R. Localization of Ganglioside 9-O-Acetyl GD3 in Point Contacts of Neuronal Growth Cones. J. Neurobiol. 2003, 57, 31–37. [Google Scholar] [CrossRef]
- Varki, A.; Hooshmand, F.; Diaz, S.; Varki, N.M.; Hedrick, S.M. Developmental Abnormalities in Transgenic Mice Expressing a Sialic Acid-Specific 9-O-Acetylesterase. Cell 1991, 65, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, K.; Yamamoto, A.; Furukawa, K.; Yamashiro, S.; Shin, M.; Okada, M.; Fukumoto, S.; Haraguchi, M.; Takeda, N.; Fujimura, K.; et al. Mice with Disrupted GM2/GD2 Synthase Gene Lack Complex Gangliosides but Exhibit Only Subtle Defects in Their Nervous System. Proc. Natl. Acad. Sci. USA 1996, 93, 10662–10667. [Google Scholar] [CrossRef] [Green Version]
- Gubert, F.; Zaverucha-do-Valle, C.; Furtado, M.; Pimentel-Coelho, P.M.; Mortari, N.; Leão, A.C.M.; Hayashi, E.A.; Nobrega, A.; Mendez-Otero, R.; Santiago, M.F. CD60b: Enriching Neural Stem/Progenitor Cells from Rat Development into Adulthood. Stem Cells Int. 2017, 2017, 5759490. [Google Scholar] [CrossRef] [Green Version]
- Campos, F.S.O.; Piña-Rodrigues, F.M.; Reis, A.; Atella, G.C.; Mermelstein, C.S.; Allodi, S.; Cavalcante, L.A. Lipid Rafts from Olfactory Ensheathing Cells: Molecular Composition and Possible Roles. Cell. Mol. Neurobiol. 2021, 41, 525–536. [Google Scholar] [CrossRef]
- Santiago, M.F.; Berredo-Pinho, M.; Costa, M.R.; Gandra, M.; Cavalcante, L.A.; Mendez-Otero, R. Expression and Function of Ganglioside 9-O-Acetyl GD3 in Postmitotic Granule Cell Development. Mol. Cell. Neurosci. 2001, 17, 488–499. [Google Scholar] [CrossRef]
- Miyakoshi, L.M.; Mendez-Otero, R.; Hedin-Pereira, C. The 9-O-Acetyl GD3 Gangliosides Are Expressed by Migrating Chains of Subventricular Zone Neurons in Vitro. Braz. J. Med. Biol. Res. 2001, 34, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro-Resende, V.T.; Oliveira-Silva, A.; Ouverney-Brandão, S.; Santiago, M.F.; Hedin-Pereira, C.; Mendez-Otero, R. Ganglioside 9-O-Acetyl GD3 Expression Is Upregulated in the Regenerating Peripheral Nerve. Neuroscience 2007, 147, 97–105. [Google Scholar] [CrossRef]
- Ribeiro-Resende, V.T.; Araújo Gomes, T.; de Lima, S.; Nascimento-Lima, M.; Bargas-Rega, M.; Santiago, M.F.; Reis, R.A.d.M.; de Mello, F.G. Mice Lacking GD3 Synthase Display Morphological Abnormalities in the Sciatic Nerve and Neuronal Disturbances during Peripheral Nerve Regeneration. PLoS ONE 2014, 9, e108919. [Google Scholar] [CrossRef] [Green Version]
- Santiago, M.F.; Liour, S.S.; Mendez-Otero, R.; Yu, R.K. Glial-Guided Neuronal Migration in P19 Embryonal Carcinoma Stem Cell Aggregates. J. Neurosci. Res. 2005, 81, 9–20. [Google Scholar] [CrossRef]
- Santiago, M.F.; Scemes, E. Neuroblast Migration and P2Y(1) Receptor Mediated Calcium Signalling Depend on 9-O-Acetyl GD3 Ganglioside. ASN Neuro 2012, 4, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Santos-Silva, A.; Piña-Rodrigues, F.M.; Mermelstein, C.D.S.; Allodi, S.; Barradas, P.C.; Cavalcante, L.A. A Role for Gangliosides and Β1-Integrin in the Motility of Olfactory Ensheathing Glia. J. Anat. 2019, 235, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.F.; Costa, M.R.; Mendez-Otero, R. Immunoblockage of 9-O-Acetyl GD3 Ganglioside Arrests the in Vivo Migration of Cerebellar Granule Neurons. J. Neurosci. 2004, 24, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-R.; Liour, S.S.; Dasgupta, S.; Yu, R.K. Inhibition of Neuronal Migration by JONES Antibody Is Independent of 9-O-Acetyl GD3 in GD3-Synthase Knockout Mice. J. Neurosci. Res. 2007, 85, 1381–1390. [Google Scholar] [CrossRef]
- Miyakoshi, L.M.; Todeschini, A.R.; Mendez-Otero, R.; Hedin-Pereira, C. Role of the 9-O-Acetyl GD3 in Subventricular Zone Neuroblast Migration. Mol. Cell. Neurosci. 2012, 49, 240–249. [Google Scholar] [CrossRef]
- Leclerc, N.; Schwarting, G.A.; Herrup, K.; Hawkes, R.; Yamamoto, M. Compartmentation in Mammalian Cerebellum: Zebrin II and P-Path Antibodies Define Three Classes of Sagittally Organized Bands of Purkinje Cells. Proc. Natl. Acad. Sci. USA 1992, 89, 5006–5010. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.A.; Crandall, J.E.; Leclerc, N.; Yamamoto, M. Effects of Nervous Mutation on Purkinje Cell Compartments Defined by Zebrin II and 9-O-Acetylated Gangliosides Expression. Neurosci. Res. 1994, 19, 167–174. [Google Scholar] [CrossRef]
- Baader, S.L.; Vogel, M.W.; Sanlioglu, S.; Zhang, X.; Oberdick, J. Selective Disruption of “Late Onset” Sagittal Banding Patterns by Ectopic Expression of Engrailed-2 in Cerebellar Purkinje Cells. J. Neurosci. 1999, 19, 5370–5379. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Schwarting, G.A.; Crandall, J.E. Altered 9-O Acetylation of Disialogangliosides in Cerebellar Purkinje Cells of the Nervous Mutant Mouse. Brain Res. 1994, 662, 223–232. [Google Scholar] [CrossRef]
- Zimmer, G.; Suguri, T.; Reuter, G.; Yu, R.K.; Schauer, R.; Herrler, G. Modification of Sialic Acids by 9-O-Acetylation Is Detected in Human Leucocytes Using the Lectin Property of Influenza C Virus. Glycobiology 1994, 4, 343–349. [Google Scholar] [CrossRef]
- Kniep, B.; Flegel, W.A.; Northoff, H.; Rieber, E.P. CDw60 Glycolipid Antigens of Human Leukocytes: Structural Characterization and Cellular Distribution. Blood 1993, 82, 1776–1786. [Google Scholar] [CrossRef]
- Lünsdorf, H.; Kniep, E.; Kniep, B. Immunocytochemical Localization of CDw60 Antigens on Human Peripheral T Cells. Carbohydr. Res. 2000, 329, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Rieber, E.P.; Rank, G. CDw60: A Marker for Human CD8+ T Helper Cells. J. Exp. Med. 1994, 179, 1385–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vater, M.; Kniep, B.; Gross, H.J.; Claus, C.; Dippold, W.; Schwartz-Albiez, R. The 9-O-Acetylated Disialosyl Carbohydrate Sequence of CDw60 Is a Marker on Activated Human B Lymphocytes. Immunol. Lett. 1997, 59, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Kniep, B.; Claus, C.; Peter-Katalinic, J.; Monner, D.A.; Dippold, W.; Nimtz, M. 7-O-Acetyl-GD3 in Human T-Lymphocytes Is Detected by a Specific T-Cell-Activating Monoclonal Antibody. J. Biol. Chem. 1995, 270, 30173–30180. [Google Scholar] [CrossRef] [Green Version]
- Reivinen, J.; Holthöfer, H.; Miettinen, A. Tyrosine Phosphorylation of P72syk Induced by Anti-9-O-Acetyl GD3 Antibodies in Human Peripheral Blood Mononuclear Cells. Scand. J. Immunol. 1998, 48, 615–622. [Google Scholar] [CrossRef]
- Erdmann, M.; Wipfler, D.; Merling, A.; Cao, Y.; Claus, C.; Kniep, B.; Sadick, H.; Bergler, W.; Vlasak, R.; Schwartz-Albiez, R. Differential Surface Expression and Possible Function of 9-O- and 7-O-Acetylated GD3 (CD60 b and c) during Activation and Apoptosis of Human Tonsillar B and T Lymphocytes. Glycoconj. J. 2006, 23, 627–638. [Google Scholar] [CrossRef]
- Mukherjee, K.; Chowdhury, S.; Mondal, S.; Mandal, C.; Chandra, S.; Bhadra, R.K.; Mandal, C. 9-O-Acetylated GD3 Triggers Programmed Cell Death in Mature Erythrocytes. Biochem. Biophys. Res. Commun. 2007, 362, 651–657. [Google Scholar] [CrossRef]
- Mukherjee, K.; Chowdhury, S.; Mondal, S.; Mandal, C.; Chandra, S.; Mandal, C. 9-O-Acetyl GD3 in Lymphoid and Erythroid Cells. Adv. Exp. Med. Biol. 2011, 705, 317–334. [Google Scholar] [CrossRef]
- Simons, M.; Schwarz, K.; Kriz, W.; Miettinen, A.; Reiser, J.; Mundel, P.; Holthöfer, H. Involvement of Lipid Rafts in Nephrin Phosphorylation and Organization of the Glomerular Slit Diaphragm. Am. J. Pathol. 2001, 159, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Kohla, G.; Stockfleth, E.; Schauer, R. Gangliosides with O-Acetylated Sialic Acids in Tumors of Neuroectodermal Origin. Neurochem. Res. 2002, 27, 583–592. [Google Scholar] [CrossRef]
- Cheresh, D.A.; Varki, A.P.; Varki, N.M.; Stallcup, W.B.; Levine, J.; Reisfeld, R.A. A Monoclonal Antibody Recognizes an O-Acylated Sialic Acid in a Human Melanoma-Associated Ganglioside. J. Biol. Chem. 1984, 259, 7453–7459. [Google Scholar] [CrossRef]
- Herlyn, M.; Rodeck, U.; Mancianti, M.; Cardillo, F.M.; Lang, A.; Ross, A.H.; Jambrosic, J.; Koprowski, H. Expression of Melanoma-Associated Antigens in Rapidly Dividing Human Melanocytes in Culture. Cancer Res. 1987, 47, 3057–3061. [Google Scholar]
- Berd, D.; Herlyn, M.; Koprowski, H.; Mastrangelo, M.J. Flow Cytometric Determination of the Frequency and Heterogeneity of Expression of Human Melanoma-Associated Antigens. Cancer Res. 1989, 49, 6840–6844. [Google Scholar]
- Hamilton, W.B.; Helling, F.; Lloyd, K.O.; Livingston, P.O. Ganglioside Expression on Human Malignant Melanoma Assessed by Quantitative Immune Thin-Layer Chromatography. Int. J. Cancer 1993, 53, 566–573. [Google Scholar] [CrossRef]
- Sjoberg, E.R.; Manzi, A.E.; Khoo, K.H.; Dell, A.; Varki, A. Structural and Immunological Characterization of O-Acetylated GD2. Evidence That GD2 Is an Acceptor for Ganglioside O-Acetyltransferase in Human Melanoma Cells. J. Biol. Chem. 1992, 267, 16200–16211. [Google Scholar] [CrossRef]
- Ren, S.L.; Slominski, A.; Yu, R.K. Glycosphingolipids in Bomirski Transplantable Melanomas in Hamsters. Cancer Res. 1989, 49, 7051–7056. [Google Scholar]
- Kageshita, T.; Nakamura, T.; Yamada, M.; Kuriya, N.; Arao, T.; Ferrone, S. Differential Expression of Melanoma Associated Antigens in Acral Lentiginous Melanoma and in Nodular Melanoma Lesions. Cancer Res. 1991, 51, 1726–1732. [Google Scholar]
- Natali, P.G.; Bigotti, A.; Nicotra, M.R.; Nardi, R.M.; Delovu, A.; Segatto, O.; Ferrone, S. Analysis of the Antigenic Profile of Uveal Melanoma Lesions with Anti-Cutaneous Melanoma-Associated Antigen and Anti-HLA Monoclonal Antibodies. Cancer Res. 1989, 49, 1269–1274. [Google Scholar]
- Kanda, S.; Cochran, A.J.; Lee, W.R.; Morton, D.L.; Irie, R.F. Variations in the Ganglioside Profile of Uveal Melanoma Correlate with Cytologic Heterogeneity. Int. J. Cancer 1992, 52, 682–687. [Google Scholar] [CrossRef]
- Sela, B.A.; Iliopoulos, D.; Guerry, D.; Herlyn, D.; Koprowski, H. Levels of Disialogangliosides in Sera of Melanoma Patients Monitored by Sensitive Thin-Layer Chromatography and Immunostaining. J. Natl. Cancer Inst. 1989, 81, 1489–1492. [Google Scholar] [CrossRef]
- Ren, S.; Ariga, T.; Scarsdale, J.N.; Zhang, Y.; Slominski, A.; Livingston, P.O.; Ritter, G.; Kushi, Y.; Yu, R.K. Characterization of a Hamster Melanoma-Associated Ganglioside Antigen as 7-O-Acetylated Disialoganglioside GD3. J. Lipid Res. 1993, 34, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Popa, I.; Pons, A.; Mariller, C.; Tai, T.; Zanetta, J.-P.; Thomas, L.; Portoukalian, J. Purification and Structural Characterization of De-N-Acetylated Form of GD3 Ganglioside Present in Human Melanoma Tumors. Glycobiology 2007, 17, 367–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.X.; Chammas, R.; Varki, A. Regulation of Sialic Acid 9-O-Acetylation during the Growth and Differentiation of Murine Erythroleukemia Cells. J. Biol. Chem. 1996, 271, 31517–31525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, K.; Chava, A.K.; Mandal, C.; Dey, S.N.; Kniep, B.; Chandra, S.; Mandal, C. O-Acetylation of GD3 Prevents Its Apoptotic Effect and Promotes Survival of Lymphoblasts in Childhood Acute Lymphoblastic Leukaemia. J. Cell. Biochem. 2008, 105, 724–734. [Google Scholar] [CrossRef]
- Scala, E.; Abeni, D.; Pomponi, D.; Narducci, M.G.; Lombardo, G.A.; Mari, A.; Frontani, M.; Picchio, M.C.; Pilla, M.A.; Caprini, E.; et al. The Role of 9-O-Acetylated Ganglioside D3 (CD60) and {alpha}4{beta}1 (CD49d) Expression in Predicting the Survival of Patients with Sezary Syndrome. Haematologica 2010, 95, 1905–1912. [Google Scholar] [CrossRef] [Green Version]
- Paller, A.S.; Arnsmeier, S.L.; Robinson, J.K.; Bremer, E.G. Alteration in Keratinocyte Ganglioside Content in Basal Cell Carcinomas. J. Investig. Dermatol. 1992, 98, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Fahr, C. Detection of Sialic Acids and Gangliosides with Special Reference to 9-O-Acetylated Species in Basaliomas and Normal Human Skin. J. Investig. Dermatol. 2001, 116, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, R.; Allman, R.; Mason, M.D. Ganglioside Expression in Lung Cancer Cell Lines. Lung Cancer 1997, 18, 21–33. [Google Scholar] [CrossRef]
- Gocht, A.; Rutter, G.; Kniep, B. Changed Expression of 9-O-Acetyl GD3 (CDw60) in Benign and Atypical Proliferative Lesions and Carcinomas of the Human Breast. Histochem. Cell Biol. 1998, 110, 217–229. [Google Scholar] [CrossRef]
- Cavdarli, S.; Dewald, J.H.; Yamakawa, N.; Guérardel, Y.; Terme, M.; Le Doussal, J.-M.; Delannoy, P.; Groux-Degroote, S. Identification of 9-O-Acetyl-N-Acetylneuraminic Acid (Neu5,9Ac2) as Main O-Acetylated Sialic Acid Species of GD2 in Breast Cancer Cells. Glycoconj. J. 2019, 36, 79–90. [Google Scholar] [CrossRef]
- Cavdarli, S.; Schröter, L.; Albers, M.; Baumann, A.-M.; Vicogne, D.; Le Doussal, J.-M.; Mühlenhoff, M.; Delannoy, P.; Groux-Degroote, S. Role of Sialyl-O-Acetyltransferase CASD1 on GD2 Ganglioside O-Acetylation in Breast Cancer Cells. Cells 2021, 10, 1468. [Google Scholar] [CrossRef]
- Ariga, T.; Suetake, K.; Nakane, M.; Kubota, M.; Usuki, S.; Kawashima, I.; Yu, R.K. Glycosphingolipid Antigens in Neural Tumor Cell Lines and Anti-Glycosphingolipid Antibodies in Sera of Patients with Neural Tumors. Neurosignals 2008, 16, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Birks, S.M.; Danquah, J.O.; King, L.; Vlasak, R.; Gorecki, D.C.; Pilkington, G.J. Targeting the GD3 Acetylation Pathway Selectively Induces Apoptosis in Glioblastoma. Neuro Oncol. 2011, 13, 950–960. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, Y.; Zheng, Q.; Li, J. Analysis of O-Acetylated Sialic Acids in Dried Blood Spots. Anal. Chem. 2019, 91, 2744–2751. [Google Scholar] [CrossRef]
- Malisan, F.; Franchi, L.; Tomassini, B.; Ventura, N.; Condò, I.; Rippo, M.R.; Rufini, A.; Liberati, L.; Nachtigall, C.; Kniep, B.; et al. Acetylation Suppresses the Proapoptotic Activity of GD3 Ganglioside. J. Exp. Med. 2002, 196, 1535–1541. [Google Scholar] [CrossRef] [Green Version]
- Giussani, P.; Tringali, C.; Riboni, L.; Viani, P.; Venerando, B. Sphingolipids: Key Regulators of Apoptosis and Pivotal Players in Cancer Drug Resistance. Int. J. Mol. Sci. 2014, 15, 4356–4392. [Google Scholar] [CrossRef] [Green Version]
- Romero-Ramírez, L.; Nieto-Sampedro, M. Inhibiting Human Astrocytoma Growth: Structure-Activity Relationships in Neurostatin Related Glycolipids. J. Med. Chem. 2004, 47, 4983–4984. [Google Scholar] [CrossRef]
- Valle-Argos, B.; Gómez-Nicola, D.; Nieto-Sampedro, M. Synthesis and Characterization of Neurostatin-Related Compounds with High Inhibitory Activity of Glioma Growth. Eur. J. Med. Chem. 2010, 45, 2034–2043. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Helling, F.; Lloyd, K.O.; Livingston, P.O. Increased Tumor Cell Reactivity and Complement-Dependent Cytotoxicity with Mixtures of Monoclonal Antibodies against Different Gangliosides. Cancer Immunol. Immunother. 1995, 40, 88–94. [Google Scholar] [CrossRef]
- Zhang, S.; Cordon-Cardo, C.; Zhang, H.S.; Reuter, V.E.; Adluri, S.; Hamilton, W.B.; Lloyd, K.O.; Livingston, P.O. Selection of Tumor Antigens as Targets for Immune Attack Using Immunohistochemistry: I. Focus on Gangliosides. Int. J. Cancer 1997, 73, 42–49. [Google Scholar] [CrossRef]
- Ritter, G.; Ritter-Boosfeld, E.; Adluri, R.; Calves, M.; Ren, S.; Yu, R.K.; Oettgen, H.F.; Old, L.J.; Livingston, P.O. Analysis of the Antibody Response to Immunization with Purified O-Acetyl GD3 Gangliosides in Patients with Malignant Melanoma. Int. J. Cancer 1995, 62, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Dumontet, C.; Rebbaa, A.; Portoukalian, J. Very Low Density Lipoproteins and Interleukin 2 Enhance the Immunogenicity of 9-O-Acetyl-GD3 Ganglioside in BALB/c Mice. J. Immunol. Methods 1997, 206, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ye, J.; DeLaitsch, A.T.; Rashidijahanabad, Z.; Lang, S.; Kakeshpour, T.; Zhao, Y.; Ramadan, S.; Saavedra, P.V.; Yuzbasiyan-Gurkan, V.; et al. Chemoenzymatic Synthesis of 9NHAc-GD2 Antigen to Overcome the Hydrolytic Instability of O-Acetylated-GD2 for Anticancer Conjugate Vaccine Development. Angew. Chem. Int. Ed. Engl. 2021, 60, 24179–24188. [Google Scholar] [CrossRef] [PubMed]
- Schultze, B.; Zimmer, G.; Herrler, G. Virus Entry into a Polarized Epithelial Cell Line (MDCK): Similarities and Dissimilarities between Influenza C Virus and Bovine Coronavirus. J. Gen. Virol. 1996, 77 Pt 10, 2507–2514. [Google Scholar] [CrossRef]
- Li, Z.; Lang, Y.; Liu, L.; Bunyatov, M.I.; Sarmiento, A.I.; de Groot, R.J.; Boons, G.-J. Synthetic O-Acetylated Sialosides Facilitate Functional Receptor Identification for Human Respiratory Viruses. Nat. Chem. 2021, 13, 496–503. [Google Scholar] [CrossRef]
- Zimmer, G.; Reuter, G.; Schauer, R. Use of Influenza C Virus for Detection of 9-O-Acetylated Sialic Acids on Immobilized Glycoconjugates by Esterase Activity. Eur. J. Biochem. 1992, 204, 209–215. [Google Scholar] [CrossRef]
- Manuguerra, J.C.; DuBois, C.; Hannoun, C. Analytical Detection of 9(4)-O-Acetylated Sialoglycoproteins and Gangliosides Using Influenza C Virus. Anal. Biochem. 1991, 194, 425–432. [Google Scholar] [CrossRef]
- Ribeiro-Resende, V.T.; Ribeiro-Guimarães, M.L.; Lemes, R.M.R.; Nascimento, I.C.; Alves, L.; Mendez-Otero, R.; Pessolani, M.C.V.; Lara, F.A. Involvement of 9-O-Acetyl GD3 Ganglioside in Mycobacterium Leprae Infection of Schwann Cells. J. Biol. Chem. 2010, 285, 34086–34096. [Google Scholar] [CrossRef] [Green Version]
- Hitoshi, S.; Kusunoki, S.; Kon, K.; Chiba, A.; Waki, H.; Ando, S.; Kanazawa, I. A Novel Ganglioside, 9-O-Acetyl GD1b, Is Recognized by Serum Antibodies in Guillain-Barré Syndrome. J. Neuroimmunol. 1996, 66, 95–101. [Google Scholar] [CrossRef]
- Fox, D.A.; He, X.; Abe, A.; Hollander, T.; Li, L.L.; Kan, L.; Friedman, A.W.; Shimizu, Y.; Shayman, J.A.; Kozarsky, K. The T Lymphocyte Structure CD60 Contains a Sialylated Carbohydrate Epitope That Is Expressed on Both Gangliosides and Glycoproteins. Immunol. Investig. 2001, 30, 67–85. [Google Scholar] [CrossRef]
- Fantini, J.; Azzaz, F.; Chahinian, H.; Yahi, N. Electrostatic Surface Potential as a Key Parameter in Virus Transmission and Evolution: How to Manage Future Virus Pandemics in the Post-COVID-19 Era. Viruses 2023, 15, 284. [Google Scholar] [CrossRef]
- Sun, X.-L. The Role of Cell Surface Sialic Acids for SARS-CoV-2 Infection. Glycobiology 2021, 31, 1245–1253. [Google Scholar] [CrossRef]
- Oh, L.; Varki, A.; Chen, X.; Wang, L.-P. SARS-CoV-2 and MERS-CoV Spike Protein Binding Studies Support Stable Mimic of Bound 9-O-Acetylated Sialic Acids. Molecules 2022, 27, 5322. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Acronym | Sialic Acid Modification | Main Structural Features |
|---|---|---|
| GM | N/A | One sialic acid residue |
| GD | N/A | Two sialic acid residues |
| GT | N/A | Three sialic acid residues |
| GM1 | N/A | One sialic acid and four neutral sugar residues |
| GD1 | N/A | Two sialic acid and four neutral sugar residues |
| GT3 | N/A | Three sialic acid and two neutral sugar residues. All three sialic acid residues are linked to galactose residue in position 2 from the ceramide backbone. |
| 9-O-acGM3 | O-acetylated sialic acid | One sialic acid and two neutral sugar residues; O-acetylation on carbon 9 of one sialic acid. |
| 9-O-acGD3 | O-acetylated sialic acid | Two sialic acid and two neutral sugar residues; O-acetylation on carbon 9 of one sialic acid. |
| 7-O-acGD3 | O-acetylated sialic acid | Two sialic acid and two neutral sugar residues; O-acetylation on carbon 7 of one sialic acid residue. |
| 9-N-acGD2 | N-acetylated sialic acid | Two sialic acid and three neutral sugar residues; N-acetylation on carbon 9 of one sialic acid residue. |
| 9-O-acGD1a | O-acetylated sialic acid | Two sialic acid and four neutral sugar residues; O-acetylation on carbon 9 of one sialic acid residue. One sialic acid residue is linked to the galactose as second neutral sugar from the ceramide backbone. |
| 9-O-acGD1b | O-acetylated sialic acid | Two sialic acid and four neutral sugar residues; O-acetylation on carbon 9 of one sialic acid residue. The two sialic acid residues are linked to the galactose as the second neutral sugar from the ceramide backbone. |
| 9-O-acGT2 | O-acetylated sialic acid | Three sialic acid and three neutral sugar residues; O-acetylation on carbon 9 of one sialic acid residue. The three sialic acid residues are linked to the galactose as the second neutral sugar from the ceramide backbone. |
| 9-O-acGT3 | O-acetylated sialic acid | Three sialic acid and two neutral sugar residues; O-acetylation on carbon 9 of one sialic acid residue. The three sialic acid residues are linked to the galactose as the second neutral sugar from the ceramide backbone. |
| Embryogenesis | |||
|---|---|---|---|
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 1987 [55] | Ganglio-series replace globo-series when differentiation is induced by retinoic acid | NTERA-2 (human embryonic carcinoma line) | TLC + Antibody (ME-311): 9-O-acGD3 |
| 2005 [56] | 9-O-acGD3 presence in neuroepithelial precursor cells | Neuroepithelial precursor cells | FC + Antibody (D1.1): 9-O-acGD3 |
| 1988 [58] | 9-O-acGD3 rise between day E15 and postnatal day 2, and pronounced drop between day 2 and day 4 PN | Rat developing retinae | IF + Antibody (JONES): 9-O-acGD3 |
| 1991 [60] | Detection of 9-O-ac gangliosides in the optic fiber layer of central retina | Cultured cells from chicken embryo retinae | TLC/electron microscopy + Antibody (Mabs D1.1/JONES and 8A2): 9-O-acGD3 and unspecific gangliosides |
| 1989 [61] | 9-O-acGT3 increased in rat cerebral cortex at day 14 of gestation, then decreased and was absent in adult rats | Fetal rat cerebral cortex | TLC + Antibody (M6704): c-series gangliosides. |
| 1997 [62] | 9-O-acGD3 increased in rat cerebral cortex at day 14 of gestation, then decreased and was absent in adult rats | Fetal rat cerebral cortex | TLC + Antibody (493D4): O-acGD3, O-acLD1, O-acGD2 and O-acGD1b |
| 1990 [63] | Acetylated gangliosides associated with granule cell migration (neurons) and glial cells require some form of neuron-glia interaction to display acetylated gangliosides | Cultured cells from 2 to 6 day postnatal rat cerebellums | ICC + Antibody (JONES): JONES antigens |
| 1994 [64] | Acetylated gangliosides associated with the formation of mature olfactory bulb | Developing embryonic rat nervous system and postnatal rats | IHC + Antibody (JONES): JONES antigens |
| 1996 [65] | Acetylated gangliosides associated with the formation of hippocampus and rapid decrease after birth. | Embryonic, postnatal and adult rat hippocampus | IHC + Antibody (JONES): JONES antigens |
| 1996 [66] | 9-O-ac gangliosides are involved in tangential cell migration both in lateral ventricle and rostral subventricular zone, along the rostral migratory stream and in the olfactory bulb in developing animals and, at lower levels, in adulthood. | Embryonic, postnatal and adult rat brain | IHC + Antibody (JONES): JONES antigens |
| 1990 [67] | Monoclonal antibody A2B5 detects GT3, 9-O-acGT3 and other antigens. All A2B5-detected antigens decrease during chicken brain development | 10-day embryonic chicken brain | TLC + Antibody (A2B5): GT3 and 9-O-acGT3 |
| 1996 [68] | 9-O-ac gangliosides play a role in the extension of growth cones in neurites | Neurons of embryonic rat dorsal root ganglia explants grown on laminin substratum | IHC + Antibody (JONES): JONES antigens |
| 1997 [69] | 9-O-ac gangliosides regulate the microfilament and microtubular structure of neurites | Unavailable information | Unavailable information |
| 2003 [70] | 9-O-acGD3 localizes in contact points of neural growth cones and is associated with β-1-integrin and vinculin | Cultured neurites from dorsal root ganglia from embryonic rat | IHC + CM + Antibody (JONES): JONES antigens |
| 1991 [71] | The cleavage of 9-O-ac esters on sialic acids causes 2-cell stage arrest in murine embryogenesis | Transgenic mice with a loss of O-ac of Sialic Acid | N/A |
| 1991 [71] | Cleavage of 9-O-ac esters on sialic acids in retina and adrenal gland leads to impaired morphology and function in these organs (postnatal) | Transgenic mice with a loss of O-ac of sialic acid in adrenal gland and retina | IHC + Antibody(27A): 9-O-acGD3 |
| Postnatal nervous system | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 2008 [38] | The absence of GM2/GD2 in nervous tissue increases GM3 and GD3 (this also includes 9-O-acGD3) | GM2/GD2 synthase KO mice | TLC + Antibody (JONES and GMR2): 9-O-acGD3 |
| 1988 [58] | Dorsal–ventral gradient of 9-O-acGD3 in postnatal rat retina | Developing rat retina | IHC + ICC + Antibody (JONES and R24) |
| 1996 [66] | Dorsal–ventral gradient of 9-O-acGD3 in lateral ventricle rostral subventricular zone, along the rostral migratory stream and in the olfactory bulb at lower levels than in the developing nervous | Embryonic, postnatal, and adult rat brains | IHC + Antibody (JONES): JONES antigens |
| 1991 [60] | 9-O-ac gangliosides are not detected in the central optic fiber. In contrast, they remain in the inner and outer plexiform layer, and in the outer nuclear layer, | Adult chicken | TLC/electron microscopy + Antibody (Mabs D1.1/JONES and 8A2): 9-O-acGD3 and unspecific gangliosides |
| 1996 [65] | 9-O-ac ganglioside is absent in rat adult hippocampus | Adult rat | IHC + Antibody (JONES): JONES antigens |
| 2017 [73] | 9-O-acGD3 presence in subventricular zone from neural stem and progenitor cells in the adult | Postnatal Lister Hooded rats | IHC + Antibodies (CD60b and JONES): CD60b antigens |
| 1990 [63] | Acetylated gangliosides associated with granule cell migration (neurons) and glial cells require some form of neuron-glia interaction to be displayed | Cultured cells from 2 to 6 day postnatal rat cerebellums | ICC + Antibody (JONES): JONES antigens |
| 2001 [75] | Finding of 9-O-acGD3 in the contact sites of migrating granule cells and in radial glia; 9-O-acGD3 involvement in granule cell migration in the developing cerebellum | Postnatal rat cerebellum and rat cerebellar explants | IHC/IF/IEM + Antibody (JONES): JONES antigens. |
| 2001 [74] | The identification of 9-O-acGD3 in membrane rafts | Primary culture of olfactory ensheathing glia from rat | Membrane raft isolation. Dot blotting + Antibody (JONES) |
| 2001 [76] | 9-O-acGD3 may participate in neuronophilic and gliophilic migration | Culture explants of anterior subventricular zone (SVZ) of cerebral cortex from postnatal rats | CM + Antibody (JONES): JONES antigens Immunoblockage (JONES) |
| 2007 [77] | 9-O-acGD3 is re-expressed in neurons and glia cells involved in axonal regeneration | Sciatic nerve from adult rats and its explant culture | CM + Antibodies (mouse IgM monoclonal anti-9-O-acGD3 (Sigma) and JONES): 9-O-acGD3 |
| 2014 [78] | Defective axonal regeneration in GD3 synthase KO that can be rescued through administration of exogenous GD3 | Sciatic nerve from adult rats and its explant culture | N/A |
| 2005 [79] | Participation of 9-O-ac gangliosides in granule cell migration | Neuron-like cultured cells derived from P19 embryonal carcinoma stem cells | TLC/IF + Antibodies (Jones and D1.1): 9-O-acGD3. Blockage of migration (JONES) |
| 2012 [80] | Participation of 9-O-ac gangliosides in granule cell migration through a calcium-signaling mechanism involving PY2 receptors | Explant culture from mouse early postnatal cerebellum | IF + Antibody (JONES). Blockage of migration (JONES) |
| 2019 [81] | Antibody inhibition of olfactory ensheathing glia migration | Organotypical olfactory ensheathing cultures from rats | IF+ anti-9-O-acGD3 (mouse IgM monoclonal antibody; Sigma). Immunoblockage (JONES) |
| 2004 [82] | Inmunoblockage of neuronal migration by JONES antibody but not by A2B5 antibody | Cerebellar granule neurons from postnatal rats | CM + BrU. Immunoblockage (JONES) |
| 2007 [83] | Independence of the mice model in the inhibition of neuronal migration by JONES antibody + JONES-positive proteins raises questions on antibody specificity | Cerebella from wild-type and GD synthase KO mice | IHC, IF, TLC, WB + Antibodies (JONES, D1.1, or A2B5 (c-series gangliosides)) |
| 2012 [84] | Inhibition of neuronal migration by inmunoblocking with JONES antibody; 9-O-acGD3 role in cell–cell and cell–substrate interactions in neuroblast | Subventricular zone explants from rat brain | Videomicroscopy, IF, Immunoblockage (JONES) |
| 1992 [85] | Two subtypes of Purkinje cells contain 9-O-ac glycolipids | Adult mice cerebellum | IHC/TLC + Antibody (P-path): 9-O-ac glycolipids |
| 1994 [86] | Nervous mutation-surviving Purkinje cells in the cerebellum correspond to those positive for 9-O-ac gangliosides | Nervous mutation (nr/nr and nr/+) and wild-type (+/+) mice | IHC + Antibodies (P-path): 9-O-ac glycolipids (9-O- acGD3 and 9-O-acLDI) |
| 1999 [87] | Purkinje cell P-path antigens mark the late-onset sagittal banding patterns and they are En-2-sensitive | Postnatal wild-type and En-2 mutant mice | IHC + Antibodies (P-path): 9-O-ac glycolipids (9-O- acGD3 and 9-O-acGD1) |
| 1994 [88] | Nervous mutation-surviving Purkinje cells in the cerebellum correspond to those positive mainly for 9-O-acGD3 | Nervous mutation (nr/nr and nr/+) and wild type (+/+) mice | IHC/TLC + Antibody (P-path): 9-O-ac glycolipids (9-O- acGD3 and 9-O-acLDI) |
| Immune system | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 1994 [89] | Characterization of T lymphocyte CDw60 antigen as 9-O-acGD3 | Leukocytes from children’s tonsils and from healthy adult donors | TLC + influenza C virus incubation: 9-O-ac gangliosides |
| 1995 [90] | T lymphocytes (mostly CD4+) and granulocytes present high amounts of CD60 antigen, in contrast to low levels present in B cells, thymus cells and monocytes | Human leukocytes | TLC + Antibodies R24 do not detect 9-O-acGD3 but UM4D4 does (unspecific). Mass spectrometry |
| 2000 [91] | 25% of peripheral T cells present a surface localization of CD60, while roughly all T cells express intracellularly CD60 in Golgi vesicles | T lymphocytes | FC/IEM + Antibody (M-T32): CD60 antigen |
| 1994 [92] | CD8+ CD60+ subset of T cells (T helper CD8+) secretes more IL-4 and less interferon gamma than CD8+ CD60- T cells | T lymphocytes from healthy volunteer donors | FC + Ab mAb M-T32: CD60 antigen |
| 1997 [93] | CD60 is an activation marker of human B cells. Peripheral and tonsillar B cells become CD60+ when activated by phorbol esters | Peripheral blood lymphocytes from healthy donors and tonsillar B cells from children | FC/TLC + Antibodies (UM4D4, F6 and Z17): CD60 |
| 1997 [50] | TCR activation decreases 9-O-ac sialic acid at the surface of T cells, but due to decreased sialomucins, not necessarily to gangliosides | Mouse lymphocytes from either spleen or lymph nodes | Lipid extraction + ELISA (CHE-FcD): 9-O-ac sialic acid |
| 1998 [95] | Induction of Syk, phosphoinositide mobilization and cell proliferation in PBMC by treatment with a monoclonal antibody targeting 9-O-acGD3 | Human PBMC | TLC/FC/IEM + Antibodies (27A and R24): 9-O-acGD3 and GD3 respectively |
| 2006 [96] | CD60 antigen is subdivided into CD60a (GD3), CD60b (9-O-acetylated form), and CD60c (7-O-acetylated form) Anti-CD60b with IL-4 can costimulate B cells CD60b is present in Extrafollicular T cells and can be costimulated with antiCD60b and PHA | Human tonsillar lymphocytes | IHC/CM/FC + Antibodies (R24, UM4D4 and U5): GD3, 9-O-acGD3 and 7-O-acGD3 |
| CD60b is present in tonsillar B cells in the activated germinal center, colocalizing in lipid rafts with Syk and Lyn | |||
| 2011 [45] | Both T and B cells present CD60b staining in a patchy fashion as compared to the other forms of CD60 antigen CD4+ cells show the strongest, and CD8+ the weakest, presence of CD60b at the surface in thymocytes Subcellular distribution of 9-O-acGD3 is non-raft microdomains in T cells and raft microdomains in B cells | Human tonsillar lymphocytes | IHC/CM/FC + Antibodies (R24, UM4D4 and U5): GD3, 9-O-acGD3 and 7-O-acGD3 |
| Hematopoiesis | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 2007 [97] | 9-O-acGD3 is present in human bone marrow erythroid progenitors, is progressively lost during maturation, and becomes proapoptotic in mature erythrocytes | Bone marrow and peripheral blood erythrocytes from children with acute lymphoblastic leukemia and clinical remission | FC + Antibody (JONES): 9-O-acGD3 |
| Kidney | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 1996 [42] | Cultured podocytes contain 9-O-acGD3 and it immunoprecipitates with a non-characterized podocyte protein | Cultured podocyte line from rat glomerular explants | IF/IP + Antibodies (27A): 9-O-acGD3 |
| 2001 [99] | 9-O-acGD3 colocalizes in podocyte lipid rafts with nephrin at the slit diaphragm, a constituent of the glomerular filtration barrier | Rat kidneys and glomeruli | IHC/IP/IEM + Antibody (27A): 9-O-acGD3 |
| Cancer | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 2002 [100] | 9-O-acetylation of gangliosides as a marker of cell and tissue growth in cancer | Review article | Review article |
| 1984 [101] | Band comigrating with 9-O-ac gangliosides from melanoma cell lipid extracts | Rat (B49) and human (M14) melanoma cell lines | TLC + Antibody (D1.1): 9-O-acGD3 |
| 1985 [23] | 9-O-ac gangliosides detected in nevi and melanoma cells and also in lymphocytes in 30% of cases studied | 27 melanoma cell lines | FAB-MS + NMR/IHC + Antibody (ME 311) |
| 1987 [102], 1989 [103], 1993 [104] | 9-O-acGD3 considered as a melanoma antigen | 20 melanoma cell lines and 5 human tissues | TLC + Antibody (D1.l): 9-O-acGD3 |
| 1992 [105] | 9-O-acGD2 is a melanoma antigen | M21 melanoma cell line | FABS-MS/NMR/TLC + Antibodies(14.G2A): 9-O-acGD2 |
| 1989 [106] | 9-O-acGD3 increased in amelanotic, fast-growing stage, as compared with slow-growing, highly differentiated forms, suggesting a role in cell growth | Hamster melanoma cells: Ab amelanotic melanoma (fast-growing), Ma melanotic melanoma (slow-growing), and MI hypomelanotic melanoma (slow-growing) | Unavailable information |
| 1991 [107] | 9-O-acGD3 presence in nodular melanoma higher than in metastatic acral lentiginous melanoma | Primary and metastatic acral lentiginous melanoma and nodular melanoma lesions from patients | Unavailable information |
| 1989 [108], 1992 [109] | 9-O-acGD3 is not present in uveal melanoma | Surgically removed uveal melanoma lesion | ME311 [108], TLC [109]: 9-O-acGD3 |
| 2007 [112] | In human melanoma, a high presence of sphingosine C24:1 in both 9-O-acGD3 and GD3 | Human melanoma tumors | HPLC-GLC-MS/TLC: 9-O-acGD3, GD3 |
| 1996 [113] | 9-O-acGD3 is present in mouse erythroleukemia cells intracellularly | Murine erythroleukemia (MEL) cells | Ganglioside extraction + ELISA (CHE-FcD, 27A): 9-O-ac gangliosides, 9-O-acGD3 |
| 2008 [114] | Lymphoblasts from acute lymphoblastic leukemia patients have increased levels of 9-O-acGD3 and it accumulates in mitochondrial membrane Exogenous 9-O-acGD3 (but not GD3) prevents mitochondrial membrane depolarization, cytochrome C release and caspase activation in lymphoblasts | (MOLT-4) ALL cell line and PBMC from patient | IEM/TLC +Antibody (MT-6004): 9-O-acGD3 |
| 2010 [115] | In Sézary syndrome, circulating levels of 9-O-acGD3-positive T cells are a malignancy marker | Human PBMC | FC + Antibody (anti-CD60 from BD Biosciences): 9-O-acGD3 |
| 1992 [116] | 9-O-acGD3 is a marker of neuroectodermal cancers | Human skin from donors and nodular and sclerosis basal cell carcinoma from patients | TLC+ Antibody (JONES): 9-O-ac sialic acid |
| 2001 [117] | 9-O-acGD3 is increased in basal cell carcinoma cells | Human basal cell carcinoma tumor samples and healthy skin from patients and healthy donors | TLC + (influenza C virus and Antibody): MoAb against 9-O-acGD3 |
| 1997 [118] | 9-O-acGD3 is a marker of small cell lung cancer | Small cell and non-small cell lung cancer cell lines | Antibody (limited information) |
| 1998 [119] | In well-differentiated and invasive duct carcinoma, 9-O-acGD3 is present at the surface, with a decreased presence in non-differentiated carcinomas | Benign and malignant breast lesions and normal mammary gland tissue, cell lines of breast carcinoma (MCF-7 and EFM-19) | IHC/TLC + Antibody (M-T21): 9-O-acGD3 |
| 2019 [120] | In some breast cancer cell lines, 9-O-acGD2 and not 9-O-acGD3 has been identified | Breast cancer cell lines (Hs 578T, SUM159PT, MDA-MB-231 and MCF-7) | LCMS/FC/CM/IHC + Antibodies (7H2 mouse IgG3 and 8B6 mouse IgG3): anti-O-ac-GD3 and anti-O-acGD2, respectively. |
| 2021 [121] | CASD1 is the enzyme responsible for 9-O-acGD2 as well as for 9-O-acGD3 synthesis | SUM159PT and CHO cell lines | TLC/IHC/CM + Antibodies (M-T6004 and 8B6): 9-O-acGD3 and O-acGD2, respectively. |
| 2008 [122] | GD3 and 9-O-acGD3 increased in neural tumor cell lines High titer of anti-9-O-acGD3 antibodies in medulloblastoma patients’ serum | 13 neural tumor cell lines + NSC-34, CHO cells, and fibroblasts as controls Sera from patients with neural tumors and healthy controls | TLC/ELISA + Antibodies (R24 and D.1.1): GD3 and 9-O-ac-GD3 |
| 2011 [123] | The ratio between GD3 and 9-O-acGD3 is critical to tumor survival in glioblastoma | Three glioblastoma cell lines: SNB-19, an in-house-derived adult biopsy cell line, and IN699 | FC + Antibody (MB3.6 and Clone D1.1): GD3 and 9-O-acGD3 |
| 2002 [125], 2006 [39], 2014 [126] | GD3 is considered proapoptotic in vitro, while its 9-O-ac form is antiapoptotic | HEK-293 and U87 cells Jurkat and Molt-4 cell lines | FC/CM/TLC + Antibody (M-T6004, P-Path, UM4D4): 9-O-acGD3 |
| 2006 [39] | 9-O-acGD3 in Jurkat and Molt-4 cells prevents cell death by proapoptotic agents (N-acetyl sphingosine and daunorubicin) | Jurkat and Molt-4 cell lines | FC/CM/TLC + Antibody (M-T6004): 9-O-acGD3 |
| 2004 [127] | 9-O-acGD1 has antiproliferative effects on astrocytoma cells | Human glioma cell lines U-373 and T98G | N/A |
| 2006 [96] | In lymphocytes, acetylated gangliosides (CD60) decrease apoptosis | Human tonsillar lymphocytes | IHC/CM/FC + Antibodies (R24, UM4D4 and U5): GD3, 9-O-acGD3 and 7-O-acGD3. |
| 2007 [97] | Proapoptotic impact of 9-O-acGD3 on mature erythrocytes | Bone marrow and peripheral blood erythrocytes from children with acute lymphoblastic leukemia and clinical remission | FC + Antibody (JONES): 9-O-acGD3 |
| 1995 [129], 1997 [130] | 9-O-acGD3 as a potential target for cancer immunotherapy | 14 tumor cell lines: 7 melanomas, 3 neuroblastomas, 1 astrocytoma and 3 sarcomas | FC + Antibody (D1.1 and 5BI): 9-O-acGD3 |
| 1995 [131] | Antibody response in melanoma patients after injection of 9-O-acGD3 not antigen-specific | N/A | N/A |
| 1997 [132] | Improved antibody response in mice after injection of 9-O-acGD3 combined with VLDL and enhanced by IL-2 | BALBc mice | ELISA/TLC + Antibody (MAb 7H2) |
| 2021 [133] | 9-N-acGD2 (9-O-acGD2 surrogate) conjugated with a carrier bacteriophage (Qbeta) elicits a strong and long-lasting immune response | dogs | N/A |
| Infection | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 1996 [134] | Influenza C virus infects cells through binding to N-acetyl-9-O-ac sialic acid, like bovine coronavirus | Polarized Madin–Darby canine kidney (MDCK) cells | N/A |
| 2021 [135] | Human CoVs OC43 and HKU1, and human orthomyxovirus ICV, preferentially bind to 9-O-ac α2,8-linked sialosides | HEK-293T cells | N/A |
| 1987 [5] | Treatment of cells with 9-O acetylesterase confers resistance to influenza C virus infection; this is reversed by ganglioside containing 9-O-ac forms | MDCK II cells | N/A |
| 1992 [136] | Influenza C virus binds to 9-O-acGD1a | Immobilized glycoconjugates | TLC: 9-O-acGD1a |
| 1988 [7] | Influenza C virus is able to hydrolyze in vitro 9-O-acGD1a | N/A | TLC/LC-MS: GD1a |
| 1991 [137] | Influenza C virus is able to hydrolyze in vitro 9-O-acGT3 | N/A | TLC + Antibody (A2B5): GT3 |
| 1991 [71] | Influenza C virus hemagglutinin contains a 9-O-ac sialic-acid-specific acetyl esterase activity | Transgenic mice with partial or total loss of O-acetylation of sialic acids | IHC + Antibody (27A): 9-O- acGD3 |
| 2010 [138] | Mycobacterium leprae invades Schwann cells with the help of endogenous 9-O-acGD3; immunoblocking of the ganglioside reduces the demyelinization effect of the bacterium | Schwan cell line (ST-8814) and mice | CM/TLC + Antibody (JONES): 9-O-acGD3. Inmunoblockage |
| Autoimmune disease | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 1996 [139] | The serum of some Guillain–Barré syndrome patients reacts with 9-O-acGD1b, GD1b and GM1 | Patients’ serum | ELISA |
| 1997 [43] | Psoriatic basal and suprabasal keratinocytes display 9-O-acGD3 at the surface This is upregulated by IL-4 and IL-13, and dowregulated by IFNγ secreted by T cells | Primary cultures of keratinocytes and biopsies | FC/IHC + Antibody (UM4D4): 9-O-acGD3 |
| Toxicology | |||
| Date (Reference) | Observation | Sample | Detection Method: Target Molecule |
| 2008 [44] | Association between lead exposure and an accumulation of 9-O-acGD3 and other gangliosides in glomeruli | Male Wistar rat kidneys | IHC/TLC + Antibody (CDW60): 9-O-acGD3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera-Marcos, L.V.; Sahali, D.; Ollero, M. 9-O Acetylated Gangliosides in Health and Disease. Biomolecules 2023, 13, 827. https://doi.org/10.3390/biom13050827
Herrera-Marcos LV, Sahali D, Ollero M. 9-O Acetylated Gangliosides in Health and Disease. Biomolecules. 2023; 13(5):827. https://doi.org/10.3390/biom13050827
Chicago/Turabian StyleHerrera-Marcos, Luis Vicente, Dil Sahali, and Mario Ollero. 2023. "9-O Acetylated Gangliosides in Health and Disease" Biomolecules 13, no. 5: 827. https://doi.org/10.3390/biom13050827
APA StyleHerrera-Marcos, L. V., Sahali, D., & Ollero, M. (2023). 9-O Acetylated Gangliosides in Health and Disease. Biomolecules, 13(5), 827. https://doi.org/10.3390/biom13050827